



All-trans-retinoic acid inhibits tumour growth of malignant pleural mesothelioma in mice
- C. Tabata 1 ,
- R. Tabata 2 ,
- N. Hirayama 1 ,
- A. Yasumitsu 1 ,
- S. Yamada 1 ,
- A. Murakami 1 ,
- S. Iida 1 ,
- K. Tamura 1 ,
- T. Terada 1 ,
- K. Kuribayashi 1 ,
- K. Fukuoka 1 and
- T. Nakano 1
- 1Division of Respiratory Medicine, Dept of Internal Medicine, Hyogo College of Medicine, Nishinomiya, and 2Dept of Internal Medicine, Hyogo Prefectural Tsukaguchi Hospital, Amagasaki, Japan.
- C. Tabata, Division of Respiratory Medicine, Dept of Internal Medicine, Hyogo College of Medicine, 1-1 Mukogawa-cho, Nishinomiya, Hyogo, 663-8501 Japan. E-mail: ctabata{at}hyo-med.ac.jp
Abstract
Malignant pleural mesothelioma (MPM) is an aggressive malignant tumour of mesothelial origin associated with asbestos exposure. Because MPM has limited response to conventional chemotherapy and radiotherapy, the prognosis is very poor. Several researchers have reported that cytokines such as interleukin (IL)-6 play an important role in the growth of MPM. Previously, it was reported that all-trans-retinoic acid (ATRA) inhibited the production and function of IL-6 and transforming growth factor (TGF)-β1 in experiments using lung fibroblasts.
We investigated whether ATRA had an inhibitory effect on the cell growth of MPM, the origin of which was mesenchymal cells similar to lung fibroblasts, using a subcutaneous xenograft mouse model. We estimated the tumour growth and performed quantitative measurements of IL-6, TGF-β1 and platelet-derived growth factor (PDGF) receptor (PDGFR)-β mRNA levels both of cultured MPM cells and cells grown in mice with or without the administration of ATRA.
ATRA significantly inhibited MPM tumour growth. In vitro studies disclosed that the administration of ATRA reduced 1) mRNA levels of TGF-β1, TGF-β1 receptors and PDGFR-β, and 2) TGF-β1-dependent proliferation and PDGF-BB-dependent migration of MPM cells.
These data may provide a rationale to explore the clinical use of ATRA for the treatment of MPM.
- Cytokines
- mesothelioma
Malignant pleural mesothelioma (MPM) is an aggressive malignant tumour of mesothelial origin associated with asbestos exposure 1. Although recently asbestos usage has decreased throughout the world, the incidence of MPM is expected to markedly increase over the next few decades because there is a long latency period (20–40 yrs) between asbestos exposure and tumour development 2. MPM has limited response to conventional chemotherapy and radiotherapy, so the prognosis is very poor, with median survival durations of 8–18 months 3. Despite much research into MPM treatment, there has been little progress in effective therapeutic and preventive strategies against MPM, and the development of novel treatment is urgently needed 4.
According to recent research investigating cytokines in relation to MPM, several cytokines, such as interleukin (IL)-6 5 and hepatocyte growth factor/scatter factor 6, play an important role in the growth of MPM.
All-trans-retinoic acid (ATRA), a physiological metabolite of vitamin A, is known to affect cell differentiation, proliferation and development. ATRA has been widely used in differentiation therapy for acute promyelocytic leukaemia (APL) with the ability to overcome promyelocytic leukaemia/retinoic acid receptor fusion protein. There have been several reports about the effects of ATRA on cytokine production 7–9. ATRA induced the growth inhibition of myeloma cells, which proliferated in IL-6 autocrine and paracrine manners, with the reduction of both IL-6 production and its receptor (IL-6R) expression 10, 11. In a previous report, it was demonstrated that ATRA reduced irradiation-induced proliferation of lung fibroblasts by inhibiting both IL-6 production and IL-6R expression 12. Moreover, it was recently reported that ATRA prevented both irradiation- and bleomycin-induced pulmonary fibrosis in mice via an inhibitory effect on both IL-6-dependent fibroblast proliferation and transforming growth factor (TGF)-β1-dependent transdifferentiation of fibroblasts into myofibroblasts 13.
In the present study, we investigated whether ATRA had an inhibitory effect on the cell growth of MPM, the origin of which was mesenchymal cells similar to lung fibroblasts and associated with several cytokines, including IL-6.
METHODS
Cell culture
Human MPM cell lines H28 (epithelioid), H2052 (sarcomatoid), H2452 (biphasic) and MSTO-211H (biphasic) and human mesothelial cell line MeT-5A were obtained from the American Type Culture Collection (Rockville, MD, USA). These cells were cultured in RPMI 1640 (Sigma Chemical Co., St Louis, MO, USA) supplemented with 10% heat-inactivated fetal calf serum. ATRA (Sigma) was added to the growth medium to yield the final dimethyl sulfoxide (DMSO) solvent concentration <0.05% (v/v). In some experiments, the cells were pre-incubated with proteasome inhibitor MG-132 (5 μM) 14, Jun N-terminal kinase (JNK) inhibitor SP600125 (10 μM) 15, p38 mitogen-activated protein kinase (MAPK) inhibitor SB203580 (10 μM) 16 or extracellular signal-regulated kinase (ERK)1/2 inhibitor PD98059 (25 μM) 13 for 60 min (all Calbiochem, San Diego, CA, USA).
Animals
6-week-old C.B-17/Icr-scid Jcl (scid/scid) (SCID) female mice were purchased from Clea Japan (Tokyo, Japan) and maintained in our specific pathogen-free animal facility. All animals were kept according to the Animal Protection Guidelines of Hyogo College of Medicine (Hyogo, Japan). All protocols for animal use and euthanasia were reviewed and approved by the Institute of Laboratory Animals, Graduate School of Medicine, Hyogo College of Medicine.
Ectopic (subcutaneous) xenograft model
To produce subcutaneous (s.c.) tumours, a single-cell suspension of 107 MSTO-211H cells was implanted s.c. into the back of SCID mice. In some experiments, mice were injected intraperitoneally with 0.5 mg of ATRA dissolved in 0.1 mL cottonseed oil or with 0.1 mL cottonseed oil alone (controls). Injections were repeated three times weekly, 1) throughout the course, or 2) in the latter half of the period from inoculation to the end of the observation period. In previous studies, 0.5 mg of ATRA administration three times per week for 6 months did not produce noticeable morbidity and mortality 12, 13. The tumours were measured every 7 days with calipers, and their volumes were calculated using the formula a(b2)/2, in which a and b represent the longest and shortest diameters, respectively.
Quantitative real-time reverse transcriptase PCR
Total RNA was isolated by RNeasy Mini kit (QIAGEN, Valencia, CA, USA), and reverse-transcribed by High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). Quantitative real-time reverse transcriptase (RT)-PCR was performed as previously described 12, 13, using TaqMan Gene expression products for human IL-6, IL-6R, TGF-β1, TGF-β1 receptor (TGF-β1R) type 1, TGF-β1R type 2, platelet-derived growth factor (PDGF)-β, PDGF receptor (PDGFR)-β and cytochrome gene CYP26A1. 18S ribosomal (r)RNA served as an endogenous control (Applied Biosystems).
Measurement of nuclear factor-κB p65, p38MAPK and JNK
Nuclear extracts were prepared and protein concentration in nuclear extracts was measured as previously described 12. Nuclear factor (NF)-κB p65, cytoplasmic phospho-p38MAPK (pThr180/pThr182) and cytoplasmic JNK were detected by ELISA Kit (BioSource (Camarillo, CA, USA), Sigma and Active Motif (Carlsbad, CA, USA), respectively).
Cell proliferation assay
Cell proliferation assays were performed as previously described 12. Cells were cultured in 96-well flat-bottomed culture plates for 2 days with or without TGF-β1 (1–10,000 pg·mL−1), and/or ATRA (10−5 M).
Cell migration assay
In vitro migration assays were performed by CytoSelect 24-well Cell Migration Assay (8 μm, Colorimetric Format; CELL BIOLABS, Huissen, the Netherlands), according to the manufacturer’s instructions. Briefly, MSTO-211H cells were pre-cultured overnight with or without ATRA (10−5 M) and were then suspended at a density of 1×106 cells·mL−1 in RPMI 1640 and placed in the upper half of the Boyden chamber. The lower half of the Boyden chamber was filled with RPMI 1640 containing 10 ng·mL−1 human recombinant PDGF-BB (PeproTech, London, UK) or RPMI 1640 alone.
Analysis of apoptosis
MPM cells undergoing apoptosis were detected in tissue sections by the terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate-biotin nick-end labelling (TUNEL) method, using the in situ apoptosis detection TUNEL kit (Takara, Shiga, Japan) according to the manufacturer’s instructions.
Statistical analysis
The results are given as the mean±sd of three experiments performed in triplicate. Statistical analysis was performed using the Bonferroni/Dunn multiple comparisons test. In all tests, a p-value <0.05 was considered significant.
RESULTS
Inhibitory effect of ATRA on proliferation of MPM cells
We first investigated the effect of ATRA on the growth of both MPM (H28, H2052, H2452 and MSTO-211H) and mesothelial (MeT-5A) cells. Cells were cultured with or without various concentrations of ATRA for 48 h. The addition of ATRA had a suppressive effect on the proliferation of all of these MPM cells in a dose-dependent manner. The maximum inhibitory effect was observed at the concentration of 10−5 M ATRA (H28: 59% decrease (p<0.0001); H2042: 33% decrease (p<0.0001); H2452: 45% decrease (p<0.0001); MSTO-211H: 29% decrease (p<0.0001)) (fig. 1a–d⇓), whereas the lower concentrations (10−7 M or 10−6 M) of ATRA showed a minor effect compared with 10−5 M. Conversely, ATRA had no effect on the proliferation of MeT-5A (fig. 1e⇓). The final concentration of DMSO (0.05% v/v) had no gross effect on any cells (data not shown). The concentration of 10−5 M ATRA had no effect on cell viabilities in any cell lines (data not shown).
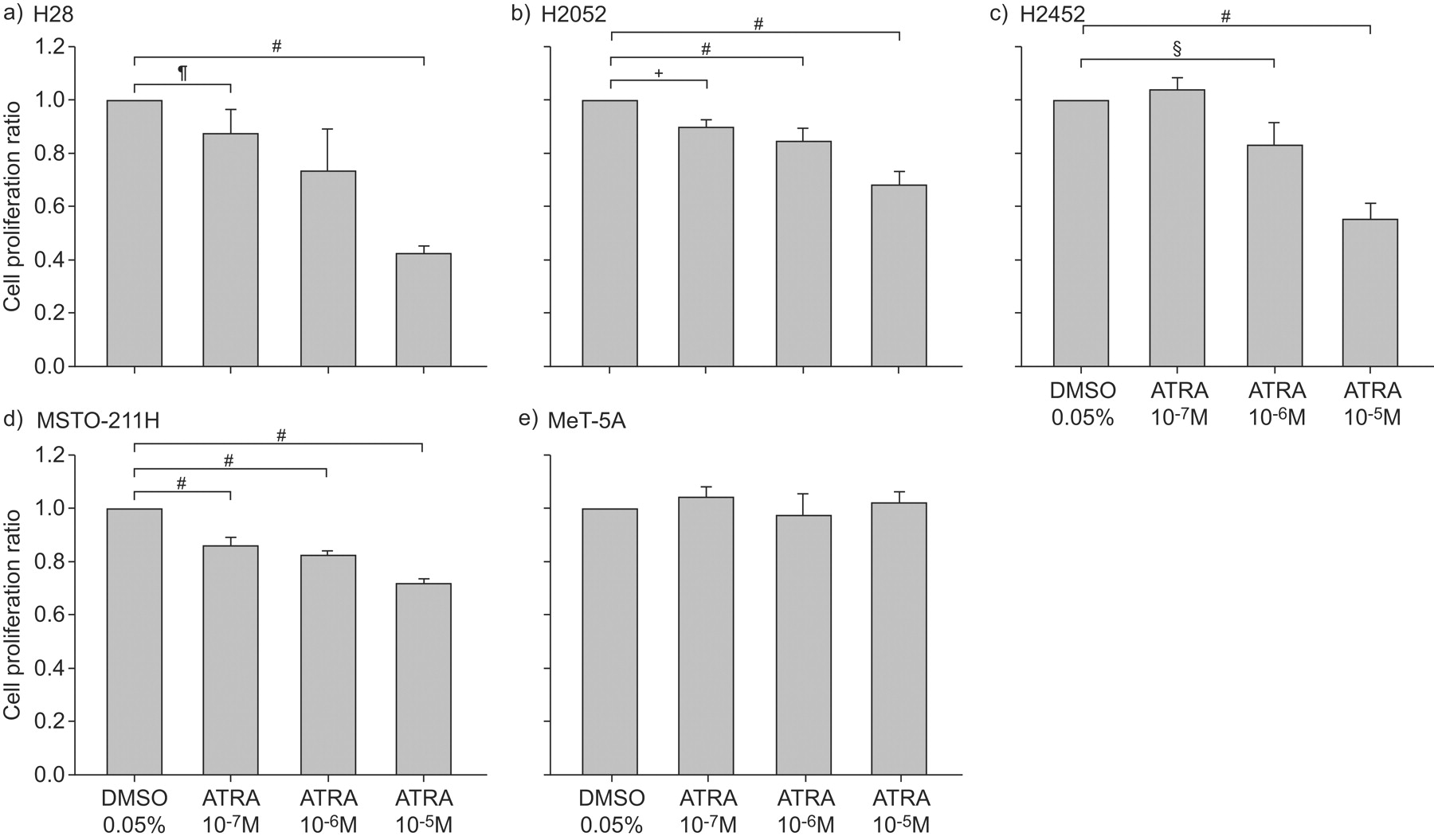
Inhibitory effect of all-trans-retinoic acid (ATRA) on proliferation of malignant pleural mesothelioma (MPM) cells. a) H28, b) H2052, c) H2452 and d) MSTO-211H MPM cells and e) human mesothelial cell line MeT-5A were cultured in 96-well flat-bottomed culture plates for 48 h in serum-free medium with or without (dimethyl sulfoxide (DMSO) alone) various concentrations (10−7, 10−6 or 10−5 M ) of ATRA, and cell proliferation was assayed. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p<0.0001; ¶: p = 0.0005; +: p = 0.0035; §: p = 0.0026.
Effect of ATRA on TGF-β1/TGF-β1R mRNA expression of MPM cells
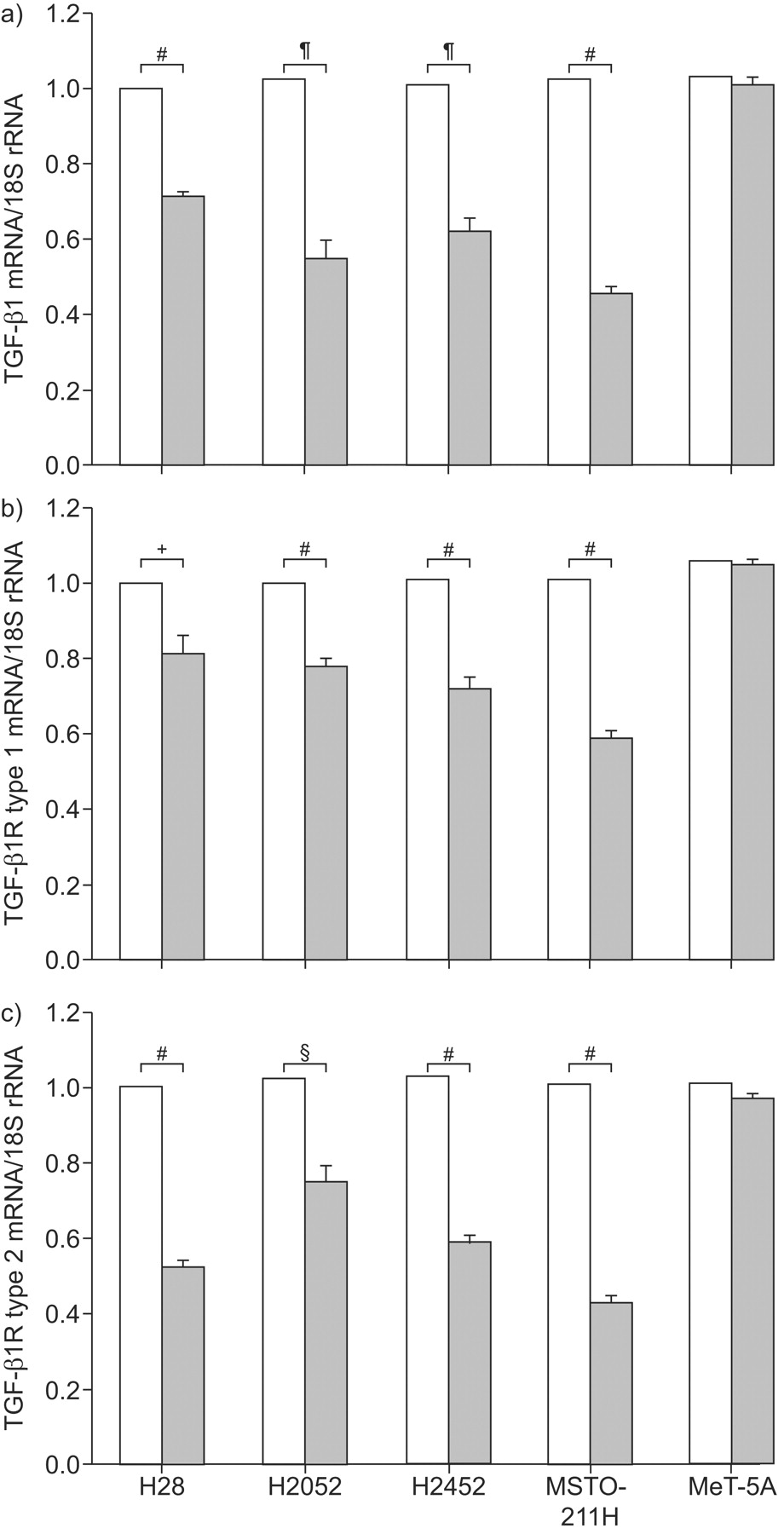
As shown in figure 2a⇓, the TGF-β1 mRNA as a ratio of 18S rRNA expression was decreased following 7 h of culture with 10−5 M ATRA: by 29% in H28 (p<0.0001), by 47% in H2052 (p = 0.0002), by 38% in H2452 (p = 0.0002) and by 56% in MASTO-211H cells (p<0.0001), compared with cells incubated with DMSO alone. Conversely, ATRA had no effect on TGF-β1 mRNA expression in MeT-5A mesothelial cells. Both TGF-β1R type 1 and type 2 mRNA/18S rRNA ratios were also decreased with ATRA compared with cells incubated with DMSO alone (type 1: 19% in H28 (p = 0.0023), 22% in H2052 (p<0.0001), 29% in H2452 (p<0.0001) and 42% in MSTO-211H (p<0.0001); type 2: 48% in H28 (p<0.0001), 27% in H2052 (p = 0.0003), 43% in H2452 (p<0.0001) and 57% in MSTO-211H (p<0.0001). Conversely, ATRA had no effect on TGF-β1R type 1 and type 2 mRNA expression in MeT-5A (fig. 2b⇓ and c).

Effect of all-trans-retinoic acid (ATRA) on transforming growth factor (TGF)-β1/TGF-β1 receptor (TGF-β1R) mRNA expression of malignant pleural mesothelioma (MPM) cells. Real-time reverse transcriptase PCR was performed to determine the changes in mRNA levels for TGF-β1/TGF-β1Rs. H28, H2052, H2452 and MSTO-211H MPM cells and the human mesothelial cell line MeT-5A were cultured in the presence (▓) or absence (dimethyl sulfoxide (DMSO) alone; □) of 10−5 M of ATRA for 7 h. The levels of mRNA for a) TGF-β1, b) TGF-β1R type 1 and c) TGF-β1R type 2 are represented as the ratio to 18S ribosomal (r)RNA, an endogenous control. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p<0.0001; ¶: p = 0.0002; +: p = 0.0023; §: p = 0.0003.
Involvement of NF-κB in the suppressive effect of ATRA on TGF-β1 mRNA expression
Proteasome inhibitor MG-132 is also known to have an inhibitory effect on NF-κB activity. Pretreatment of MPM cells with MG-132 led to decreased TGF-β1 mRNA levels by 47% in H28 (p<0.0001), 68% in H2052 (p<0.0001), 60% in H2452 (p<0.0001) and 75% in MSTO-211H (p<0.0001) compared with cells incubated with DMSO alone (fig. 3a⇓). Next, we demonstrated that the levels of nuclear NF-κB p65 in these cells were suppressed in the presence of ATRA by 11% in H28 (p = 0.0162), 13% in H2052 (p = 0.018), 10% in H2452 (p = 0.0302) and 14% in MSTO-211H (p = 0.0012) compared with cells incubated with DMSO alone (fig. 3b⇓). Although pretreatment with inhibitors for both p38MAPK and JNK led to decreased TGF-β1 mRNA levels in these cells, the level of phospho-p38MAPK and activity of JNK were not affected by ATRA (data not shown). Inhibitors of ERK1/2 had no effect on TGF-β1 mRNA levels (data not shown).

Involvement of nuclear factor (NF)-κB in the suppressive effect of all-trans-retinoic acid (ATRA) on transforming growth factor (TGF)-β1 mRNA expression. a) H28, H2052, H2452 and MSTO-211H malignant pleural mesothelioma cells were cultured in the presence (▪) or absence (dimethyl sulfoxide (DMSO) alone; □) of 5 μM MG-132 for 7 h. Real-time reverse transcriptase PCR was performed to determine changes in TGF-β1 mRNA levels. b) The activities of NF-κB were analysed. H28, H2052, H2452 and MSTO-211H cells were treated with (▓) or without (DMSO alone; □) ATRA (10−5 M) for 1 h and NF-κB p65 amounts in nuclear protein extracts were analysed. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p<0.0001; ¶: p = 0.0162; +: p = 0.018; §: p = 0.0302; ƒ: p = 0.0012.
Effect of ATRA on TGF-β1-mediated proliferation of MPM cells
To clarify the involvement of TGF-β1 in the development of MPM tumour growth, we studied the effect of TGF-β1 on the proliferation of MPM cells and MeT-5A mesothelial cells. As shown in figure 4a⇓, the addition of TGF-β1 stimulated all MPM cell growth in a dose-dependent manner and reached a plateau at the concentration of 1,000 pg·mL−1 (H28: 18% increase (p = 0.0012); H2042: 19% increase (p = 0.0015); H2452: 30% increase (p<0.0001); MSTO-211H: 46% increase (p = 0.0069)). Conversely, TGF-β1 had no effect on the proliferation of MeT-5A mesothelial cells (fig. 4a⇓). We then evaluated the effect of ATRA on TGF-β1-mediated proliferation of MPM cells, and showed that the TGF-β1-mediated proliferation was decreased with ATRA by 39% in H28 (p<0.0001), 48% in H2052 (p = 0.0013), 80% in H2452 (p<0.0001) and 73% in MSTO-211H (p<0.0001) compared with cells incubated with DMSO alone (fig. 4b⇓).

Effect of all-trans-retinoic acid (ATRA) on transforming growth factor (TGF)-β1-mediated proliferation of malignant pleural mesothelioma (MPM) cells. a) H28, H2052, H2452 and MSTO-211H MPM cells and human mesothelial cell line MeT-5A were cultured in 96-well flat-bottomed culture plates for 48 h in serum-free medium with the indicated concentrations of TGF-β1 (0–1,000 pg·mL−1) and cell proliferation was assayed. b) H28, H2052, H2452 and MSTO-211H MPM cells were cultured in the presence of 1,000 pg·mL−1 TGF-β1 with (▪) or without (dimethyl sulfoxide alone; □) ATRA (10−5 M), and cell proliferation was assayed. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p = 0.0012; ¶: p = 0.0022; +: p = 0.0017; §: p = 0.0015; ƒ: p<0.0001; ##: p = 0.0069; ¶¶: p = 0.0013.
Antitumour efficacy of ATRA in a subcutaneous xenograft model
We next examined the effect of ATRA on a s.c. xenograft mouse model of MPM cells. A single-cell suspension of 107 H28, H2052, H2452 and MSTO-211H cells with a viability of >95% was implanted s.c. into the back of SCID mice. Only MSTO-211H cells could grow on the back of SCID mice. To study the “preventive” and “therapeutic” effects of ATRA on MSTO-211H cell growth, i.p. injections of ATRA were repeated three times weekly, 1) throughout the course, or 2) in the latter half of the period from inoculation to the end of the observation period. As shown in figure 5a⇓, i.p. administration of ATRA three times per week throughout the course greatly inhibited tumour growth at 28 days after inoculation (68% decrease; p<0.0001). Moreover, treatment with ATRA three times per week for the latter half of the period from inoculation to the end of the observation period ameliorated tumour growth better than the vehicle control group (36% decrease; p = 0.0011). The i.p. injection with 0.5 mg of ATRA three times a week for 28 days had no effect on the health of the mice.

Antitumour efficacy with all-trans-retinoic acid (ATRA) in a subcutaneous xenograft model. MSTO-211H cells with a viability of >95% were implanted subcutaneously into the back of SCID mice. a) The tumours were measured and their volume calculated in mice treated with vehicle control (□), treated i.p. with ATRA three times per week throughout the course (day 0–28; ▪) or in the latter half of the period from inoculation to the end of the observation period (day 14–28; ▓). b and c) Real-time reverse transcriptase PCR was performed to determine changes in mRNA levels in tumours for b) transforming growth factor (TGF)-β1 and c) platelet-derived growth factor receptor (PDGFR)-β. The levels of mRNA are the ratio to 18S ribosomal (r)RNA. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p = 0.0014; ¶: p = 0.0011; +: p<0.0001; §: p = 0.0008; ƒ: p = 0.0004.
mRNA levels of TGF-β1 and PDGFR-β in implanted grown tumours on SCID mice
TGF-β1 mRNA levels of implanted grown MPM tumours on SCID mice at 28 days after inoculation with or without an i.p. injection of ATRA were analysed by real-time RT-PCR. We demonstrate in figure 5b⇑ that TGF-β1 mRNA expression was significantly suppressed by the administration of ATRA (32% decrease; p = 0.0008). To study whether another mechanism was involved in the preventive effect of ATRA on MPM tumour progression, we focused on previous reports demonstrating that human mesothelioma cell lines expressed PDGFR-β 17. We then analysed PDGFR-β expression in implanted grown MPM tumours and found that, in mice treated with ATRA, the levels of PDGFR-β expression were markedly decreased as compared with mice without ATRA (31% decrease; p = 0.0004; fig. 5c⇑).
Inhibitory effect of ATRA on PDGFR-β expression and PDGF-BB-induced migration of MPM cells
We next examined the impact of ATRA on PDGFR-β mRNA expression in MPM cells. As shown in figure 6a⇓, ATRA decreased the expression of PDGFR-β mRNA by 32% in MSTO-211H cells (p<0.0001); however, ATRA had no effect on the production of PDGF-BB (data not shown). It is well known that cell migration plays an important role in tumour cell invasion, especially in the wide spread of MPM tumours. We therefore performed an in vitro migration assay to study the effect of PDGF, which is a potent mitogen and chemotactic factor for several mesenchymal cells 18, on MPM progression, and revealed that MSTO-211H cell migration was induced (1.3 fold increase; p = 0.0004) by PDGF-BB, which was inhibited in the presence of ATRA (19% decrease; p = 0.0011; fig. 6b⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Inhibitory effect of all-trans-retinoic acid (ATRA) on platelet-derived growth factor (PDGF) receptor (PDGFR)-β expression and PDGF-BB-induced migration of malignant pleural mesothelioma (MPM) cells. a) Real-time reverse transcription PCR was performed to determine changes in mRNA levels for PDGFR-β. MSTO-211H MPM cells were cultured in the presence or absence (dimethyl sulfoxide (DMSO) alone) of 10−5 M of ATRA for 7 h. The levels of mRNA for PDGFR-β are the ratio to 18S ribosomal (r)RNA, an endogenous control. The results are indicated as the mean±sd of three separate experiments in triplicate. b) Cell migration assay. MSTO-211H cells were pre-cultured overnight with or without (DMSO alone) ATRA (10−5 M), and further cultured in the presence or absence of PDGF-BB (10 ng·mL−1) with or without ATRA. The results are indicated as the mean±sd of three separate experiments in triplicate. #: p<0.0001; ¶: p = 0.0004; +: p = 0.0011.
DISCUSSION
MPM is an aggressive malignant tumour of mesothelial origin associated with asbestos exposure that has limited response to conventional chemotherapy and radiotherapy; the prognosis is very poor. Recently, the multi-targeted anti-folate pemetrexed has been approved as a first-line agent in combination with cisplatin for the treatment of MPM; however, overall survival remains very poor 19.
It has previously been reported that ATRA prevented both irradiation- and bleomycin-induced pulmonary fibrosis in mice by the inhibition of both IL-6-dependent proliferation and TGF-β1-dependent transdifferentiation of lung fibroblasts. As MPM cells originate from mesenchymal cells similar to lung fibroblasts, here we examined the effect of ATRA on the progression of MPM tumour in SCID mice.
In the present study, we found that ATRA inhibited the proliferation of MPM cells but not mesothelial cells. Several factors, including IL-6, TGF-β1 and PDGF, have been reported to be associated with MPM cells 5, 6, 17, 20. Here, we first examined whether the IL-6/IL-6R system also plays an important role in ATRA-mediated inhibition of MPM cell proliferation; however, in this study, ATRA had no effect on IL-6/IL-6R mRNA expression in MPM cells.
We then investigated the effect of ATRA on the production of TGF-β1, another key cytokine in ATRA-mediated inhibition of pulmonary fibrosis, as we have previously demonstrated. We here showed that ATRA suppressed mRNA expression of both TGF-β1 and TGF-β1R in these cells and, moreover, inhibited TGF-β1-dependent cell proliferation, suggesting that ATRA demonstrated the inhibitory effect on MPM cell proliferation via a TGF-β1/TGF-β1R autocrine mechanism. Generally, TGF-β1 is produced by various normal cells observed surrounding MPM tissues, including fibroblasts, macrophages, neutrophils and lymphocytes 21, 22. In vivo, therefore, in addition to the inhibitory effect of ATRA via the TGF-β1/TGF-β1R autocrine mechanism, ATRA could decrease MPM tumour progression via the TGF-β1/TGF-β1R paracrine loop by MPM cells and these TGF-β1-producing surrounding cells. To determine the cellular mechanism in the regulation of TGF-β1 production of MPM cells, we used some well characterised pharmacological inhibitors 23. There are at least three distinct MAPK signal transduction pathways in mammalian cells that lead to activation of the ERK, JNK and p38MAPK pathways. As the induction of most cytokine genes requires the activation of NF-κB, we next examined whether changes in NF-κB activity were involved in the suppressive effect of ATRA on TGF-β1 expression in MPM cells, and found an important role of NF-κB in this process (fig. 3⇑). These results suggest a possible mechanism whereby ATRA could reduce TGF-β1 expression through an NF-κB-dependent pathway. Furthermore, it has been recently reported that the mechanism of asbestos-induced oncogenesis was associated with the activation of NF-κB 24, so the inhibitory effect of ATRA on NF-κB activation itself may be beneficial for the prevention of tumour growth in early MPM.
Next, we demonstrated in an in vivo study that i.p. administration of ATRA three times per week throughout the course greatly inhibited MPM tumour growth 28 days after inoculation. The administration of ATRA inhibited TGF-β1 mRNA expression in grown MPM tumours in SCID mice. A recent study reported that TGF-β1 was significantly associated with the growth of MPM cells in a murine MPM tumour model through TGF-β1/TGF-β1R systems and TGF-β1 signalling 25, and our study appears to support this.
It is well known that cell migration plays a pivotal role in the disease progression of cancer. PDGF acts as two types of peptide, A (16-kDa) and B (14-kDa) chains, with about 60% sequence identity, disulfide linked into three diametric molecules, PDGF-AA, -AB and -BB 26. A and B chains bind to two cell-surface receptors: the α receptor can bind all three dimers (PDGF-AA, PDGF-AB and PDGF-BB) with high affinity and the β receptor can only recognise PDGF-BB with high affinity and PDGF-AB with lower affinity 27. It has been previously reported that MPM cells express PDGF-α and PDGF-β mRNA, whereas no PDGF-β and a low level of PDGF-α mRNA expression are detected in normal mesothelial cells. Moreover, PDGFR-β mRNA expression is detected in MPM cells, whereas only PDGFR-α mRNA expression is observed in mesothelial cells 28. In this study, we demonstrated that PDGFR-β mRNA expression in MPM cells was inhibited by ATRA (fig. 6a⇑), while ATRA had no effect on PDGF-β mRNA expression (data not shown). Moreover, MPM cells migrated by PDGF-BB, which was suppressed by ATRA, as shown in figure 6b⇑. PDGF-BB is also synthesised and released by several cells associated with MPM cells growth, such as fibroblasts, vascular smooth muscle cells and vascular endothelial cells 29. The administration of ATRA had no effect on PDGF-β mRNA expression in MPM tumour cells grown on SCID mice in the present study. However, the reduction of PDGFR-β mRNA expression on MPM cells by ATRA may be estimated to inhibit the PDGF-BB/PDGFR-β paracrine loop of MPM cells and PDGF-BB-producing surrounding cells, which might increase MPM cell migration and tumour invasion in cases of human MPM. As for cell migration, it has been reported that retinoic acid inhibited fibronectin and laminin synthesis and cell migration of human pleural mesothelioma in vitro 30.
de Cupis et al. 31 demonstrated that fenretinide (4HPR), a synthetic derivative of retinoic acid, induced apoptosis of the MPM cell line ZL34. However, in their report, 4HPR induced apoptosis not only of tumour cells but also of MeT-5A, SV40-transformed normal mesothelial cells. Here, we demonstrate the selective inhibitory effect of ATRA on growth of MPM cells, but not of MeT-5A. By TUNEL assay, the increase of apoptosis was not observed in ATRA-treated MPM tumour cells in mice compared with untreated mice (data not shown). Next, we examined the mRNA expression of CYP26A1 32, a retinoic acid regulated gene, by real-time RT-PCR, and found that mRNA levels of CYP26A1 were decreased in ATRA-treated MPM tumour cells in mice compared with untreated mice, which suggested an increased amount of active retinoic acid, namely, an autoregulation feedback loop (data not shown). However, it is not clear whether the observed effect is specific to retinoic acid signalling. Although the precise cellular mechanism has not been fully investigated, we propose the possibility that TGF-β1 and PDGF receptors play an important role in this mouse model and ATRA prevents MPM cell growth through the inhibition of these cytokine/cytokine receptor systems. Namely, we propose the dual inhibitory effect of ATRA on TGF-β1-dependent proliferation and PDGF-BB-dependent migration of MPM cells, which may be the mechanism underlying the preventive and therapeutic effect of ATRA on MPM.
It is noteworthy that in this report we showed the “late”, i.e. “therapeutic”, effect of ATRA in MPM cell growth (fig. 5a⇑) in addition to the “throughout”, i.e. “preventive”, effect, because in clinical use, the therapeutic effect is often more important when clinicians find that MPM is already progressive in their patients.
ATRA is known to affect cell differentiation, proliferation and development. Clinically, ATRA has been widely used in differentiation therapy for APL 33. Furthermore, oral administration of the drug results in good compliance. Our data may lead to the development of novel strategies incorporating ATRA for the prevention and treatment of MPM.
Support statement
Funding for the present study was received from KAKENHI, Grant-in-Aid for Scientific Research (C) (20590936) and Special Coordination Funds for Promoting Science and Technology (H18-1-3-3-1) (both Tokyo, Japan).
Statement of interest
None declared.
Acknowledgments
We thank R. Morimoto and H. Kitai for technical assistance (both Division of Respiratory Medicine, Dept of Internal Medicine, Hyogo College of Medicine, Nishinomiya, Japan).
- Received December 24, 2008.
- Accepted April 8, 2009.
- © ERS Journals Ltd