Article Text

Abstract
Background: Sarcoidosis is a chronic granulomatous lung disease of unknown origin. The accumulation of activated T cells at sites of inflammation represents an early stage in granuloma formation. Since mechanisms governing the normal resolution of inflammatory processes are poorly understood, this study aimed to investigate the apoptotic phenotype of peripheral blood and lung T lymphocytes from patients with sarcoidosis.
Methods: Bronchoalveolar lavage (BAL) was performed in 10 patients with active sarcoidosis and five healthy controls.
Results: Virtually no lymphocyte apoptosis, as determined by annexin V or Hoechst staining, was seen in either patients or controls. Sustained caspase-3 activity in non-apoptotic BAL fluid lymphocytes of the patients was detected, however, in agreement with in vitro studies demonstrating caspase activation after T cell receptor (TCR) triggering as a physiological response required for efficient T cell activation. Only 11.0% (range 7.7–17.6) of the BAL lymphocytes from sarcoidosis patients were annexin V positive after exposure to the apoptotic stimulus tributyltin compared with 55.0% (range 42.0–62.0) of BAL lymphocytes from healthy controls (p<0.001). After anti-Fas treatment only 8.5% (range 6–10) of BAL fluid lymphocytes from patients but 45.5% (range 38–62) from healthy controls were apoptotic.
Conclusion: BAL fluid lymphocytes from patients with sarcoidosis display a non-apoptotic morphology associated with endogenous caspase-3 activity. They seem to be resistant to apoptosis, which might contribute to the accumulation of inflammatory cells in the lungs, persistence of inflammation, and the development and maintenance of granuloma.
- apoptosis
- sarcoidosis
- lymphocytes
Statistics from Altmetric.com
Sarcoidosis is a chronic granulomatous disease of unknown aetiology which primarily affects the lungs. In early stages of the disease there is a mononuclear cell alveolitis dominated by activated CD4+ T cells and macrophages. The coordinated interplay between these cells leads to the formation of characteristic non-caseating granulomas and, in a subgroup of patients, to fibrosis and permanently impaired lung function.1,2 During the inflammatory process the lymphocyte numbers and, in particular, certain T cell subsets are increased in the bronchoalveolar lavage (BAL) fluid.1,3 The composition of the lymphocyte subset in BAL fluid depends on the balance of immigration (regulated by adhesion molecules), local proliferation, apoptosis, and migration.4 Cytokines such as tumour necrosis factor-α (TNFα) and interferon-γ (IFNγ) are known to be important for granuloma formation,5,6 but the mechanisms by which granulomas resolve have not been well established. It is thought that the granulomatous inflammation may be resolved by the withdrawal of inflammatory cytokines and/or by induction of apoptosis.7–10
Apoptosis results from the activation of an internally encoded suicide programme induced by a variety of extrinsic and intrinsic signals. The intracellular implementation of apoptotic cell death involves the activation of a conserved biochemical pathway. A family of cysteine proteases termed caspases appears to be critical for the execution of the death signal.11 In particular, the caspase-3 subfamily has consistently been associated with apoptosis and no other function for these enzymes was known. However, recent studies have shown high levels of caspase-3 activity in the absence of apoptotic cell death in activated T cells.12–16 Moreover, caspase inhibitors block proliferation, interleukin (IL)-2 processing, and MHC class II expression in these cells,13,14 indicating that the caspase-3 family of enzymes is not only involved in the induction of cell death but might also function in lymphocyte proliferation.15 Caspase activation detected in stimulated T cells might therefore be an intrinsic part of the cellular activation machinery. However, the mechanism of caspase-3 activation in CD3 stimulated T cells remains obscure. Moreover, the T cell receptor (TCR) induced caspase activity was associated with apoptosis resistance against mitochondria triggered apoptosis and this condition seems to be transient since, when cells were stimulated with CD3 for a prolonged period, the caspase activity declined and the cells again became susceptible to apoptosis.16 It therefore seems that there is a correlation between TCR induced caspase activity, the activation state of the cell, and resistance to apoptosis.
A delicate balance between apoptosis and survival of activated inflammatory cells is thus a fundamental mechanism for the homeostasis of the immune system. Insufficient T cell apoptosis may interfere with clonal deletion and maintenance of tolerance, resulting in appropriate T cell accumulation contributing to chronic inflammation. This has indeed been observed in inflammatory bowel diseases where T cell resistance against apoptosis contributes to inappropriate T cell accumulation and perpetuation of chronic inflammation.17
Although our understanding of the regulation and molecular mechanisms of sarcoidosis has improved considerably during the last few years, it is still far from complete. In particular, mechanisms governing the normal resolution processes of the sarcoid inflammatory process are poorly understood. Only a few studies have focused on apoptosis in pulmonary sarcoidosis and these data have indicated an altered apoptotic phenotype of cells involved in the inflammatory process.18,19 Furthermore, there is conflicting information about the role of apoptosis in sarcoidosis. Cree et al20 observed apoptotic cells within granulomas of patients with sarcoidosis, but there was a wide variation in the results between individual lesions and the degree of apoptosis seemed to correlate with the spontaneous resolution of the disease. Recent studies have shown that the Fas receptor (FasR) and the Fas ligand (FasL) are expressed at higher levels in pulmonary T lymphocytes from patients with active sarcoidosis than in normal T cell populations.21,22 Also, alveolar macrophages from sarcoidosis patients have a higher expression of both FasR and TNFR1.23. Like Fas, the anti-apoptotic protein Bcl-2 is highly expressed by lymphocytes surrounding granulomatous lesions of patients with sarcoidosis.24 Previous data may therefore indicate alterations in the apoptotic phenotype of cells involved in the sarcoid process. However, the most important issue—the sensitivity to apoptosis induction—has not been established.
We have compared the sensitivity of bronchoalveolar lavage (BAL) fluid lymphocytes and peripheral lymphocytes (PBL) to apoptosis induction in patients with pulmonary sarcoidosis and healthy controls. Apoptosis was triggered via the mitochondrion by tributyltin (TBT), an agent that has previously been shown to be a potent inducer of caspase activation and apoptosis in human leukaemia T cell lines and human peripheral T lymphocytes.25–28
METHODS
Subjects
Ten patients with sarcoidosis (eight men) of median age 33.5 years (range 26–49) participated in the study. Eight were non-smokers and two smoked occasionally. The diagnosis was established in accordance with the ATS statement of sarcoidosis29 with a positive biopsy (n=5) or the clinical picture of Löfgren’s syndrome—that is, bilateral hilar lymph adenopathy (BHL) and erythema nodosum and/or ankle arthritis. In one patient the diagnosis was based on a typical clinical picture, BAL fluid lymphocytosis, BHL, and parenchyma infiltrates on the CT scan (table 1). None of the patients was being treated with corticosteroids at the time of BAL which was performed at a median of 3.5 months (range 1–16) from presentation of symptoms. The disease was classified as active according to the WASOG criteria30 in all patients. Five healthy non-smoking volunteers (three men) of median age 27 years (range 26–49) served as controls. All had a normal chest radiograph and none had any clinical signs of lung disease or any other inflammatory disease. The study was approved by the local ethical committee.
Characteristics of study population (n=10)
Separation of cells
BAL was performed and peripheral blood mononuclear cells (PBMC) isolated as previously described.31,32 For analysis by flow cytometry a lymphocyte gate was set on the basis of forward and side scatter to separate BAL lymphocytes and PBL.
Assessment of apoptotic nuclei
BAL cells or PBMC (2–3 × 106 cells) were spun at 3000g for 5 minutes and the pellet was resuspended in paraformaldehyde (4% in PBS) and spread on a poly-l-lysine (100 mg/ml) treated glass microscope slide and left to dry. The cells were rehydrated, dried, and stained with Hoechst 33342 (500 ng/ml) in the dark for 10 minutes. The slides were rinsed in water, dried, and covered with glycerol:PBS (50:50 v/v) and a cover slip. Cells were examined by fluorescence microscopy (Wetzlar, Germany) to identify condensed nuclei. At least 200 cells per slide were scored.
Measurement of phosphatidylserine
Annexin V binds specifically to phosphatidylserine (PS) that is translocated from the inner to the outer leaflet of the cell membrane during apoptosis. Propidium iodide (PI) distinguishes viable from non-viable cells, since viable cells exclude PI whereas dead cells with permeable cell membranes are permeable to PI. Exposure of PS was determined by the binding of FITC labelled annexin V (Pharmingen, CA, USA) according to the manufacturer’s protocol. Briefly, BAL cells and PBMC were resuspended in annexin binding buffer containing annexin and PI (100 μg/ml) and incubated for 15 minutes at room temperature. The fluorescence emission from a lymphocyte gate was determined using FL-1 channel (annexin) and FL-3 (PI) of a FACSCalibur, (Becton Dickinson, USA). Ten thousand events were analysed and dot plots used to determine the percentage of apoptotic cells showing annexin V-FITC binding and permeability to PI. Low fluorescence debris and necrotic cells that had lost their membrane integrity and thus were permeable to PI were gated out before analysis.
Culture conditions and induction of cell death
BAL cells and PBMC from sarcoidosis patients and healthy controls were resuspended in complete medium (RPMI-1640 containing 5% FCS, Gibco, BRL, Life Technologies, UK). The death signals were given by 2 μM tributyltin chloride (TBT; Sigma Aldrich, Germany)16,25–28 or by anti-Fas antibody (clone CH-11, 250 ng/ml) (MBL, Nagoya, Japan), and the cells were kept at 37°C in humidified air with 5% CO2 before being harvested at 1 hour and 4 hours, respectively.
Detection of caspases in intact cells
Caspase activity in intact BAL fluid lymphocytes and PBL was measured using the cell permeable substrate PhiPhilux-G2D2 according to the manufacturer’s recommendations (OncoImmunin Inc, Kensington, MD, USA). Briefly, cells were washed and resuspended in 50 μl substrate (10 μM) and incubated for 1 hour at 37°C/5%CO1. Fluorescence emission from a lymphocyte gate was determined using FL-2 channel of a FACSCalibur (Becton Dickinson, USA).
Data analysis
Differences between sarcoidosis patients and controls were tested using the non-parametric Mann-Whitney U test. The results are presented as median (range) values. A p value of <0.05 was considered significant.
RESULTS
Determination of apoptosis in PBL/PBMC and BAL fluid cells
One way of identifying apoptotic cells is to detect PS which is exposed on the outer leaflet of the plasma membrane. Freshly isolated BAL fluid lymphocytes and PBL from healthy controls and patients with sarcoidosis were therefore monitored for their exposure of PS, determined by annexin V staining. A median of 7.0% (range 4.0–10.0) and 5.3% (range 3.9–18.0) of unstimulated PBL of healthy controls and patients, respectively, were positive for annexin V (fig 1A and B). In unstimulated BAL fluid lymphocytes of controls and patients, the corresponding figures were 5.0% (range 2.0–12.0) and 9.0% (range 4.5–14.4) (fig 1C and D).

Influence of tributyltin (TBT) on the induction of apoptosis in peripheral blood lymphocytes (PBL) and BAL fluid lymphocytes of healthy controls (A and C, n=5) and patients with sarcoidosis (B and D, n=8). The cells were left unstimulated or were stimulated with 2 μM TBT for 60 minutes before being analysed for apoptotic morphology using annexin V staining.
Another characteristic feature of apoptosis is nuclear fragmentation which can be visualised as condensed nuclei by Hoechst staining. Approximately 5.0% (range 3.0–8.3) of BAL fluid lymphocytes and PBMC of controls and patients had condensed nuclei (not shown), again suggesting that most of the cells were non-apoptotic.
Induction of apoptosis in PBL and BAL fluid cells
To study the sensitivity for apoptosis induction, paired BAL fluid cells and PBMC from healthy controls and sarcoidosis patients were exposed to two classical apoptotic stimuli—anti-Fas and tributyltin (TBT)25–28—and analysed for annexin V. After 1 hour of exposure to 2 μM TBT, 85.0% (range 78.0–100.0) of PBL from controls (fig 1A) and 50.8% (range 24.0–72.1) of PBL from patients (fig 1B) became annexin V positive. In one patient the PBL appeared more resistant to apoptosis induction since TBT exposure only resulted in a 6% increase in annexin V staining (from 18.0% to 24.0%; fig 1B). This particular patient also had distinct clinical features since he was the only one with chest radiography stage III (table 1).
Following exposure to TBT for 1 hour, 55.0% (range 42.0–62.0) of BAL fluid lymphocytes of controls but only 11.0% (range 7.7–17.6) of BAL fluid lymphocytes of patients with sarcoidosis became positive for annexin V (p<0.001; fig 1C and D). The percentage of annexin positive PBL and BAL cells incubated with RPMI alone was less than 10% across the time course. Prolongation of the exposure time did not result in any additional apoptosis; instead the cells became necrotic. Thus, the increase in PS exposure following TBT treatment was significantly higher in BAL fluid lymphocytes of controls than of patients. The same trend was seen on exposure to anti-Fas antibodies with 45.5% (range 38–62) of BAL fluid lymphocytes of controls but only 8.5% (range 6–10) of BAL fluid lymphocytes of patients becoming positive for annexin V (table 2).
Sensitivity of BAL fluid lymphocytes to Fas induced apoptosis (n=4)
Detection of caspase-3 activity in PBL and BAL fluid lymphocytes
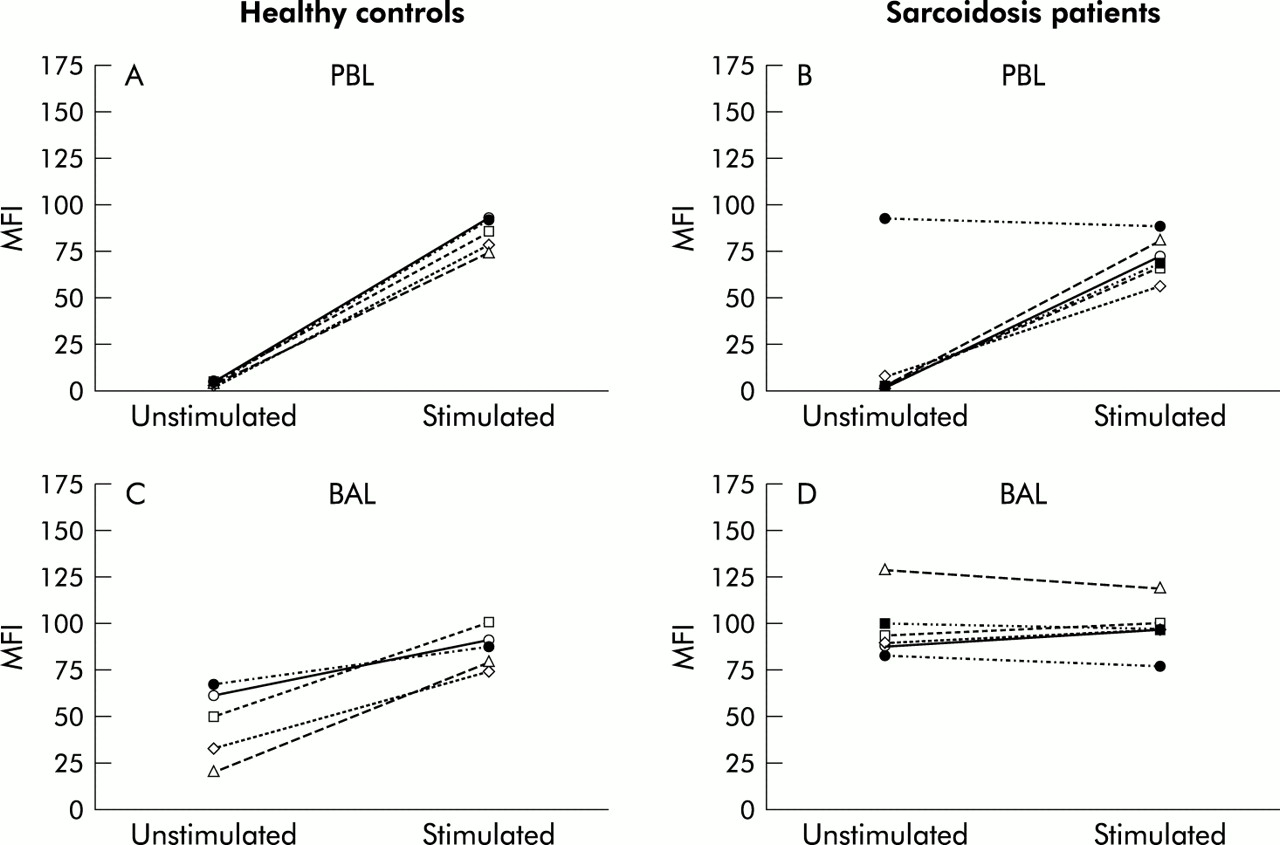
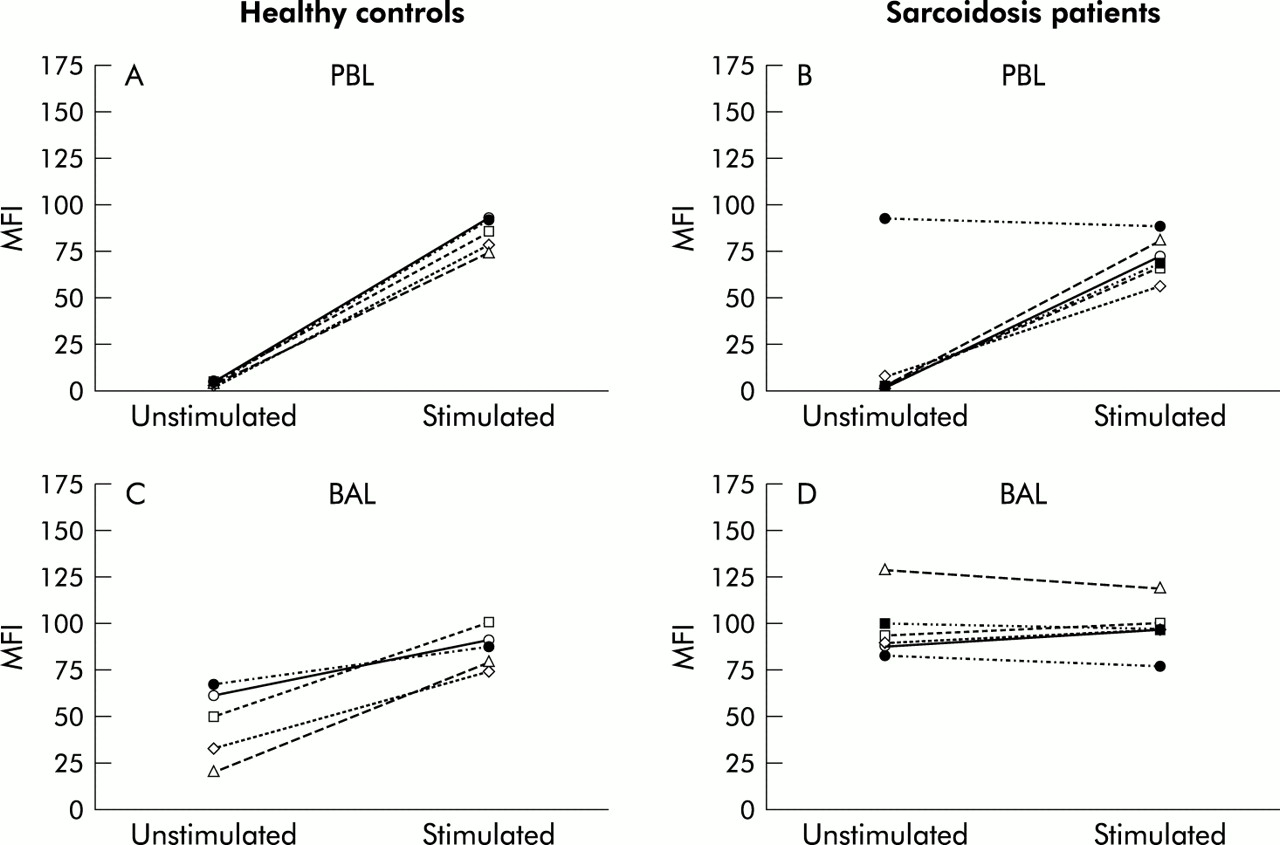
To evaluate whether BAL fluid lymphocytes and PBL from patients and controls had signs of caspase activation, cells were loaded with a fluorigenic caspase-3 substrate and the cleavage product was monitored by flow cytometry. As expected, only a small amount of caspase activity was detected in PBL of controls (mean fluorescence intensity (MFI) 4.0, range 3.0–6.0; fig 2A) and of patients with sarcoidosis (MFI 4.5, range 2.1–93.3). One patient displayed sustained caspase activity in PBL (MFI=93.3); this was the same patient who had apoptosis resistance of PBL to TBT treatment (fig 1B).

Caspase-3 activity in (A) peripheral blood lymphocytes (PBL) and (B) BAL fluid lymphocytes of patients with sarcoidosis (n=10) and healthy controls (n=5). Cells were loaded with the cell permeable substrate PhiPhilux (10 μm) to detect caspase-3 activity. Data are expressed as mean fluorescence intensity (MFI) and the horizontal lines indicate mean values.
Increased caspase-3 activity was detected in BAL fluid lymphocytes from controls (MFI 51.0, range 21.0–68.0; fig 2B), and even more pronounced caspase activity was observed in BAL fluid lymphocytes from patients (MFI 89.1, range 19.7–128.9, p=0.06; fig 2B).
Only a slight increase in caspase-3 levels was observed in BAL fluid lymphocytes from sarcoidosis patients after TBT treatment (MFI 98.0, range 78–120; fig 3D). In contrast, caspase-3 activity was increased in PBL from patients and controls and in BAL fluid cells from controls (fig 3A–C). In the patient in whom the caspase-3 activity was already increased, TBT exposure did not increase the activity further (fig 3B), nor were the PBL lymphocytes apoptotic after treatment (fig 1B).
{kind=link}
{kind=link}
{kind=link}
Influence of tributyltin (TBT) treatment on caspase-3 activity in peripheral blood lymphocytes (PBL) and BAL fluid lymphocytes of healthy controls (A and C, n=5) and patients with sarcoidosis (B and D, n=8). The cells were left unstimulated or were stimulated with 2 μM TBT for 60 minutes before being labelled with the cell permeable substrate PhiPhilux (10 μM) to detect caspase-3 activity. The data are expressed as mean fluorescence intensity (MFI).
DISCUSSION
We have analysed the apoptotic phenotype and its susceptibility to apoptosis induction in freshly isolated PBMC and BAL fluid lymphocytes from 10 patients with sarcoidosis and five healthy controls. Apoptosis was determined both by annexin V, for detection of exposure to PS, and by Hoechst staining for determination of condensed apoptotic nuclei. About 5–10% of the lymphocytes were positive for annexin or Hoechst, whereas positive signals were detected for both FasR and CD69 on most BAL fluid cells, as previously reported.31–33 The BAL fluid cells therefore seem highly activated but non-apoptotic.
The removal of activated lymphocytes by apoptosis is one prerequisite for the resolution of inflammation. It is still unclear whether a defect in the mechanism of apoptosis could prolong T cell survival in patients with sarcoidosis and thereby contribute to the chronic inflammation. Failure of cells to die might be one mechanism contributing to the formation of immune granulomas, characterised by the persistence of inflammation at the site of disease activity. In other inflammatory conditions such as inflammatory bowel disease a disturbed T cell apoptotic phenotype has been demonstrated.17 A number of apoptosis associated molecules regulate immune mechanisms implicated in the sarcoid inflammatory process. Furthermore, the speed of the resolution process of the granulomatous disorders is highly dependent on the local production of signals, favouring or suppressing the apoptotic process when cytokines are withdrawn.7–10
The FasR/FasL system plays a crucial role in limiting the expansion of T cells and preventing excessive accumulations of activated cells at the site of inflammation. It has been shown that FasR and FasL were expressed at higher levels in pulmonary lymphocytes of sarcoidosis patients than in normal T cell populations, but apoptosis was only observed in the patients who improved during the follow up period which suggests that the Fas system might play an important role, especially in the resolution phase of the granulomatous inflammation.22 We have shown that, despite high FasR expression, anti-Fas monoclonal antibody treatment did not result in extensive apoptosis of BAL lymphocytes from the patients with sarcoidosis. It is thus likely that the apoptotic machinery in these cells is “switched off” and apoptosis cannot be induced no matter whether apoptosis is triggered via the receptor pathway (by anti-Fas) or via the mitochondrion (by TBT). However, the FasR/FasL pathway can still play a crucial role in the resolution phase of inflammation.
We have shown that activated human PBL resist apoptosis induction during the initial 3–5 days of CD3 stimulation.16 The resistance was associated with a sustained caspase-3 activity in these cells, in line with studies which show that caspase activation after TCR triggering may be a physiological response required for efficient T cell activation.12–16 We have detected a large amount of caspase-3 activity in the BAL fluid lymphocytes of the patients with sarcoidosis and, as expected, these cells were also largely resistant to apoptosis induction. Exposure to TBT did not increase the caspase-3 activity any further in these cells, nor did it induce apoptotic morphology. Instead, the cells became necrotic several hours later. In contrast, when BAL fluid lymphocytes from healthy controls were similarly treated, apoptosis was successfully induced. The role of caspases in the process of T cell activation is therefore still not clearly defined, neither is it apparent why target proteins in cells with high caspase-3 activity are protected from degradation. It is clear, however, that the activity of caspases must be tightly regulated to protect regulatory and other vital protein structures from proteolytic degradation.
PBL from healthy controls (representing a low activated state) were most susceptible to apoptosis induction, whereas BAL fluid lymphocytes from patients (representing a highly activated state) were less susceptible. A similar degree of susceptibility towards apoptosis induction was observed in BAL fluid lymphocytes from healthy controls and PBL from sarcoidosis patients, where both populations of cells represent an intermediate degree of activation. Our results indicate the importance of the activation state of the T cells at the time of exposure to an apoptotic stimuli. The wide variation in the results between individual lesions, which has been observed previously,20 might be due to differences in the activation/proliferation state of the cells.
T lymphocytes already engaged in an immunological response must proliferate and differentiate into effector cells. The need to control and avoid premature death of cells participating in an immune response is of physiological relevance and may be related to a short period of resistance towards apoptosis following activation. The apoptosis resistant phenotype of BAL fluid lymphocytes from patients with sarcoidosis might therefore be linked to the highly activated state of these cells.
The only patient with a stage III disease had particularly high caspase-3 activity in PBL in the absence of an apoptotic morphology, and these cells also displayed resistance to apoptosis induction but, overall, the disease stage did not obviously influence the number of apoptotic cells in this group of sarcoidosis patients.
In conclusion, our results found no apoptotic morphology in either fresh BAL fluid lymphocytes or PBL from patients with sarcoidosis or controls, despite high levels of caspase-3 activity in the BAL fluid lymphocytes of the patients. Moreover, BAL lymphocytes from sarcoidosis patients were resistant to apoptosis induction and this mechanism might partially explain the accumulation of inflammatory cells and the persistence of chronic inflammation. The high caspase-3 activity detected in BAL fluid lymphocytes of patients with sarcoidosis probably reflects the physiological response after TCR triggering, as previously shown in vitro, and this phenomenon has now been shown for the first time in vivo in a pathological condition.
REFERENCES
Footnotes
-
This work was supported by grants from The Swedish Medical Research Council, The Swedish Heart Lung Foundation, The King Oscar II Jubilee Foundation, Åke Wiberg Foundation and Karolinska Institutet.
Linked Articles
- PostScript