Article Text

Abstract
Background: Smooth muscle contraction is one of the hallmarks of asthma. A recently developed pyridine derivative, Y-27632, a selective Rho kinase inhibitor, has been reported to inhibit the smooth muscle contraction of human and animal trachea in ex vivo systems but its effect in animal models of airway hyperresponsiveness (AHR) has not been examined. The purpose of this study was to evaluate the effect of Y-27632 in a murine model of allergic and virally induced AHR.
Methods: Baseline lung resistance and methacholine induced AHR were measured in mice sensitised to ovalbumin (OVA) and also in mice infected with respiratory syncytial virus (RSV) following ovalbumin sensitisation (OVA/RSV).
Results: Time course and dose ranging experiments indicated that 30 mg/kg Y-27632 given by gavage 2 hours before methacholine challenge significantly reduced baseline lung resistance and prevented AHR in OVA sensitised mice. Y-27632 also suppressed AHR induced by the bronchospastic agent serotonin in OVA sensitised mice and prevented methacholine induced AHR in OVA/RSV mice.
Conclusions: These results suggest that the signalling pathway mediated through Rho kinase may have an important role in bronchial smooth muscle tone in allergen induced and virus induced AHR and should be considered as a novel target for asthma treatment.
- rho kinase inhibitor
- Y-27632
- respiratory syncytial virus
- airway hyperresponsiveness
Statistics from Altmetric.com
Smooth muscle contraction is one of the hallmarks of asthma. Smooth muscle tone is primarily regulated by the level of phosphorylation of myosin light chain (MLC) by both calcium (Ca2+) dependent and independent mechanisms. An increase in the cytoplasmic concentration of Ca2+ leads to Ca2+-calmodulin complex formation which activates MLC kinase (MLCK). This enzyme phosphorylates MLC, resulting in smooth muscle contraction.1 In the presence of a fixed Ca2+ concentration, bronchoconstrictive agonists can also increase MLC phosphorylation. In addition, inhibition of MLC phosphatase (MLCP), which effectively raises MLC phosphorylation, can occur at a fixed cytoplasmic Ca2+ concentration.2 Ca2+ independent mechanisms for MLC phosphorylation and smooth muscle contraction are referred to as Ca2+ sensitisation.3,4 The mechanisms underlying Ca2+ sensitisation have not been fully elucidated, but RhoA, a small GTPase in the ras superfamily, and its target Rho associated kinase are involved in this process.5–7 Rho associated coiled coil-forming protein kinase (ROCK I) and its isoform ROCK II8–10 inhibit MLCP activity and promote the activity of MLCK, thereby increasing the level of MLC phosphorylation and increasing muscular tension.11 Blocking RhoA signalling through Rho kinase would therefore be expected to attenuate contractile responses via the inhibition of the Ca2+ sensitising mechanism.
Recent studies have shown that a pyridine derivative, (+)-(R)-trans–4-(1-aminoethyl)-N-(4-pyridyl)cyclohexane carboxamide (Y-27632), a selective ROCK family inhibitor (Ki value for ROCK I: 0.22 μM, for ROCK II: 0.30 μM),12 decreased blood pressure of hypertensive rats and inhibited smooth muscle contraction ex vivo in rabbit aorta and in guinea pig and bovine trachea.10,13 Furthermore, Y-27632 relaxes smooth muscle in the human bronchus14,15 and pulmonary artery.15 We hypothesised that the Rho/ROCK mediated signalling pathway may be a novel target for the treatment of allergen or virus induced airway hyperresponsiveness (AHR), two of the most common triggers of acute exacerbations of asthma.16–18
To our knowledge the in vivo effects of a ROCK inhibitor such as Y-27632 on bronchial reactivity have not previously been reported. We evaluated the effect of Y-27632 on baseline lung resistance and bronchial reactivity in two well characterised murine models of AHR; OVA sensitised mice (OVA mice), a model of allergic airway inflammation, and OVA sensitised mice infected with RSV (OVA/RSV mice), a model of virus induced AHR.19
METHODS
Mice
Pathogen free 8 week old female BALB/c mice were purchased from Jackson Laboratory (Bar Harbor, ME, USA). Investigators adhered to the Care and Use of Laboratory Animals of the Institute of Laboratory Animal Resources, National Research Council (NIH Publication No 86-23, revised 1985).
Reagents
Acetyl-β-methacholine (methacholine), serotonin (5-hydroxytryptamine, 5HT), and ovalbumin (chicken OVA, grade V) were purchased from Sigma Chemical Co (St Louis, MO, USA). Y-27632 was a generous gift from Welfide Corporation (Iruma, Japan).
Virus
The A2 strain of RSV was provided by Dr Robert Chanock, National Institutes of Health. Master stocks and working stocks of RSV were prepared as previously described.20
Allergen sensitisation
Mice were injected intraperitoneally with 0.1 ml (10 μg) ovalbumin complexed with 2 mg Al(OH)3 as previously described.19 After 14 days the mice were placed in an acrylic box and exposed to aerosols of 1% ovalbumin diluted in sterile phosphate buffered saline (PBS) using a jet nebuliser (Ultraneb 99, DeVilbiss, Somerset, PA, USA) for 40 minutes each day for 8 days.
Methacholine and serotonin challenges
AHR was measured as previously described.19 Methacholine and serotonin were dissolved in normal saline and administered intravenously at starting doses of 5 μg/kg and 6.25 μg/kg, respectively. The mean volume per methacholine and serotonin dose was approximately 35 μl and 50 μl, respectively. The concentrations of methacholine and serotonin were increased threefold and twofold, respectively.
Administration of Y-27632
Y-27632 was dissolved in distilled water and diluted to given concentrations. The mice were held upright with the neck fully extended and 100 μl Y-27632 was administered by gavage.
Experimental design for establishing timing and dose of Y-27632
We first determined the optimal timing of treatment. One day after the 8 day OVA aerosol treatment, AHR was measured by methacholine challenge. Mice were treated with Y-27632 in a dose of 30 mg/kg 24 hours, 8 hours, 4 hours, and 2 hours before methacholine challenge. A control group was treated with water. A dose ranging experiment was then performed. Methacholine induced AHR was measured in OVA mice treated with Y-27632 in doses of 3 mg/kg, 10 mg/kg, and 30 mg/kg 2 hours before methacholine challenge. A control group received water only.
Evaluation of Y-27632 on OVA sensitised RSV infected mice
Mice were sensitised with OVA as described above and infected with RSV (100 μl inoculum of 107 plaque forming units of RSV) as previously described on day 3 of OVA inhalation.19 Fourteen days after RSV inoculation (and 9 days after OVA inhalation) the mice underwent methacholine challenge and AHR was measured.
Statistical analysis
Percentage of baseline resistance was calculated from the lung resistance at each dose of methacholine or serotonin divided by baseline lung resistance. Results are expressed as mean (SE). The D'Agostino Kurtosis test was performed to determine if the data were normally distributed. Data for the baseline resistance with Y-27632 versus water in the non-sensitised mice, OVA sensitised mice, and OVA/RSV mice were normally distributed and were compared by an unpaired t test. Data for the methacholine challenges establishing the time course and dose response curves to Y-27632 were normally distributed and were compared by repeated measures ANOVA with Fisher's least significant difference performed as a post hoc analysis. Data for the serotonin challenge and the methacholine challenges comparing Y-27632 versus water in the OVA sensitised and non-sensitised mice were also normally distributed and were compared using a repeated measures unpaired t test. Data for the methacholine challenge comparing Y-27632 with water in the OVA/RSV mice were not normally distributed and were compared using a repeated measures Mann-Whitney non-parametric test. Differences were considered to be significant if the p value was <0.05.
RESULTS
Effect of Y-27632 on baseline lung resistance in OVA sensitised mice
Baseline lung resistance was measured in both water and Y-27632 (30 mg/kg) treated mice in three groups: non-sensitised mice, OVA mice, and OVA/RSV mice. There was no significant difference in baseline airway resistance between the water gavage and Y-27632 (30 mg/kg) gavage group in non antigen-sensitised mice (table 1). In contrast, Y-27632 reduced baseline lung resistance in OVA mice (p=0.04). In OVA/RSV mice there was a trend toward an increase in baseline resistance in water treated mice compared with Y-27632 treated mice (p=0.10). This difference was not statistically significant because of lack of power.
Effect of Y-27632 on baseline lung resistance in OVA sensitised mice
Effect of Y-27632 on AHR in OVA sensitised mice
Airway responsiveness to intravenous methacholine was measured in mechanically ventilated mice in a similar fashion to that used previously.19,21 A time course experiment was performed to determine the interval between Y-27632 dosing and methacholine challenge which had the maximal effect on AHR (fig 1A). The maximal effect of Y-27632 was observed in the group treated 2 hours before methacholine challenge (p=0.005 v non-treated OVA sensitised mice). AHR was reduced in mice treated with Y-27632 4 hours before the methacholine challenge compared with non-treated mice (p=0.04). Furthermore, there was a statistically significant difference in AHR between the groups treated with Y-27632 2 hours and 24 hours before the methacholine challenge (p=0.02), and a trend towards a difference in AHR between the groups treated 2 and 8 hours before the methacholine challenge (p=0.08). At a methacholine dose of 441 μg/kg, the mean (SE) baseline lung resistance was 128.3 (7.7)%, 261.1 (19.2)%, 382.0 (64.7)%, 526 (134.0)%, and 513.0 (149.2)% in the groups treated 2, 4, 8, and 24 hours respectively before the methacholine challenge and in the untreated group. Administration of Y-27632 more than 8 hours before the methacholine challenge did not result in a significant difference in AHR from the untreated group.

Inhibition of AHR by Y-27632 in OVA sensitised mice. (A) Mice were sensitised with OVA and Y-27632 was administered 2 hours (•, n=4), 4 hours (♦, n=4), 8 hours (▴, n=4), and 24 hours (▪, n=3) before methacholine challenge. A control group (ø, n=3) was untreated. Each point represents the mean (SE) value. *p<0.05, **p<0.001 compared with no gavage group; ***p<0.03 compared with the group treated 24 hours before methacholine challenge. (B) Y-27632 was given before 2 hours before the methacholine challenge at doses of 3 mg/kg (▪, n=2), 10 mg/kg (▴, n=5), or 30 mg/kg (•, n=5). A control group was treated with water gavage (♦, n=5). Each point represents the mean (SE) value. *p<0.02; **p<0.001 compared with water gavage group.
We then performed a dose ranging experiment 2 hours before the methacholine challenge (fig 1B). The increase in lung resistance induced by methacholine was suppressed by Y-27632 in a dose related fashion. The maximal effect of Y-27632 was observed in the group treated with 30 mg/kg (p=0.0007), but Y-27632 in a dose of 10 mg/kg also reduced AHR compared with water (p=0.01). There was a trend in the difference in effect of Y-27632 in doses of 30 mg/kg and 3 mg/kg on AHR (p=0.06). At a methacholine dose of 441 μg/kg the mean (SE) baseline lung resistance was 456.5 (100.6)%, 316.6 (67.0)%, 232.8 (31.1)%, and 129.4 (10.1)% in groups treated with water and Y-27632 in doses of 3 mg/kg, 10 mg/kg, and 30 mg/kg, respectively.
Effect of Y-27632 on serotonin induced AHR in OVA sensitised mice
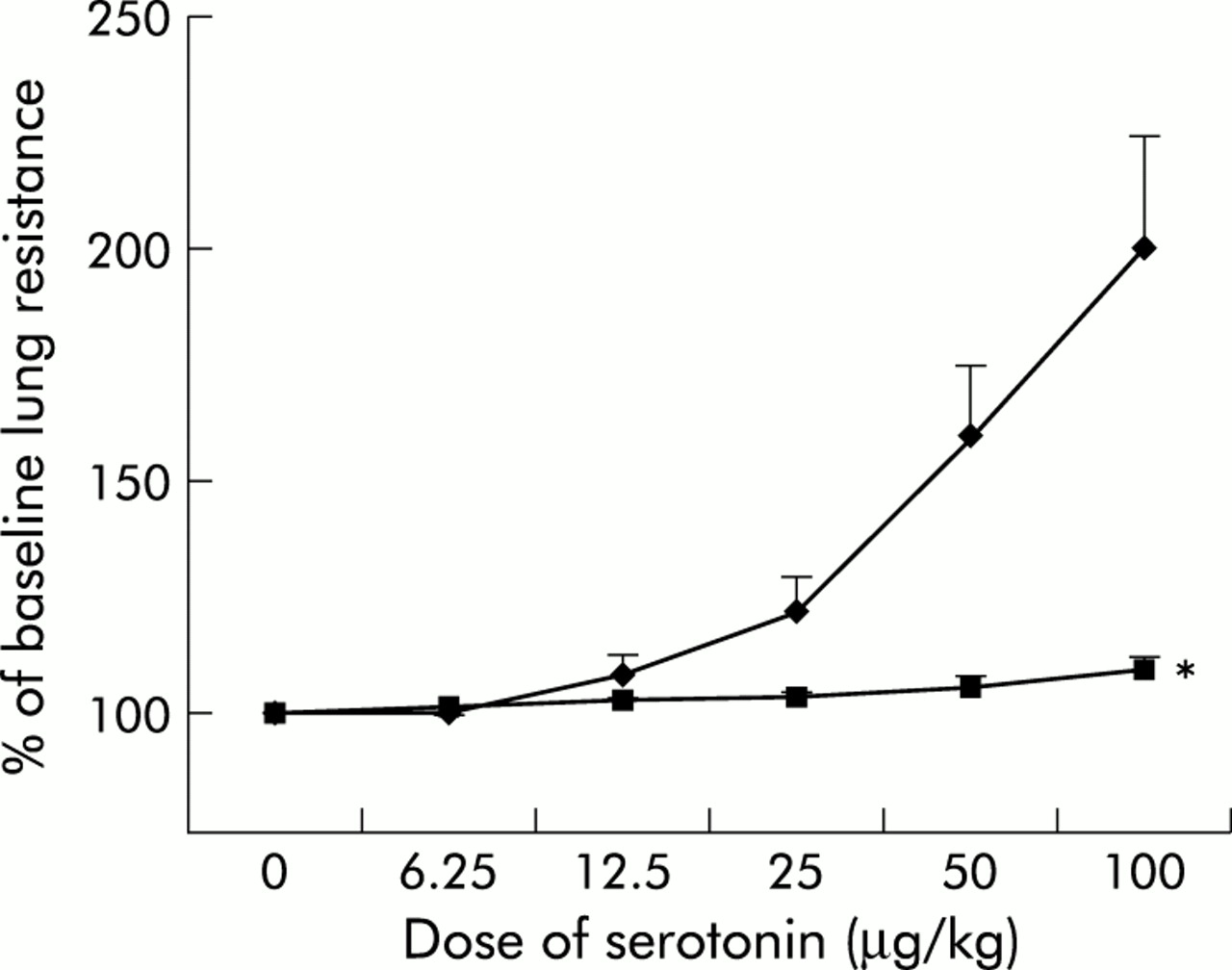
Y-27632 significantly suppressed serotonin induced AHR in OVA sensitised mice (p<0.001, fig 2).

Inhibition of serotonin induced airway hyperresponsiveness by Y-27632 in OVA sensitised mice. Mice were sensitised with OVA and treated with water (♦, n=5) or Y-27632 30 mg/kg (▪, n=7) 2 hours before serotonin challenge. Each point represents the mean (SE) value. *p<0.01 compared with water gavage group.
Effect of Y-27632 on methacholine induced lung resistance in non-sensitised mice
Y-27632 administered to mice orally at a dose of 30 mg/kg 2 hours before methacholine challenge significantly suppressed the methacholine induced increase in lung resistance in non-sensitised mice (p=0.01, fig 3). At a methacholine dose of 411.0 μg/kg the mean (SE) baseline lung resistance in the water and Y-27632 gavage groups was 228.6 (41.8)% and 112.7 (2.0)%, respectively.

Effect of Y-27632 on methacholine induced lung resistance in non-sensitised mice. Mice were separated into two groups and given water (♦, n=6) or Y-27632 30 mg/kg (▪, n=6) 2 hours before measurement of baseline lung resistance. Each point represents the mean (SE) value. *p<0.02 compared with water gavage mice.
Effect of Y-27632 on AHR in OVA sensitised RSV infected mice
Y-27632 administered orally in a dose of 30 mg/kg 2 hours before methacholine challenge significantly suppressed AHR in OVA/RSV mice (p=0.02, fig 4). At a methacholine dose of 411.0 μg/kg the mean (SE) baseline lung resistance in the water and Y-27632 treated groups was 684.8 (186.6)% and 183.8 (21.7)%, respectively.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of Y-27632 on lung resistance in OVA sensitised RSV infected mice. Mice were infected with RSV during OVA sensitisation and treated with water (♦, n=7) or Y-27632 30 mg/kg (▪, n=6) 2 hours before methacholine challenge. Each point represents the mean (SE) value. *p<0.03 compared with water gavage group.
DISCUSSION
This is the first report to suggest that in vivo inhibition of RhoA signalling may be a target for the treatment of asthma. Y-27632 significantly suppressed airway responsiveness induced by bronchospastic agonists in normal mice, allergen sensitised mice, and allergen sensitised mice infected with a virus.
Airway constriction caused by smooth muscle contraction is a critical feature of asthma. Histamine, neurokinin A, cysteinyl leukotriene, serotonin, methacholine, and acetylcholine are known bronchospastic agonists.22,23 Stimulation of their respective trimeric G protein-coupled receptors activates phospholipase C signalling pathways,24–26 leading to intracellular Ca2+ flux and smooth muscle contraction. Rho kinase inhibition can relax bronchi precontracted by histamine and neurokinin A, suggesting that RhoA activation is an integral part of the pathway leading to smooth muscle contraction.15 Methacholine and acetylcholine stimulate both muscarinic 3 (M3) and muscarinic 2 (M2) receptors.22,27 These pathways can also activate Rho/ROCK mediated signalling resulting in Ca2+ sensitisation.28 Y-27632 relaxes the bronchi precontracted by acetylcholine, histamine, and neurokinin A, and also relaxes the basal tone of isolated human bronchi in vitro.15
We have evaluated the effects of Y-27632 on baseline lung resistance and methacholine induced AHR in three distinct settings with different patterns of inflammation and cytokine production. Normal mice have no lung inflammation and do not have detectable levels of type 1 or type 2 cytokines. The low level of baseline lung resistance in normal mice is not modified by Rho kinase inhibition, but methacholine induced AHR is completely inhibited. OVA sensitised mice have airway inflammation characterised by the presence of lymphocytes and eosinophils, and interleukin (IL)-4, IL-5, IL-13, and IgE are readily detected. OVA sensitised mice infected with RSV not only have type 2 cytokines, IgE, and eosinophilia, but also abundant interferon (IFN)-γ and IL-10, resulting in a longer period of methacholine induced AHR than in mice sensitised with OVA alone.29 In addition, the RSV F glycoprotein has been shown to interact with RhoA and RSV infection can activate RhoA signalling pathways potentially providing another direct stimulus for smooth muscle contraction in RSV infected mice.30 Rho kinase inhibition reduced baseline lung resistance in OVA mice and methacholine induced AHR in both OVA and OVA/RSV mice, suggesting that the inhibition was intrinsic to smooth muscle and not related to the pattern of inflammation, cytokine production, or direct effects of RSV. The inhibition of serotonin induced AHR also suggested that Rho kinase inhibition works through a final common pathway, irrespective of the initiating stimulus for smooth muscle contraction.
These results suggest that the signalling pathway mediated through Rho kinase may have an important role in bronchial smooth muscle tone in allergen induced and virus induced AHR and should be considered as a novel target for asthma treatment.
Acknowledgments
This project was supported by K08-HL-03730, American Lung Association of Tennessee, GM 15431, and RO1-AI-45512. The authors thank Welfide Corporation for generously providing Y-27632.
REFERENCES
Footnotes
-
K Hashimoto and R Stokes Peebles Jr contributed equally to this work.