


Abstract
The effect of four macrolide antibiotics (roxithromycin, clarithromycin, erythromycin, and azithromycin) on the generation of some mediators and cytokines involved in the inflammatory process has been studied both in vivo and in vitro. Rat carrageenin pleurisy was used as a model of acute inflammation, and the macrolides were administered (10, 20, and 40 mg/kg p.o.) 1 h before the carrageenin challenge. Exudate volume and leukocyte accumulation were both dose-dependently reduced by roxithromycin, clarithromycin and erythromycin in either normal or adrenalectomized animals. Furthermore, in normal rats, prostaglandin (PG)E2, nitrate plus nitrite, and tumor necrosis factor-α levels in pleural exudate were significantly reduced by these macrolides. Roxithromycin appeared more effective than erythromycin and clarithromycin, whereas azithromycin only slightly affected the inflammatory reaction. None of the macrolides were able to modify leukotriene B4 exudate levels. In vitro experiments have shown that the four macrolides (5–80 μM) reduced in a concentration-dependent manner the production of 6-keto-PGF1α, NO2−, tumor necrosis factor-α, interleukin-1β, and interleukin-6 by lipopolysaccharide-stimulated J774 macrophages. In J774 cells, the inhibition of 6-keto-PGF1α and NO2− production by roxithromycin and erythromycin was not dependent on direct inhibition of cyclooxygenase-2 and inducible nitric oxide synthase activity because it appears to be related to the inhibition of cyclooxygenase-2 and inducible nitric oxide synthase protein expression. In conclusion, the present study shows that macrolide antibiotics have anti-inflammatory activity, which likely depends on their ability to prevent the production of proinflammatory mediators and cytokines, and suggest that these agents, particularly roxithromycin, can exert therapeutic effects independently of their antibacterial activity.
Macrolide antibiotics are active against Gram-positive bacteria,Mycoplasma spp., Legionella spp.,Chlamydia spp., and Haemophilus influenzae (Barry et al., 1987; Young et al., 1989). Apart from their antibacterial activity, these agents exhibit a broad spectrum of pharmacological effects (Bryskier et al., 1994), including anti-inflammatory activity in humans and animals (Tarayre et al., 1987; Mikasa et al., 1992; Agen et al., 1993). Macrolides have been shown to affect several pathways of the inflammatory process, such as the migration of neutrophils, the oxidative burst in phagocytes, and the production of proinflammatory cytokines (Takeshita et al., 1989; Hand et al., 1990; Mikasa et al., 1992; Konno et al., 1994). Although the precise mechanisms of these effects are not clear, it has been suggested that the interaction between macrolides and leukocytes may be important. In fact, macrolide antibiotics are able to accumulate into polymorphonuclear leukocytes, reaching intracellular concentrations far higher than those attained in the extracellular fluids (Laufen et al., 1985; Hand et al., 1987). This ability may in turn alter the functions of phagocytes, which appear crucial for both the antibacterial defense and the inflammatory process often associated with infections. Some have suggested that the antioxidant properties, shared by several macrolides (Labro et al., 1989), may play a role in the anti-inflammatory activity of these agents (Plewig and Schöpf, 1976; Dalziel et al., 1987). In conclusion, although the evidence so far accumulated shows that macrolides do exert both local and systemic anti-inflammatory effects, the mechanisms underlying these actions are still unclear. In this study, we investigated, both in vivo (rat carrageenin pleurisy) and in vitro [lipopolysaccharide (LPS)-stimulated J774 murine macrophages], the effect of four macrolide antibiotics (roxithromycin, clarithromycin, erythromycin, and azithromycin) on the generation of some mediators and cytokines involved in the inflammatory process, such as arachidonic acid metabolites, nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6.
Materials and Methods
Chemicals.
Roxithromycin was obtained from Hoechst-Marion Roussel (Romainville, France). Erythromycin, prostaglandin (PG)E2, 6-keto-PGF1α, leukotriene (LT)B4 rabbit antisera, and anti-β-actin mouse antibody were obtained from Sigma (Milan, Italy). Clarithromycin and azithromycin were purified from the commercial products Klacid (Abbott) and Zithromax (Pfizer), respectively. LPS was derived from Salmonella typhosa (0901) purchased from Difco (Detroit, MI). Dulbecco's modified Eagle's medium and fetal bovine serum were obtained from BioWhittaker (Heidelberg, Germany). Carboxymethylcellulose and cadmium powder (325 mesh) were obtained from Aldrich (Milan, Italy). Nonfat dry milk and nitrocellulose membranes were obtained from Bio-Rad (Milan, Italy). Anti-cyclooxygenase (COX)-2 and anti-inducible NO synthase (iNOS) mouse antibodies were purchased from Transduction Laboratories (Lexington, KY). Anti-mouse immunoglobulins coupled to peroxidase were purchased from Amersham (Milan, Italy). TNF-α and IL-1β enzyme-linked immunosorbent assay (ELISA) kits were obtained from Genzyme (Milan, Italy). IL-6 ELISA kits were obtained from Endogen (Woburn, MA). All other compounds were from Sigma.
Animals.
Male Wistar rats (Harlan, Italy) weighing 240 to 260 g were used for this study. Animals were housed in propylene cages with food and water ad libitum. The light cycle was automatically controlled (on at 7:00 AM and off at 7:00 PM), and the room temperature was thermostatically regulated to 22 ± 1°C. Before the experiments, animals were housed in these conditions for 3 to 4 days to become acclimatized. Some rats were adrenalectomized or sham-operated under ether anesthesia and used 3 to 4 days after the surgical procedure. Adrenalectomized animals received isotonic saline as drinking water until use. Animal care was in accordance with Italian and European regulations on the protection of animals used for experimental and other scientific purposes.
Carrageenin-Induced Pleurisy.
Rats were slightly anesthetized with ether, and 0.2 ml of 1% λ-carrageenin, suspended in sterile saline solution, was injected into the pleural cavity. Roxithromycin, clarithromycin, erythromycin, and azithromycin, suspended in olive oil, were administered p.o. via gastric gavage (0.5 ml/rat) at 10, 20, and 40 mg/kg 1 h before carrageenin injection. In some experiments, the macrolides were administered to adrenalectomized rats. The control groups received an equal volume of the vehicle. Carrageenin pleurisy was also induced in some rats treated with indomethacin (5 mg/kg) dissolved in carboxymethylcellulose and administered p.o. 1 h before the phlogogenic agent. Four hours after the induction of pleurisy, animals were sacrificed in an atmosphere of CO2. Pleural exudate from each animal was harvested by washing the pleural cavity with 2 ml of sterile saline solution containing 5 U/ml heparin and 10 μg/ml of indomethacin. Exudates with blood contamination were rejected. The exudate volumes were measured, the samples were centrifuged at 800g for 10 min, and the cell pellet was resuspended in saline for total and differential cell count. Total cell count was estimated after trypan blue staining with the use of the Burker counting chamber. In some experiments, differential cell count was determined by May-Grunwald Giemsa staining. The supernatants were then aliquoted and stored at −80°C until use.
Cell Culture.
The murine monocyte/macrophage cell line J774 was from the European Collection of Animal Cell Cultures (Salisbury, UK). J774 cells were grown in Dulbecco's modified Eagle's medium and cultured at 37°C in humidified 5% CO2/95% air. Culture medium was supplemented with 10% fetal bovine serum, 2 mM l-glutamine, 25 mM HEPES, 100 U/ml penicillin, 100 μg/ml streptomycin, and 5 mM sodium pyruvate at 37°C. The cells were plated onto 24-well culture plates (Falcon, Meylan, France) at a density of 2.5 × 105/ml and allowed to adhere for 2 h. Thereafter, the medium was replaced with fresh medium, and cells were activated by 1 μg/ml LPS in the presence or absence of various concentrations (5–80 μM) of the antibiotics. Indomethacin (1 μM) andNG-monomethyl-l-arginine (l-NMMA; 30 μM) have been used as reference drugs. In some experiments, roxithromycin and erythromycin were added to the cells 12 h after LPS. At different time points (3, 8, and 24 h), according to the metabolite or the cytokine to be measured, culture medium was removed and centrifuged, and the supernatant was aliquoted and stored at −80°C until use. Cell viability (>95%) was determined with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide assay (Denizot and Lang, 1986).
NO3− plus NO2−(NOx) Assay.
The amount of NOx, stable metabolites of NO, present in the supernatant of inflammatory exudate was determined according to Thomsen et al. (1991). After reducing nitrate (NO3−) to nitrite (NO2−) with the use of acid-washed cadmium powder, NO2− amounts were measured according to a microplate assay method based on the Griess reaction, and the results are expressed as micrograms per rat. Nitrite levels in culture media from J774 macrophages were measured 24 h after LPS (1 μg/ml) stimulation with the Griess reaction as previously described (Di Rosa et al., 1990). Results are expressed as μg/ml and represent the mean ± S.E. of n experiments performed in triplicate.
Radioimmunoassay of PGE2, 6-Keto-PGF1α, and LTB4.
PGE2 and LTB4 in the supernatant of centrifuged exudate (800g for 10 min) were assayed by radioimmunoassay according to the procedures described by Granström and Kindhal (1978) andSalmon et al. (1982), respectively. The results are expressed as nanograms per rat and represent the mean ± S.E. of nrats. The cross-reactivity for the PGE2 rabbit antiserum was 3.4% for PGF2α, 2.1% for PGD2, and 2.0% for PGA2. The cross-reactivity for LTB4 rabbit antiserum was <0.1% for LTA4, <0.1% for LTC4, <0.1% for LTD4, and <0.1% for LTE4. The accumulation of 6-keto-PGF1α in the cell culture medium was measured, without prior extraction or purification, by radioimmunoassay (Maclouf, 1982). The anti-6-keto-PGF1α rabbit antibody showed cross-reactivity of 11% for PGE2, 10% for PGF2α, 3% for PGD2, and <0.5% for thromboxane B2. Results are expressed as pg/ml of 6-keto-PGF1α and represent the mean ± S.E. of n experiments performed in triplicate.
Assay for Cytokines.
TNF-α levels in the supernatant of centrifuged exudate (800g for 10 min) were measured with an ELISA kit according to the manufacturer's instructions, and the results are expressed as nanograms per rat. TNF-α, IL-1β, and IL-6 levels in the cell culture medium were assayed by using a commercially available mouse cytokine ELISA test kit according to the manufacturer's instructions, and the results are expressed as ng/ml and represent the mean ± S.E. of n experiments performed in triplicate.
Preparation of Cytosolic Fraction.
Extracts of unstimulated or LPS-stimulated (1 μg/ml for 24 h) J774 macrophages in the absence or presence of 80 μM roxithromycin or erythromycin were prepared as previously described (Schreiber et al., 1989). Briefly, harvested cells (2 × 107) were washed twice with ice-cold PBS and centrifuged at 180g for 10 min at 4°C. The cell pellet was resuspended in 100 μl of ice-cold hypotonic lysis buffer (10 mM HEPES, 1.5 mM MgCl2, 10 mM KCl, 0.5 mM phenylmethylsulfonyl fluoride, 1.5 μg/ml soybean trypsin inhibitor, 7 μg/ml pepstatin A, 5 μg/ml leupeptin, 0.1 mM benzamidine, 0.5 mM dithiothreitol) and incubated in ice for 15 min. The cells were lysed by five or six rapid passages through a 20-gauge needle; the cytoplasmic fraction was obtained through centrifugation at 13,000g for 1 min; and supernatant was aliquoted and stored at −80°C.
Western Blot Analysis.
Immunoblotting analysis of COX-2, iNOS, and β-actin proteins was performed on cytosolic fraction. Cytosolic fraction proteins were mixed with gel loading buffer [50 mM Tris, 10% SDS, 10% glycerol, 10% 2-mercaptoethanol, and 2 mg bromophenol (ml−1)] at a ratio of 1:1, boiled for 3 min, and centrifuged at 10,000g for 10 min. Protein concentration was determined according to the manufacturer's instructions (Bio-Rad), and equivalent amounts of protein (75 μg) from each sample were electrophoresed in an 8% discontinuous polyacrylamide minigel. The proteins were transferred onto nitrocellulose membranes, according to the manufacturer's instructions. The membranes were saturated by incubation at 4°C overnight with 10% nonfat dry milk in PBS and then incubated with anti-COX-2 (1:250), anti-iNOS (1:10,000), or anti-β-actin (1:1000) mouse antibodies for 2 h at room temperature. The membranes were washed three times with 1% Triton X-100 in PBS and then incubated with anti-mouse immunoglobulins coupled to peroxidase (1:2000). The immune complexes were visualized by the enhanced chemiluminescence method (Amersham).
Statistical Analysis.
Values are expressed as the mean ± S.E. of n animals for in vivo experiments and ofn experiments run in triplicate for in vitro experiments. Comparisons were calculated by one-way ANOVA and Bonferroni-correctedP value for multiple comparisons. The level of statistically significant difference was defined as P < .05.
Results
Carrageenin Pleurisy
Exudate Volume and Cell Migration.
The injection of 0.2 ml of 1% λ-carrageenin into the pleural cavity of rats caused an inflammatory reaction characterized by exudate formation and cell migration (Table 1). In control animals (carrageenin only), the average volume of the exudate at 4 h was 0.70 ± 0.03 ml/rat (n = 35), and the total leukocyte number (>95% neutrophils) that migrated into the pleural cavity was 126.6 ± 5.6 × 106/rat (n = 35). Treated rats received the macrolides at 10, 20, or 40 mg/kg p.o. 1 h before carrageenin injection. The treatment was ineffective when the antibiotics were administered at 10 mg/kg, whereas the inflammatory reaction was inhibited when higher doses were used. Roxithromycin at 20 and 40 mg/kg dose dependently and significantly reduced the exudate volume by 36% (P < .01, n = 8) and 50% (P < .001,n = 7), respectively, whereas the total number of cells that migrated into the pleural cavity was decreased by 20% (P < .05, n = 8) and 30% (P < .01, n = 7), respectively. Only when administered at 40 mg/kg, clarithromycin and erythromycin significantly reduced the volume of the exudate by 43% (P < .01, n = 9) and 50% (P < .001, n = 8), respectively, and the number of leukocytes that migrated was reduced by 30% (P < .01, n = 9) and 32% (P < .01, n = 8), respectively (Table1). Azithromycin (10–40 mg/kg) had no effect on the exudate volume, whereas it significantly inhibited by 19% (P < .05,n = 8) the number of leukocytes only at 40 mg/kg. Indomethacin (5 mg/kg p.o.) reduced the exudate volume by 67% (P < .001, n = 8) and the number of leukocytes by 31% (P < .05, n = 8; Table 1).
Effect of Macrolide Antibiotics and Indomethacin on Rat Carrageenin-Induced Pleurisy
In adrenalectomized animals, the inflammatory reaction induced by carrageenin, compared with control rats, greatly enhanced the exudate volume (2.25 ± 0.10 ml/rat; P < .0001,n = 12) and the total number of migrated leukocytes (342.5 ± 23.2 × 106 cells/rat;P < .0001, n = 12), whereas in sham-operated animals (0.74 ± 0.04 ml of exudate and 123.4 ± 6.3 × 106 cells, n = 5), it was superimposable on that observed in normal rats. Roxithromycin at 20 and 40 mg/kg dose dependently reduced the exudate volume by 38% (1.25 ± 0.09 ml, P < .001, n = 5) and 49% (1.00 ± 0.11 ml, P < .001,n = 5), respectively, and the total number of migrated leukocytes by 22% (229.3 ± 11.5 × 106 cells, P < .001,n = 5) and 35% (222.5 ± 3.1 x106 cells, P < .001,n = 5), respectively. Clarithromycin and erythromycin at 40 mg/kg significantly reduced the exudate volume by 45% (1.11 ± 0.04 ml, P < .001, n = 4) and 48% (1.05 ± 0.02 ml, P < .001, n = 5), respectively, and the number of migrated leukocytes by 29% (208.5 ± 5.3 × 106 cells,P < .001, n = 4) and 31% (203.3 ± 3.5 × 106 cells, P < .001, n = 5), respectively. Azithromycin (40 mg/kg) had no effect on exudate volume (2.31 ± 0.10 ml), whereas it significantly inhibited the leukocyte number by 21% (233.4 ± 7.5 × 106 cells, P < .01,n = 4).
NOx, PGE2, and LTB4.
The pleural exudate of control rats contained detectable amounts of NOx (1.51 ± 0.06 μg/rat,n = 35), PGE2 (0.428 ± 0.01 ng/rat, n = 35), and LTB4(2.5 ± 0.15 ng/rat, n = 35). The amounts of both NOx and PGE2 were virtually unaffected by macrolides administered at 10 mg/kg, whereas they were reduced when higher doses were used (Table 1). Roxithromycin at 20 and 40 mg/kg caused a dose-dependent inhibition of NOx, which was reduced by 34% (P< .001, n = 8) and 50% (P < .001,n = 7), respectively, whereas PGE2 was decreased by 21% (P < .05, n = 8) and 41% (P < .001,n = 7), respectively. Clarithromycin and erythromycin at 20 and 40 mg/kg reduced NOx by 22% (P < .05, n = 9) and 42% (P < .001, n = 9) and by 30% (P < .01, n = 8) and 44% (P < .001, n = 8), respectively, whereas PGE2 was significantly reduced by 23% (P < .05, n = 9) and 43% (P < .001, n = 9) and by 39% (P < .001, n = 8) and 53% (P < .001, n = 8), respectively. Azithromycin up to 20 mg/kg did not modify either NOx or PGE2, whereas at 40 mg/kg, it significantly reduced NOx by 30% (P < .05, n = 8) and PGE2 by 26% (P < .05,n = 8).
All of the macrolide antibiotics used in this study had no effect on the amount of LTB4 in pleural exudates. Indomethacin (5 mg/kg p.o.) reduced NOx and PGE2 by 35% (P < .05,n = 8) and 87% (P < .001,n = 8), respectively, whereas it did not modify LTB4 production.
TNF-α.
The pleural exudate of control animals contained 2.14 ng of TNF-α/rat (n = 35; Table 1). The amounts of TNF-α were unaffected by macrolides administered at 10 mg/kg, whereas they were reduced when higher doses were used. Roxithromycin at 20 and 40 mg/kg caused a dose-dependent inhibition of TNF-α, which was reduced by 38% (P < .05, n = 8) and 47% (P < .01, n = 7), respectively. Clarithromycin and erythromycin at 20 and 40 mg/kg inhibited TNF-α production by 28% (P < .05, n = 9) and 47% (P < .01, n = 9) and by 29% (P < .05, n = 8) and 52% (P < .01, n = 8), respectively. Azithromycin significantly inhibited TNF-α production by 36% (P < .01, n = 8) only at the highest dose used (40 mg/kg).
J774 Murine Macrophages
In preliminary experiments, we established that cell viability (>95%) was not affected by any of the four macrolides (up to 80 μM), 1 μM indomethacin, or 30 μM l-NMMA (data not shown).
TNF-α.
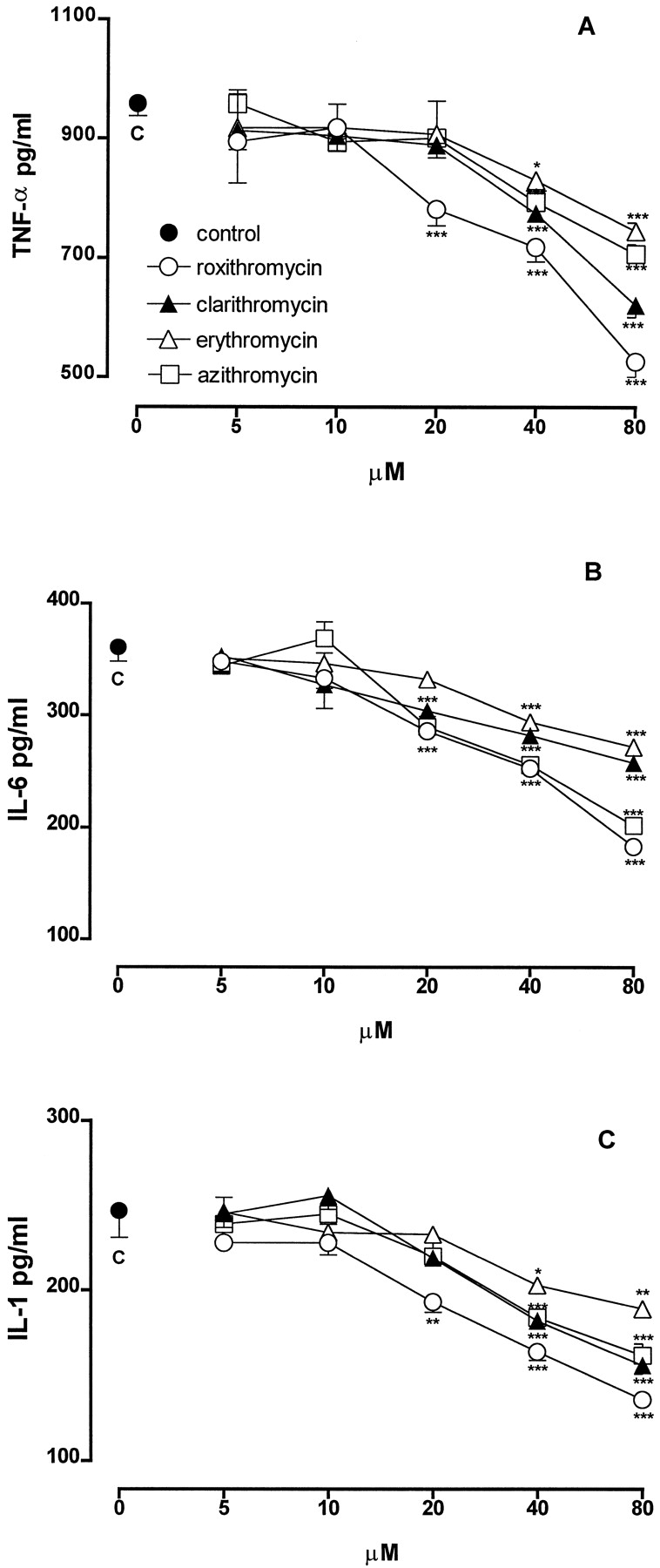
The production of TNF-α by unstimulated J774 cells was <15 pg/ml (n = 6). Incubation of these cells with LPS (1 μg/ml) for 3 h caused a substantial increase in TNF-α production (958 ± 20 pg/ml; n = 6). When J774 macrophages were stimulated with the same amount of LPS in the presence of macrolide antibiotics (5–80 μM), a concentration-dependent inhibition of TNF-α production was observed (Fig.1A). Roxithromycin, which was ineffective at 5 and 10 μM, at 20, 40, and 80 μM inhibited TNF-α production by 18, 25, and 45%, respectively (P < .001,n = 3–6). Clarithromycin, erythromycin, and azithromycin were ineffective up to 20 μM, whereas at 40 and 80 μM, they inhibited TNF-α production by 19 and 35% (P < .001, n = 3–6), by 13% (P < .05,n = 6) and 22% (P < .001,n = 3), and by 17 and 26% (P < .001,n = 3–6), respectively.
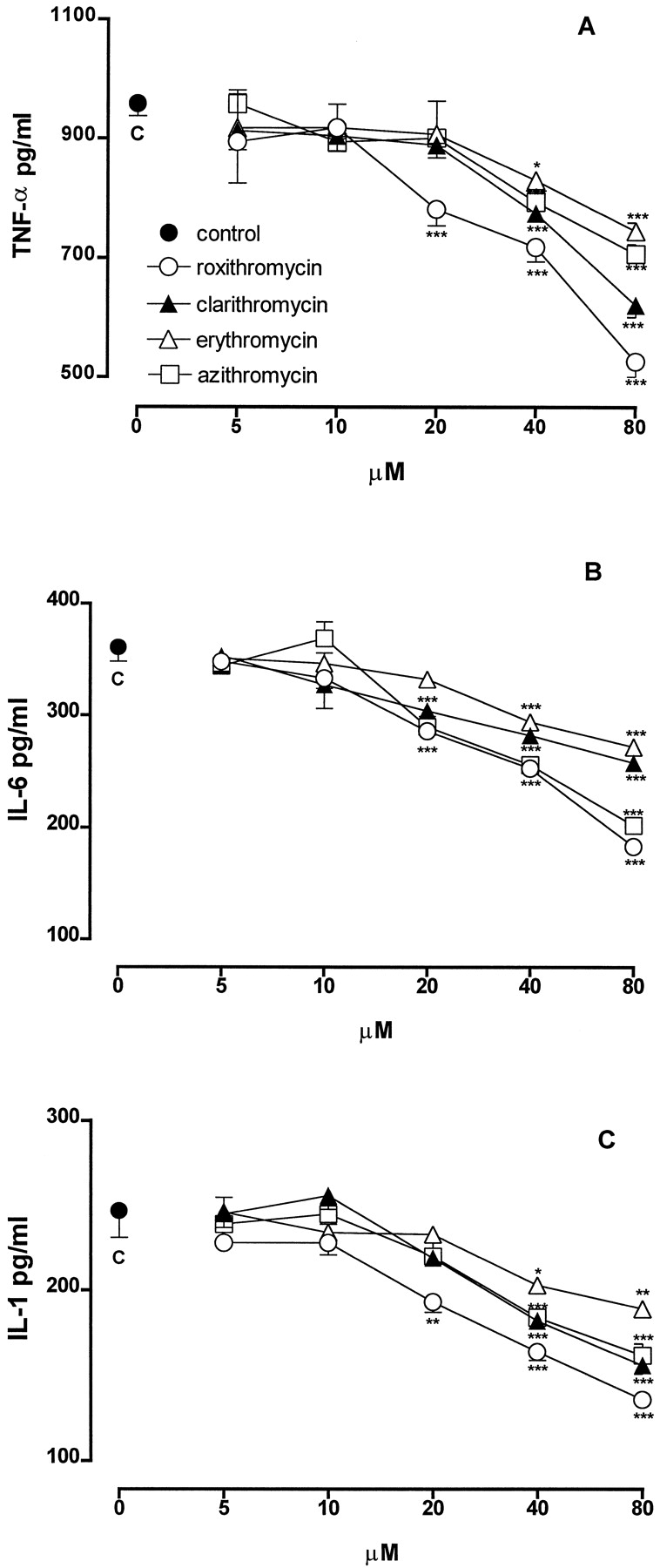
Effect of different concentrations of macrolides on the production of TNF-α (A), IL-6 (B), and IL-1β (C) by J774 macrophages stimulated with LPS (1 μg/ml). Each point represents the mean ± S.E. of three separate experiments run in triplicate. *P < .05; **P < .01; ***P < .001 versus control (LPS alone).
IL-6.
The treatment of J774 macrophages with LPS for 8 h greatly increased the production of IL-6 (361 ± 13 pg/ml;n = 6) compared with the release of unstimulated cells (<15 pg/ml; n = 6). All of the macrolides were ineffective at 5 and 10 μM (Fig. 1B). Roxithromycin at 20, 40, and 80 μM significantly inhibited IL-6 production by 21, 30, and 49% (P < .001, n = 6), respectively. Clarithromycin and azithromycin at 20, 40, and 80 μM inhibited IL-6 production by 16, 22, and 29% (P < .001,n = 6) and by 20, 29, and 44% (P < .001, n = 6), respectively. Erythromycin was ineffective at 20 μM, whereas it inhibited significantly IL-6 production at 40 and 80 μM by 25 and 31% (P < .001,n = 6), respectively.
IL-1β.
The stimulation of J774 macrophages with LPS for 24 h caused an increased release of IL-1β (247 ± 16 pg/ml;n = 6) compared with the release of unstimulated cells (<15 pg/ml; n = 6). When J774 were stimulated with LPS in the presence of macrolides (5–80 μM), a concentration-related inhibition of IL-1β production was observed (Fig. 1C). At 5 and 10 μM, all antibiotics were ineffective. Roxithromycin at 20, 40, and 80 μM significantly inhibited IL-1β generation by 22% (P < .01, n = 6), 34%, and 45% (P < .001, n = 6) respectively. Clarithromycin, erythromycin, and azithromycin were ineffective up to 20 μM, whereas at 40 and 80 μM, they significantly reduced IL-1β levels by 26 and 37% (P < .001, n = 6), by 18% (P < .05, n = 6) and 24% (P < .001, n = 6), and by 26 and 35% (P < .001, n = 6), respectively.
6-Keto-PGF1α Production and COX-2 Expression.
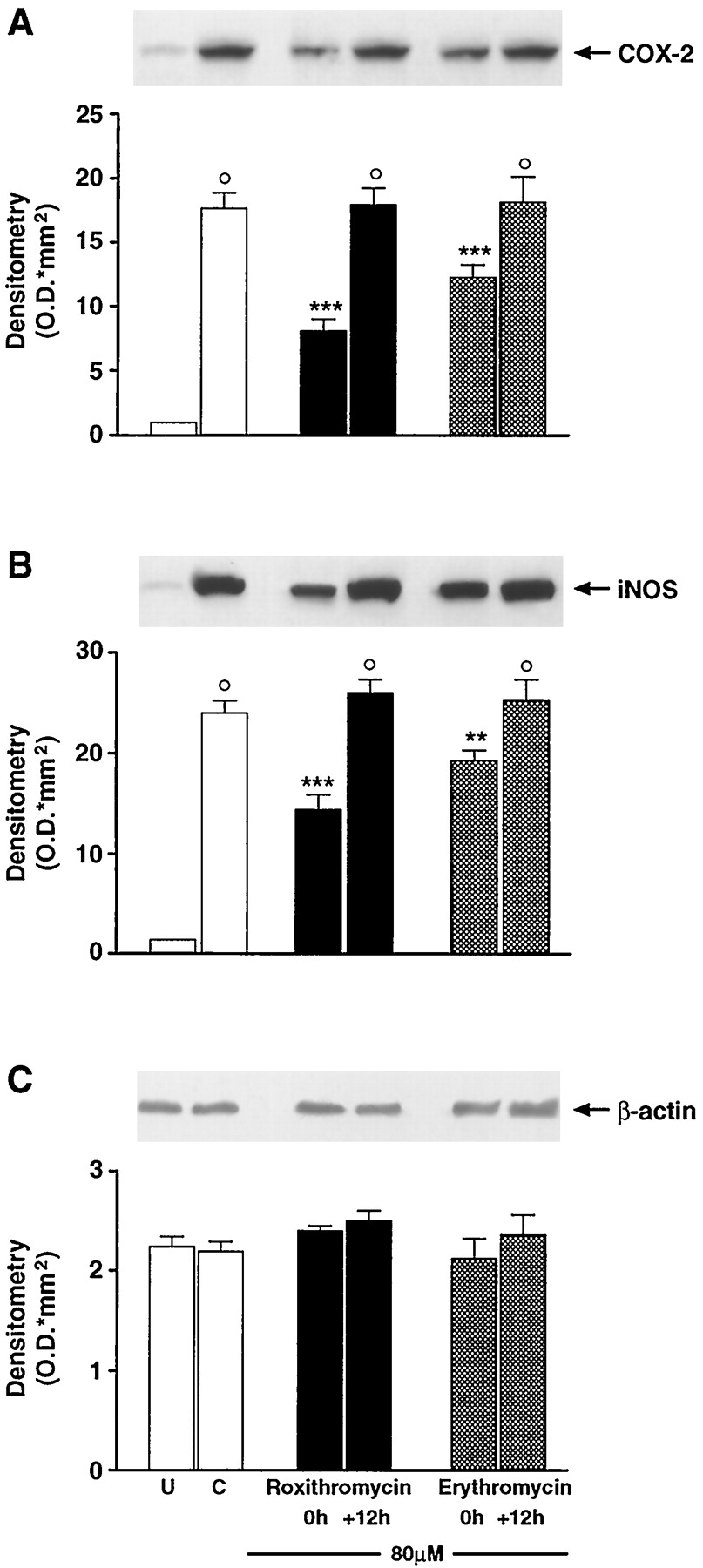
In 24 h, unstimulated J774 macrophages generated 30 ± 1 pg/ml 6-keto-PGF1α (n = 5). Stimulation of the cells with bacterial LPS produced a significant (P < .001) increase in the production of this prostanoid (350 ± 10 pg/ml, n = 6). When the cells were stimulated in the presence of macrolides (5–80 μM), a concentration-dependent inhibition of 6-keto-PGF1α generation was observed (Fig.2A). At 5 μM, all of the antibiotics were ineffective. Roxithromycin (10–80 μM) significantly inhibited 6-keto-PGF1α generation by 14% (P < .05, n = 4), 20% (P < .01, n = 4), 37%, and 47% (P < .001, n = 4) respectively. Clarithromycin and azithromycin (10–80 μM) exhibited a similar pattern of action because they significantly inhibited 6-keto-PGF1α generation by 13% (P < .05, n = 4), 15% (P < .05, n = 3), 23 and 38% (P < .001, n = 4), and 22% (P < .01, n = 4), 34%, 40% and 45% (P < .001, n = 3–5), respectively. Erythromycin was ineffective up to 20 μM, whereas it inhibited 6-keto-PGF1α generation at 40 and 80 μM by 15% (P < .01, n = 5) and 34% (P < .001, n = 5), respectively. Indomethacin (1 μM) inhibited 6-keto-PGF1αproduction by 85% (P < .001, n = 4; data not shown). Interestingly, when 80 μM roxithromycin or erythromycin was added to the incubation medium 12 h after LPS, there was no effect on 6-keto-PGF1α generation (Fig. 2B). Moreover, the stimulation of the cells with LPS resulted in an increase of COX-2 protein expression. As demonstrated in immunoblotting experiments, LPS-induced COX-2 protein expression was greatly reduced by coincubation with roxithromycin or erythromycin (80 μM) only when the antibiotics were administered concomitantly with LPS challenge, whereas it was unaffected when the macrolides were administered 12 h later (see Fig. 4A).

A, effect of different concentrations of macrolides on the production of 6-keto-PGF1α by J774 macrophages stimulated with LPS (1 μg/ml). Each point represents the mean ± S.E. of three separate experiments run in triplicate. *P < .05; **P < .01; ***P < .001 versus control (LPS alone). B, effect of delayed addiction of roxithromycin and erythromycin on 6-keto-PGF1α production by J774 macrophages stimulated with LPS (1 μg/ml). Roxithromycin (80 μM) and erythromycin (80 μM) were added to the cells at the same time (0 h) or 12 h after (+12 h) LPS challenge. Each column represents the mean ± S.E. of three separate experiments run in triplicate. ***P< .001 versus control (C, LPS alone).

Effect of delayed addiction of roxithromycin and erythromycin on COX-2 (A), iNOS (B), and β-actin (C) in J774 macrophages stimulated with LPS (1 μg/ml). Roxithromycin (80 μM) and erythromycin (80 μM) were added to the cells at the same time point (0 h) or 12 h after (+12 h) LPS challenge. Western blot analysis is referred to a single experiment representative of three separate experiments. Each column represents the mean ± S.E. of three separate experiments. °P < .001 versus unstimulated cells (U). **P < .01; ***P < .001 versus control cells (C, LPS alone).
NO2− Production and iNOS Expression.
The production of NO2− by unstimulated J774 cells was undetectable (<50 ng/ml; n= 4). Incubation of the cells with LPS caused a substantial release of NO2− (989 ± 83 ng/ml;n = 9). When J774 were stimulated with LPS in presence of macrolides (5–80 μM), a concentration-related inhibition of NO2− generation was observed (Fig. 3A). At 5 and 10 μM, all antibiotics were ineffective. Roxithromycin at 20, 40, and 80 μM significantly inhibited NO2−release by 24% (P < .01, n = 4), 36%, and 58% (P < .001, n = 4), respectively. Clarithromycin and azithromycin (20–80 μM) inhibited NO2− generation by 15% (P < .05, n = 5), 24% (P < .01, n = 4), and 39% (P < .001, n = 4) and by 25% (P < .01, n = 3), 37%, and 50% (P < .001, n = 5), respectively. Erythromycin only at 40 and 80 μM inhibited NO2− generation by 17% (P < .05, n = 5) and 27% (P < .001, n = 5), respectively.l-NMMA (30 μM) significantly inhibited NO2− generation by 49% (P < .01, n = 4; data not shown). The addition of 80 μM roxithromycin or erythromycin to the cells 12 h after LPS challenge did not significantly affect NO2− production (Fig. 3B).The stimulation of the cells with LPS resulted in an increase in iNOS protein expression as demonstrated by immunoblotting experiments (Fig.4B). The LPS-induced iNOS protein expression was prevented by coincubation with roxithromycin or erythromycin (80 μM) only when these macrolides were administered concomitantly with LPS, whereas they had no effect when administered 12 h later.

A, effect of different concentrations of macrolides on the production of NO2− by J774 macrophages stimulated with LPS (1 μg/ml). Each point represents the mean ± S.E. of three separate experiments run in triplicate. *P < .05; **P < .01; ***P < .001 versus control (LPS alone). B, effect of delayed addiction of roxithromycin and erythromycin on NO2− production by J774 macrophages stimulated with LPS (1 μg/ml). Roxithromycin (80 μM) and erythromycin (80 μM) were added to the cells at the same time (0 h) or 12 h after (+12 h) LPS challenge. Each column represents the mean ± S.E. of three separate experiments run in triplicate. ***P< .001 versus control (C, LPS alone).
β-Actin Expression.
The expression of one of the major cytoskeleton filament, β-actin, was analyzed by Western blotting for comparative purposes. In either unstimulated or LPS-stimulated J774 cells, the expression of such a protein remained unchanged. Furthermore, the β-actin expression was not affected by coincubation with roxithromycin or erythromycin (80 μM) administered either concomitantly or 12 h after the LPS challenge.
Discussion
In this study, the anti-inflammatory activity of four macrolide antibiotics, roxithromycin, clarithromycin, erythromycin, and azithromycin, and their ability to reduce the production of proinflammatory mediators and cytokines have been investigated both in vivo and in vitro. Rat carrageenin pleurisy was used as a model of acute inflammation, and the inflammatory reaction was induced in either normal or adrenalectomized animals. The macrolides were administered p.o. 1 h before the carrageenin challenge. Roxithromycin, clarithromycin, and erythromycin (10, 20, and 40 mg/kg) dose dependently reduced both exudate volume and leukocyte accumulation in normal rats. Rat carrageenin pleurisy is controlled by endogenous steroid because in adrenalectomized rats, the inflammatory reaction was much greater compared with that occurring in either normal or sham-operated animals. The doses of roxithromycin (20 and 40 mg/kg), clarithromycin (40 mg/kg), and erythromycin (40 mg/kg), which significantly reduced exudate volume and leukocyte accumulation in normal rats, exhibited an identical inhibitory effect in adrenalectomized rats, showing that the anti-inflammatory activity of macrolides was not dependent on the stimulation of endogenous corticoid production. Furthermore, in normal rats, the amounts of PGE2, NOx, and TNF-α in pleural exudates were significantly reduced by these macrolides. Roxithromycin appeared more effective than erythromycin and clarithromycin, whereas azithromycin only slightly affected the inflammatory reaction. None of the macrolides were able to modify LTB4 production, suggesting that a nonspecific effect on inflammatory mediators could be ruled out. The mechanism of the anti-inflammatory activity of macrolide antibiotics is unclear. Although it has been shown that macrolides exhibit membrane-stabilizing effects in human neutrophils and inhibit superoxide anion generation by these cells stimulated with formyl-methionyl-leucyl-phenylalanine or the calcium ionophore A23187, further studies should be carried out to clarify whether these antibiotics are able to affect some early events (e.g., shape modification, calcium mobilization, intracellular pH change) occurring in leukocyte activation. However, the inhibition of the prostanoid pathway may contribute, at least in part, to the anti-inflammatory effect of macrolides, although it has been shown that these agents suppress inflammation through mechanisms different from conventional nonsteroidal anti-inflammatory drugs (Tarayre et al., 1987; Agen et al., 1993). One of these mechanisms may be the inhibition of the l-arginine: NO pathway, as shown by the reduction in NOx, stable metabolites of NO in the pleural exudate. NO is involved in several types of acute and chronic inflammation (Ialenti et al., 1992, 1993), including rat carrageenin pleurisy (Sautebin et al., 1998) and zymosan-induced peritonitis in mice (Ajuebor et al., 1998), which, interestingly, is inhibited by erythromycin (Mikasa et al., 1992). The reduction in TNF-α in rat pleural exudate is in agreement with previous reports showing that the systemic administration of macrolides in animals and humans down-regulates the production of proinflammatory cytokines, including TNF-α and IL-1β (Konno et al., 1994; Kadota et al., 1996). Our in vitro experiments have shown that the four macrolides (5–80 μM) reduced in a concentration-dependent manner the production of TNF-α, IL-1β, IL-6, 6-keto-PGF1α, and NO2− production by LPS-stimulated J774 macrophages. These data are in agreement with the results of in vivo experiments and further support the ability of macrolides to inhibit the production of inflammatory mediators and cytokines. The suppressive effect of macrolides on the production of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β has been extensively studied in vitro. Roxithromycin has been shown to inhibit the production of IL-1β and TNF-α in human peripheral blood monocytes in a dose-dependent manner (Yoshimura et al., 1995). Clarithromycin showed an inhibitory effect on cytokine production (IL-6 and IL-1β) by synovial fibroblast-like cells (Matsuoka et al., 1996), and erythromycin inhibited TNF-α release by human monocytes stimulated with LPS (Iino et al., 1992). Because in rat carrageenin pleurisy and in LPS-stimulated J774 macrophages the inducible isoforms of both COX-2 and iNOS are expressed (Tomlison et al., 1994; Katori et al., 1995; D'Acquisto et al., 1997, 1998), the possibility that the reduction in prostanoid and NO metabolites levels could depend on the ability of macrolides to prevent the expression of both COX-2 and iNOS has been investigated. We have shown that in J774 cells, the inhibition of 6-keto-PGF1α and NO2− production by roxithromycin and erythromycin was not dependent on direct inhibition of COX-2 and iNOS activity because both macrolides, when added to the cells 12 h after LPS challenge, did not affect the enzyme catalytic activity. Moreover, the reduction in 6-keto-PGF1α and NO2− production seems to be related to the inhibition of COX-2 and iNOS protein expression by these macrolides. Thus, both antibiotics, when added to the cells concomitantly with LPS, greatly reduced the levels of COX-2 and iNOS protein expression, whereas they did not affect the expression of β-actin, a major cytoskeleton filament (i.e., a protein unrelated to inflammation). This is the first demonstration, to our knowledge, that roxithromycin and erythromycin inhibit the expression of enzymes involved in inflammation. This property, which may be shared by other macrolides, represents a relevant mechanism underlying the anti-inflammatory effect of these antibiotics. It is well known that the expression of several genes involved in the immune and inflammatory response (e.g., iNOS, COX-2, TNF-α, IL-1, IL-6) is regulated at the transcriptional level by the nuclear factor-κB (NF-κB) (Müller et al., 1993; Xie et al., 1994; Yamamoto et al., 1995). It has been recently shown that the antioxidant pyrrolidinedithiocarbamate is able to reduce COX-2 expression and prostaglandin production in LPS-stimulated J774 macrophages, suggesting an involvement of NF-κB in the induction of COX-2 (D'Acquisto et al., 1997). We have recently shown that NF-κB is activated in rat carrageenin-induced pleurisy and that its activation is inhibited by antioxidant agents, leading to a reduction in the inflammatory reaction (D'Acquisto et al., 1999). Erythromycin and roxithromycin exhibit antioxidant properties (Miyachi et al., 1986; Labro et al., 1989; Hand et al., 1990); thus, it is conceivable that these, and perhaps other macrolides, may act as anti-inflammatory agents by preventing the activation of NF-κB. Although this hypothesis requires to be supported by further experimental work, it appears of particular interest in the light of recent findings demonstrating that patients with unstable angina treated with roxithromycin showed a significant reduction in major ischemic events compared with the placebo group (Gurfinkel et al., 1997). Because there is serological evidence for an association between Chlamydia pneumoniae and coronary heart disease, the beneficial effect of roxithromycin may be related to its antichlamydial activity. However, due to its anti-inflammatory activity, roxithromycin may attenuate the persistent inflammation in the atherosclerotic plaque by reducing the release of proinflammatory mediators and cytokines through the inhibition of NF-κB activation, which has been shown to play a relevant role in the pathogenesis of the atherosclerotic lesion (Lindner and Collins, 1996).
In conclusion, the present study shows that macrolide antibiotics have anti-inflammatory activity, which likely depends on their ability to prevent the production of proinflammatory mediators and cytokines, and suggests that these agents, particularly roxithromycin, can exert therapeutic effects independent of their antibacterial activity.
Footnotes
-
Send reprint requests to: Prof. Massimo Di Rosa, Department of Experimental Pharmacology, University of Naples “Federico II,” Via D. Montesano, 49, 80131, Naples, Italy. E-mail: dirosa{at}unina.it
- Abbreviations:
- LPS
- lipopolysaccharide
- LTB4
- leukotriene B4
- PGE2
- prostaglandin E2
- ELISA
- enzyme-linked immunosorbent assay
- 6-keto-PGF1α
- 6-keto-prostaglandin F1α
- COX-2
- cyclooxygenase-2
- NO
- nitric oxide
- iNOS
- inducible nitric oxide synthase
- IL-1β
- interleukin-1β
- IL-6
- interleukin-6
- NF-κB
- nuclear factor-κB
- TNF-α
- tumor necrosis factor-α
- NO3−
- nitrate
- NO2−
- nitrite
- NOx
- nitrate plus nitrite
- l-NMMA
- NG-monomethyl-l-arginine
- Received May 10, 1999.
- Accepted September 6, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}