





Abstract
Poly (ADP-ribose) synthase or polymerase (PARS and PARP, respectively) is a cytotoxic enzyme which causes cellular damage. Nicotinamide, a compound of vitamin B complex, has been reported to exert an inhibitory effect on PARS or PARP. The present study tests the effects of nicotinamide on acute lung injury and associated alterations following ischaemia/reperfusion (I/R) of the isolated perfused rat's lung.
I/R increased the lung weight (LW) to body weight ratio, LW gain, protein and dye tracer leakage, pulmonary arterial pressure and capillary permeability. The insult also increased nitrate/nitrite, methyl guanidine, tumour necrosis factor-α and interleukin-1β in lung perfusate, while it decreased adenosine triphosphate content with an increase in PARP activity in lung tissue.
Most of the I/R-induced changes were abrogated by post-treatment (30 min after I/R) with nicotinamide (100 mg·kg−1 body weight). However, the increase in pulmonary arterial pressure was enhanced by nicotinamide post-treatment. Following I/R, the inducible nitric oxide synthase (iNOS) mRNA expression was enhanced. Nicotinamide reduced the iNOS expression.
The results suggest that nicotinamide exerted a protective effect on the acute lung injury caused by ischaemia/reperfusion. The mechanisms may be mediated through the inhibition on the poly (adenosine diphosphate-ribose) polymerase activity, inducible nitric oxide synthase expression and the subsequent suppression of nitric oxide, free radicals and pro-inflammatory cytokines with restoration of adenosine triphosphate.
- Free radical
- inducible nitric oxide synthase
- nitric oxide
- poly (ADP-ribose) polymerase
- pro-inflammatory cytokines
Acute lung injury (ALI) following ischaemia/reperfusion (I/R) has long been observed in various pulmonary disorders, such as pulmonary arterial thrombo-endarterectomy 1 and lung transplantation 2. Ischaemia causes lung injury through a temporary interruption of blood flow to the lungs followed by reperfusion, which is evidenced by increases in lung weight, microvascular permeability, pulmonary arterial pressure and lung pathology 3, 4. The current authors have provided evidence to indicate that endogenous and exogenous nitric oxide (NO) are detrimental to I/R lung injury 3, but that static inflation exerts protective effects 4.
Nicotinamide is a compound of the soluble B complex. It exerts an inhibitory effect on the poly (adenosine diphosphate (ADP)-ribose) synthase or polymerase (PARS and PARP, respectively). The nuclear enzyme can be activated by strand breaks in DNA caused by reactive oxygen and nitrogen species, and peroxynitrite 5, 6. PARP is cytotoxic due to massive depletion of intracellular nicotinamide adenine dinucleotide (NAD+) and adenosine triphosphate (ATP). Inhibition of PARP activity reduces the I/R injury of the heart, skeletal muscle and brain 6, 7. In addition, PARP inhibition with 3-aminobenzamide attenuates the acute lung injury induced by endotoxin 8. The inhibitory effects of nicotinamide or its related substances, niacinamide and nicotinic acid, on PARP activity are protective to organ dysfunction and/or cell damage caused by oxidative stress 9, 10. The current authors’ laboratory has reported that PARS or PARP inhibition with niacinamide attenuates the I/R hepatic injury 11.
Studies from the present authors’ laboratory have investigated the pathogenic mechanisms and potential therapeutic regimen on ALI induced by endotoxin 12, 13 and I/R 3, 4. The present study was designed to evaluate the protective effect of nicotinamide on ALI caused by I/R in isolated perfused lungs. The authors also aimed to elucidate the possible mechanisms of the protective role of nicotinamide on the lung changes. The current study’s results indicate that nicotinamide mitigates the ALI induced by I/R.
METHODS
Experimental models
Male rats of Sprague-Dawley strain, 12–15-weeks old and weighing 330–360 g were used. The animals were obtained from the National Animal Center (Nan-Kong, Taipei, Taiwan) and were housed in the University Laboratory Animal Center (Hualein, Taiwan) with adequate environmental control. The animal experiment was approved by the University Committee of Laboratory Animal Care and Use, and followed the guidelines of the National Animal Research Center. The room temperature was maintained at 21±1°C under a 12/12 h light/dark regimen. Food and water were provided ad libitum.
Acute experiments were carried out in isolated perfused rat's lungs. Rats were anaesthetised with an intraperitoneal injection of sodium pentobarbital (40 mg·kg−1). The rats were intubated with an endotracheal tube to provide ventilation of 95% room air–5% CO2 with a rodent ventilator. The respiratory rate and tidal volume were 60–65 breaths·min−1 and 2–3 mL (6–9 mL·kg−1), respectively.
Isolation and perfusation of the lungs in situ
Isolated perfused lungs were prepared according to the procedures described in previous studies 3, 4. A vertical incision was made along the midline of the thorax. Heparin (2 IU·g−1 body weight) was then injected into the right ventricle. An afferent silicon catheter was inserted into the pulmonary artery through the right ventricle. An efferent catheter was installed into the left atrium via the left ventricle. The pulmonary trunk and the aorta were tied. A third ligature was placed at the atrio-ventricular junction to prevent the perfusate from entering the left ventricle. The isolated lungs were perfused with Krebs-Henseleit balanced solution with 6% albumin. The perfusion system included a venous reservoir and a roller pump. The pulmonary venous outflow was diverted via the efferent catheter into the reservoir. The latter was placed in a 38°C water bath for constant temperature. Pulmonary arterial pressure (Ppa) and pulmonary venous pressure (Ppv) were measured from side ports in the afferent and efferent catheters. The lungs were perfused at constant flow (10–14 mL·min−1). Flow rate was adjusted to maintain the initial Ppa at 15–16 mmHg.
The isolated perfused lungs were left in situ and the rat was placed on an electronic balance. The digital signals of the electronic balance were converted to analogue signals by a digital–analogue converter and recorded on a polygraph recorder. Weight changes were pre-calibrated on the electronic balance before preparation for the experiment. In the isolated lung in situ preparation, the changes in body weight (BW) reflected the changes in lung weight (LW) 3, 4.
LW and LW gain
The initial LW was estimated from an equation relating to the BW 12, 13:
LW (g) = 0.0015×BW (g)+0.034 (1)
LW gain (LWG) was obtained from the increase in LW and also calculated as:
LWG = (final LW–initial LW)/initial LW (2)
Protein concentration in bronchoalveolar lavage
After the experiment, lungs were lavaged twice with saline (2.5 mL per lavage). Lavage samples were centrifuged at 1,500×g at room temperature for 10 min. The protein concentration in bronchoalveolar lavage (PC-BAL) was determined with a spectrophotometer by measuring the change in absorbance at 630 nm after the addition of bromocresol green 4.
Evans blue leakage
Evans blue dye (1 mg·kg−1) was added to the lung perfusate 5 min prior to the end of the experiment. The dye concentration in lung tissue was determined spectometrically as described by Thurston et al. 14.
Microvascular permeability
Microvascular permeability (Kfc), or capillary filtration coefficient, was calculated from the increase in LW that was produced by an elevation in Ppv. The Kfc was defined as the initial weight gain rate (g·min−1) divided by Ppv (10 cmH2O) and LW, and expressed as g·min−1·cmH2O−1·100 g−1. During the experiment, Ppv was elevated rapidly by 10 cmH2O for 7 min to measure Kfc. The hydrostatic challenge elicited a biphasic increase in LW; an initial rapid component followed by a slow and steady component. The slow component of the weight gain was plotted on a semi-log scale as a function of time. The capillary filtration rate was obtained by extrapolating the slow component of the weight gain back to zero time 3, 4.
Nitrate/nitrite, methyl guanidine, tumour necrosis factor-α and interleukin-1β in lung perfusate
Lung perfusate (0.5 mL) was taken 1 h before and 0.5, 1, 2, 3, 4, 5 and 6 h after I/R. The samples were centrifuged at 3,000×g for 10 min. The supernatant was used for nitrate/nitrite measurement with high-performance liquid chromatography (ENO-20; AD Instruments, Mountain View, CA, USA) 15. The formation of methyl guanidine (MG) has been identified as an index of hydroxyl radical production 16. It was determined with its fluorescence spectrum (Jasco 821-FP; Spectroscopic CO., Tokyo, Japan). The emission maximum was set at 500 nm and the excitation maximum at 398 nm. The assay was calibrated with authentic MG (M0377; Sigma Chemicals, St. Louis, MO, USA). Tumour necrosis factor (TNF)-α and interleukin (IL)-1β were measured with antibody ELISAs with a commercial antibody pair, recombinant standards and a biotin-streptavidin-peroxidase detection system (Endogen, Rockford, IL, USA). All agents, samples and working standards were prepared at room temperature according to the manufacturer's directions. The optical density was measured at 450/540 nm wavelengths by automated ELISA readers.
ATP content
Lungs were harvested after the experiments. A BioOrbit ATP Assay kit (Bio-Orbit Oy, Turku, Finland) was used to determine the lung ATP content with bioluminescence technique. The assay was based on quantitative measurement of a stable level of light as a result of an enzyme reaction catalysed by luciferase. Under the effect of luciferase, the luminescence evoked by ATP and luciferin interaction was recorded photometrically after amplification by a photomultiplier. The sensitivity of ATP was in a nanomolar range. The luciferin–luciferase reagent was used according to the manufacturer’s manual. ATP served as the standard. The test procedures have been described previously 11.
PARP activity
PARP activity in the harvested lung tissue was measured with a commercially available assay kit (Genzyme Diagnostics, Cambridge, MA, USA). Lung tissue samples were placed on ice in 2 mL buffer containing 50 mM Tris Cl (pH 8.0), 25 mM MgCl2 and 0.1 mM phenylmethylsulfonyl fluoride. The samples were homogenised for 30 s and then sonicated for 20 s using an ultrasonic homogeniser. The suspension was centrifuged at 3,000×g for 5 min at 4°C. Supernatant containing 20 μg protein, PARS buffer, 1 mM NAD, 2 μCi 32P-labelled NAD (1 μCi·μL−1) and distilled water was mixed in a microcentrifuge tube. The reaction was allowed to continue at room temperature for 1 min and was stopped by adding 900 μL of tricarboxylic acid. Enzyme activity was determined by measuring the incorporation of radiolabelled NAD as PARP catalysed the poly (ADP) ribosylation of proteins. The labelled ADP was determined by scintillation counting after tricarboxylic acid precipitation onto a filter. The procedures and calculation of PARP activity were carried out according to those described by Pulido et al. 17.
Inducible and endothelial NO synthases mRNA
RT-PCR was employed for a semiquantitative detection of inducible and endothelial NO synthase (iNOS and eNOS, respectively) mRNA expression in the lung tissue. Total cellular RNA was extracted from each tissue section after the experiment using the Trizol procedure (Gibco BRL, Gaithesburg, MD, USA). Primers specific for iNOS (sense: 5′-CTTCAGGTATGC-GGTATTGG-3′; antisense: 5′-CATGGTGAACACG-TTCTTGG-3′), eNOS (sense: 5′-AGCTGGCATGGGCAACTTGAA-3′; antisense: 5′-CAGCACATCAAAGCGGCCATT-3′), and reduced glyceraldehyde phosphate dehydrogenase (GAPDH; sense: 5′-TCCCTCAAGAT-TCTCAGCAA-3′; antisense: AGATCCACAACGGATACATT-3′) were used. Scanning densitometry was performed with an image scan and analysis system (Alpha-Innotech Corp., San Leandro, CA, USA). The current authors followed procedures described previously 18 and the manufacturer’s direction.
I/R
I/R was produced in the isolated perfused lungs using procedures similar to those described by Kao and co-workers 3, 4. In brief, lungs were initially ventilated with 5% CO2–95% N2 for 10 min to decrease the O2 content in the perfusate. Thereafter, ventilation and perfusion were stopped for 95 min. To facilitate subsequent reperfusion, the lungs were kept at a constant inflation state during the 95 min of ischaemia by holding ventilation at a steady inspiration state. The endotracheal pressure was set at 2 cmH2O. After ischaemia, the lungs were reperfused and ventilated with 95% room air and 5% CO2 for 50 min. The isolated perfused lungs were observed for 6 h after I/R.
Experimental protocols
In total, 30 isolated lung preparations were randomly subjected to vehicle and I/R with and without nicotinamide (NCA) post-treatment (n = 10 in each group). In isolated perfused lungs, NCA (100 mg·kg−1 BW) was added into the venous reservoir, from which LW changes, PC-BAL, Evans blue content, Kfc and Ppa were obtained. In lung perfusate, nitrate/nitrite, MG, TNF-α and IL-1β were determined, while ATP, PARP activity and NO synthase (NOS) mRNA were determined in lung tissue. NCA was purchased from Sigma Chemicals. It was dissolved in physiological saline solution before use and administered to the vehicle group. NCA was given 30 min after 95 min of ischaemia and 50 min of reperfusion. The dose of NCA administered into isolated lungs was in the range used in previous studies 11, 17, 19–21.
Data analysis
Data are expressed as means±sem. Comparisons within and among groups are made with ANOVA with repeated measures followed by post hoc comparison with Newman–Keul's test. A p-value <0.05 was considered statistically significant.
RESULTS
LW changes, Ppa and Kfc
In isolated lungs, I/R caused increases in LW/ BW ratio, LWG and Kfc. The changes in LW and capillary filtration were reduced by NCA post-treatment. I/R increased Ppa. Post-treatment with NCA augmented the I/R-induced pulmonary hypertension (fig. 1⇓).

The maximal values of a) lung weight (LW) to body weight (BW) ratio), b) LW gain (LWG), c) microvascular permeability (Kfc) and d) pulmonary arterial pressure (Ppa) in isolated lungs subjected to ischaemia/reperfusion (I/R) with (n = 10; ░) and without (n = 10; ▒) nicotinamide (NCA) post-treatment. □: vehicle (n = 10). I/R increased the LW change. The increases were abrogated by NCA post-treatment. I/R challenge elevated the Ppa, and the I/R-induced pulmonary hypertension was further augmented by NCA post-treatment. *: p<0.05 versus vehicle; #: p<0.05 versus I/R.
PC-BAL and Evans blue dye content
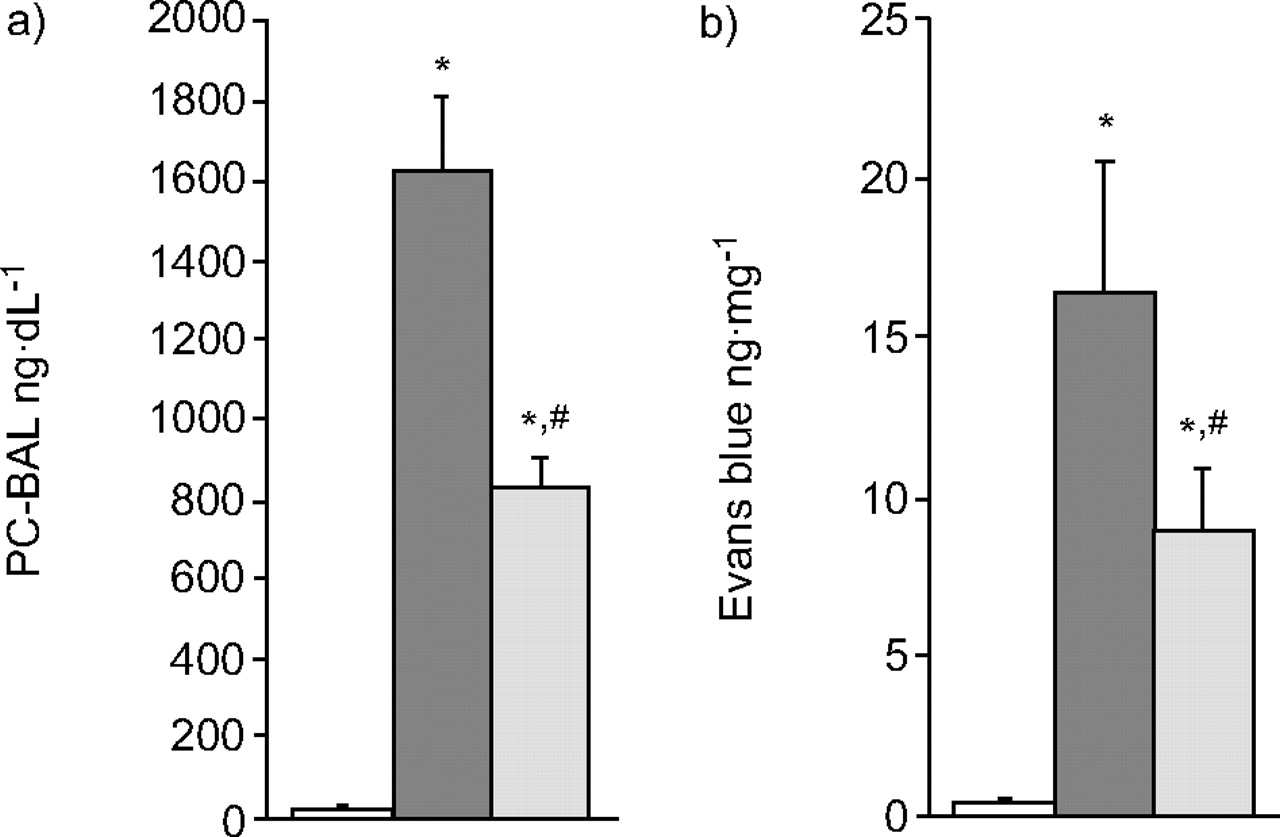
I/R caused increases in protein and tracer dye leakage in the isolated lungs. Post-treatment with NCA attenuated the PC-BAL and tracer dye content (fig. 2⇓).
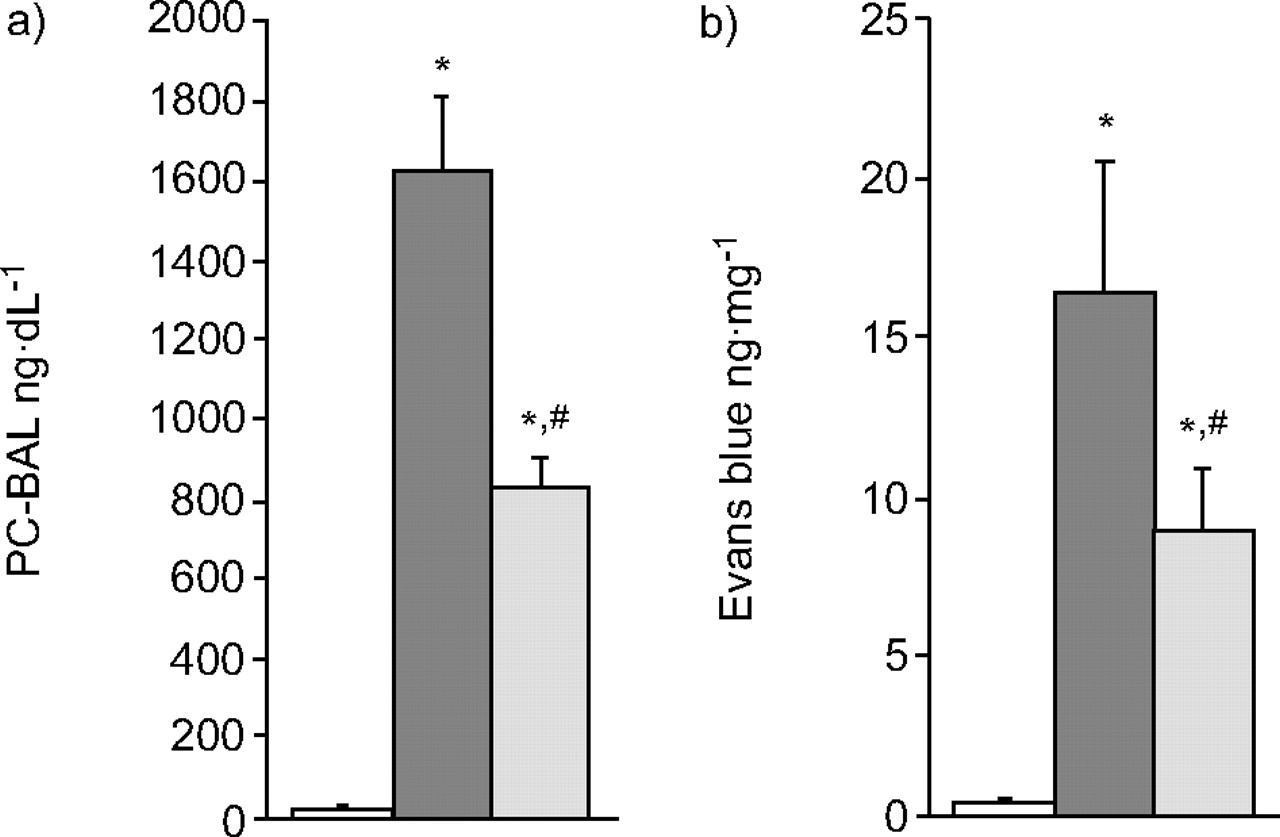
Protein concentration in a) bronchoalveolar lavage (PC-BAL) and b) Evans blue content in lung tissue. I/R increased the PC-BAL and tracer dye leakage. Post-treatment with nicotinamide (NCA) abrogated the ischaemia/reperfusion (I/R)-induced protein and dye permeation. □: vehicle (n = 10); ▒: I/R alone (n = 10); ░: I/R with NCA (n = 10). *: p<0.05 versus vehicle; #: p<0.05 versus I/R.
ATP content and PARP activity in lung tissue
I/R resulted in a decrease in ATP content and an increase in PARP activity. NCA post-treatment reduced the I/R-induced elevation of PARP, while restoring the ATP content (fig. 3⇓).

a) Adenosine triphosphate (ATP) content and b) poly (adenosine diphosphate-ribose) polymerase (PARP) activity in lung tissue. Ischaemia/reperfusion (I/R) decreased the ATP content, while it increased the PARP activity. Post-treatment with nicotinamide (NCA) reduced the PARP activity, while restoring the ATP content. □: vehicle (n = 10); ▒: I/R alone (n = 10); ░: I/R with NCA (n = 10). *: p<0.05 versus vehicle; #: p<0.05 versus I/R.
Nitrate/nitrite, MG, TNF-α and IL-1β in lung perfusate
The NO metabolites, hydroxyl radical and pro-inflammatory cytokines in lung perfusate were increased remarkably following I/R. NCA post-treatment abrogated the I/R-induced changes (fig. 4⇓).

a) Nitrate/nitrite, b) methyl guanidine (MG), c) tumour necrosis factor (TNF)-α, and d) interleukin (IL)-1β in lung perfusate. Ischaemia/reperfusion (I/R) significantly elevated these factors. Post-treatment with nicotinamide (NCA) attenuated the effects of I/R on the nitric oxide metabolites, hydroxyl radical and pro-inflammatory cytokines. □: vehicle (n = 10); ▒: I/R alone (n = 10); ░: I/R and NCA (n = 10). *: p<0.05 versus vehicle group; #: p<0.05 versus I/R group.
Expression of iNOS and eNOS mRNA in lung tissue
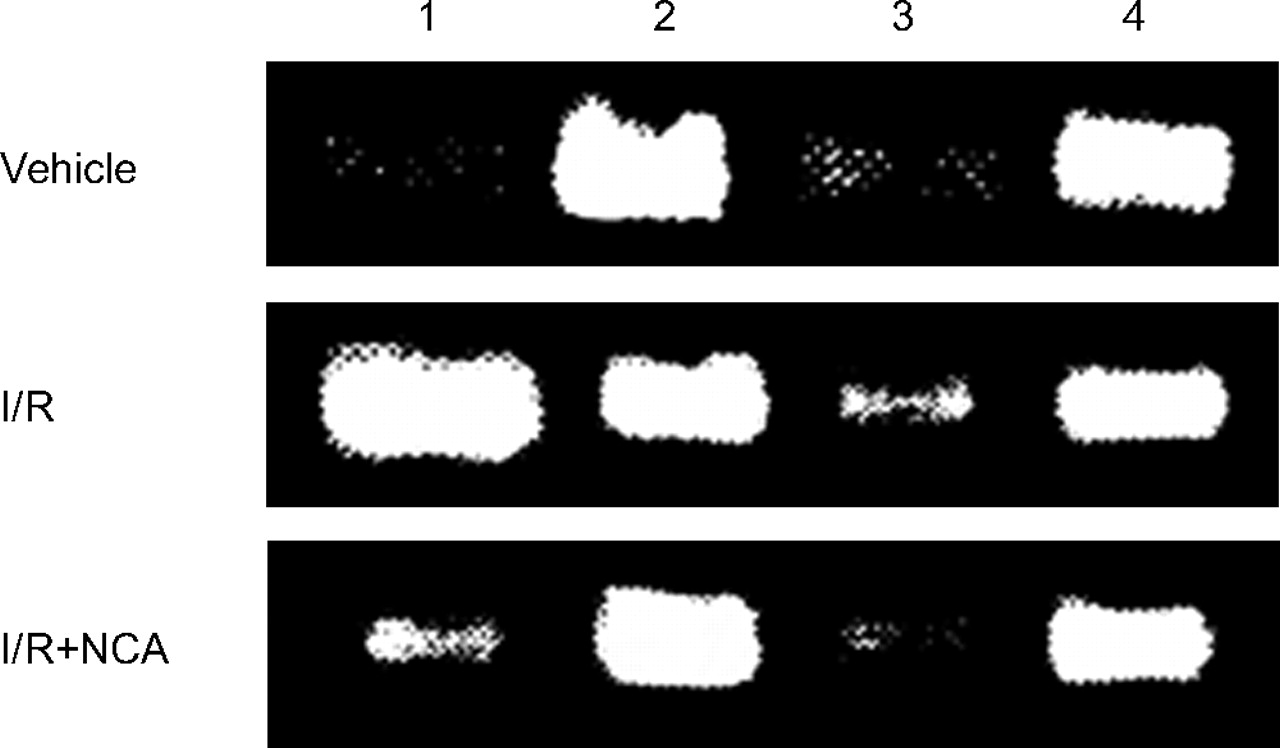
Figure 5⇓ shows the expression of iNOS and eNOS mRNA in lung tissue following vehicle, I/R alone and I/R with NCA. Table 1⇓ summarises the values of the iNOS/GAPDH and eNOS/GAPDH ratios. The data indicated that I/R in isolated lungs caused a great increase in iNOS mRNA expression and a slight increase in eNOS mRNA expression. These effects were diminished by post-treatment with NCA.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of inducible and endothelial nitric oxide synthase (iNOS and eNOS, respectively) mRNA with RT-PCR in the lung tissues following various treatments. The expression of reduced glyceraldehyde phosphate dehydrogenase (GAPDH) was used as an internal standard. Lanes 1 and 3 were iNOS and eNOS, respectively. Lanes 2 and 4 were GAPDH. I/R: ischaemia/reperfusion; NCA: nicotinamide.
Inducible and endothelial nitric oxide synthases(iNOS and eNOS, respectively) mRNA expression in lung tissue
DISCUSSION
In the present study, I/R was employed to induce acute lung injury in isolated perfused rats’ lungs. In a previous study 4, static lung inflation was able to reduce I/R lung injury. In the present study, the isolated lung was kept in an inspired state. The differences in the experimental design were that the length of ischaemia was 95 min and the endotracheal pressure was 2 cmH2O compared with 75-min ischaemia and an endotracheal pressure of 5 cmH2O in the study by Kao et al. 4. The prolongation of an ischaemia period and reduction in endotracheal pressure caused a different extent of I/R lung injury.
The current authors found that NCA, a PARP inhibitor, exerted protective effects on the ALI following I/R. This agent reduced the changes in LW, microvascular permeability, protein and dye leakage. In addition, NCA resulted in the reduction of NO metabolites, hydroxyl radical and pro-inflammatory cytokines in the lung perfusate after I/R. I/R enhanced PARP activity and NOS mRNA expression (iNOS in particular) and decreased ATP content. NCA, the vitamin B-related agent, reduced the PARP activity and iNOS mRNA expression and restored ATP. A previous study revealed the beneficial actions of NCA on the I/R hepatic injury 11. The present study further demonstrated the protective role of NCA on ALI induced by I/R.
Pharmacological inhibition and gene knockout mutants of PARS or PARP have become a new approach for the experimental therapy of various disorders, such as endotoxin shock, stroke, I/R injury, lung inflammation and others 6, 7, 17, 20–23. Activation of PARS or PARP produces cytotoxicity and subsequent cell death and organ dysfunction 24. The results of the present study demonstrated that post-treatment with NCA abrogated ALI induced by I/R in isolated lungs.
A review article addressed the fact that various pharmacological stimulations are able to facilitate alveolar epithelial fluid transport and thereby reduce the ALI caused by different challenges 25. Whether NCA affects the alveolar epithelial water and ion transport system requires further investigation.
In the lungs, PARS or PARP play a key role in the microvascular platelet–endothelial cell interaction induced by: endotoxin; acute lung inflammation following intratracheal administration of endotoxin; induction of asthma; and leukocyte recruitment in systemic endotoxaemia 8, 21–23. These cellular interactions, tissue inflammatory changes and associated changes in adhesion molecules are likely to be the fundamental basis for the pathogenesis of lung injury. In the present study, it was revealed that I/R increased the PARP activity, while reducing the ATP content. NCA attenuated the I/R-induced increase in PARP activity and restored the ATP content. The protective effect of NCA may be mediated, at least in part, through the inhibition of PARP and restoration of ATP.
In addition to the role of PARS or PARP in ALI due to various causes, iNOS may also be a crucial factor in lung damage. In endotoxin-induced lung damage, many studies have provided evidence to suggest that NO production through iNOS is harmful to the lungs in various species and different causes 26, 27. In patients with lung inflammation or injury, the iNOS expression and nitrotyrosine activity were increased 28. PARP was involved in nuclear factor (NF)-κB expression and thereby activated NOS. ADP ribosylation is required to activate NF-κB-mediated iNOS gene transcription 29. Remick et al. 30 also suggested that ADP ribosylation was crucial in the signalling pathway which leads to NOS mRNA expression. PARP inhibitors prevent NOS induction, iNOS mRNA expression and TNF-α release in interferon- and lipopolysaccharide-stimulated macrophages. After inhibition of PARP, iNOS expression, iNOS activity and NO production were reduced 8. PARP inhibition also reduced the production of peroxynitrite and prevented the presence of nitrotyrosine in the tissue 19. It is likely that iNOS is involved in the pathogenesis of tissue injury following endotoxaemia, I/R and other challenges in various organs. In the present study, RT-PCR was used to demonstrate that I/R upregulated iNOS expression. The upregulation was attenuated by post-treatment with NCA.
In the present study, Ppa was found to be elevated by I/R and post-treatment with NCA augmented the I/R-induced pulmonary hypertension. This finding may imply that NCA acts as an inhibitor of iNOS, and the reduction in NO release may account for the effect of NCA on the I/R-induced pulmonary hypertension. In this regard, the pulmonary vasodilatory effect of NO may be the only benefit in I/R lung injury. It is noteworthy that the increase in microvascular permeability was not associated with the changes in Ppa.
In conclusion, nicotinamide protects the lungs from injury by ischaemia/reperfusion. The mechanisms are possibly mediated through the inhibitory effects of this agent on the poly (adenosine diphosphate-ribose) polymerase activity and inducible nitric oxide synthase expression. Subsequently, the production of nitric oxide, free radical and pro-inflammatory cytokines was suppressed, while the adenosine triphosphate content was restored. Since nicotinamide is a compound of vitamin B complex, the protective effects of nicotinamide on the ischaemia/reperfusion lung injury may be a potential therapeutic or preventive regimen for clinical application in lung transplantation or other conditions of lung ischaemia and reperfusion.
Statement of interest
This study was supported in part by grants from the National Science Council, Taipei, Taiwan (95-2320-B-320-004 and 95-2320-B-320-008).
Acknowledgments
The authors would like to thank N. T. Lin (Dept of Microbiology, Tzu Chi University, Hualien, Taiwan) for her help in the detection of nitric oxide synthase mRNA using RT-PCR.
- Received March 2, 2007.
- Accepted April 30, 2007.
- © ERS Journals Ltd
References










