





Abstract
The pathological changes in idiopathic pulmonary fibrosis (IPF) typically start in subpleural lung regions, a feature that is currently not explained. IPF, as well as bleomycin-induced lung fibrosis, are more common in smokers. We hypothesised that carbon particles, which are major components of cigarette smoke that are transported to alveoli and pleural surface, might be involved in the development of subpleural fibrosis through interaction with pleural mesothelial cells.
Carbon particles were administered to mice in combination with bleomycin through intratracheal and/or intrapleural injection and fibrosis was assessed using histomorphometry.
Carbon administered to the chest cavity caused severe pleural fibrosis in the presence of bleomycin, whereas bleomycin alone had no fibrogenic effect. The pleural response was associated with progressive fibrosis in subpleural regions, similar to IPF in humans. Matrix accumulation within this area evolved through mesothelial-fibroblastoid transformation, where mesothelial cells acquire myofibroblast characteristics. In contrast, carbon did not exaggerate bleomycin-induced pulmonary fibrosis after combined intratracheal administration.
This represents a novel approach to induce a robust experimental model of pleural fibrosis. It also suggests that carbon particles might be involved as a cofactor in the initiation and/or progression of (subpleural) pulmonary and pleural fibrosis. Mesothelial cells appear to be critical contributors to this fibrotic process.
- Drug-induced pneumonitis
- epithelial-to-mesenchymal transition
- idiopathic pulmonary fibrosis
- myofibroblast
- pleura
- transforming growth factor-β
Fibrosis is a common denominator of chronic disorders, including lung diseases such as asthma, pulmonary fibrosis, pulmonary hypertension and pleural fibrosis. Pleural and pulmonary fibrosis have many features in common. The most obvious features are that both disorders are characterised by excessive deposition of extracellular matrix and result in restrictive impairment of lung function. Pleural fibrosis as a disease entity is not as well recognised as pulmonary fibrosis but, nevertheless, it can cause severe functional impairment and markedly reduce quality of life. The initiating events for pleural fibrosis are often identifiable and include drugs, radiation, asbestos, haemothorax, thoracotomy for coronary bypass and, most often, infectious diseases 1. This is in strong contrast to idiopathic pulmonary fibrosis (IPF), which has no underlying cause. However, there are cases in which even pleural fibrosis is idiopathic, these cases are typically progressive and lead to death 2–4. Similar to IPF, there is no effective therapy to reverse established pleural fibrosis. In contrast to its negative impact as a disease, pleural fibrosis could become a desirable therapeutic approach in order to treat the accumulation of refractory pleural effusion. Thus, research of mechanisms of pleural fibrosis and of effective pleurodesing agents is highly relevant and has excitingly divergent angles 5, 6.
IPF usually progresses quite rapidly over several years and always compromises pulmonary function. The disease is characterised by myofibroblast proliferation and accumulation of fibroblastic foci, which along with progressive collagen deposition destroy the lung architecture. Recent evidence suggests that IPF is the result of abnormal healing responses to repeated microscopic injury of the lung. Interestingly, one of the hallmarks of IPF is that not only early abnormalities but also advanced fibrosis is predominantly located in the subpleural regions 7. This characteristic feature and as yet unexplained feature of IPF was one of the rationales for the experiments described in our study, as well as the investigation of pleural fibrosis itself. We recently reported that over expression of the fibrogenic cytokine transforming growth factor (TGF)-β1 in the pleural space not only induces progressive pleural fibrosis, but is also associated with abnormal collagen deposition within the subpleural lung parenchyma. We also demonstrated that mesothelial cells play a critical role in pleural fibrosis and we proposed that they may be involved in the subpleural initiation of IPF 8.
Bleomycin is well known to induce a toxic response leading to lung fibrosis in humans 9, 10. Bleomycin is also an efficient and widely used compound for the induction of experimental pulmonary fibrosis in animals. In contrast, it is rather difficult to generate pleural fibrosis in small rodents using bleomycin, although it is a valid option for therapeutic pleurodesis in humans 11. Cigarette smoke is a risk factor for the development of IPF 12 and bleomycin-induced lung fibrosis in patients 13, and it can exaggerate experimental fibrosis in rodents 14. It is unclear if a specific compound in cigarettes enhances the fibrogenic process, or if this is due to the additional alveolar injury. Tobacco smoke is a complex mixture of >4,000 chemicals and, like other by-products of combustion, contains considerable numbers of ultrafine particles 15. Carbon particles are found in cigarette smoke and in pollution, while carbon black is also widely used in the industry. Once inhaled, these particles can reach the alveolar lumen and move towards the pleural surface 16. It is very likely that the microscopic alveolar injury proposed to exist in IPF is caused by several noxious insults, and we hypothesised that cigarette-smoke components, specifically carbon particles, might be involved as one of the cofactors in the initiation and/or progression of (subpleural) pulmonary and pleural fibrosis.
We examined the effect of carbon on bleomycin-induced fibrotic disease of the pleura and lung. We found that carbon particles did not worsen bleomycin-induced fibrosis in lung tissue, but were critical for the induction of pleural and subpleural fibrosis after intrapleural administration of bleomycin. We provide evidence suggesting that mesothelial cells play an important role in this process.
METHODS
Animal procedures
Female Swiss CD1 mice (Charles River, Saint Germain-sur-l'Arbresle, France) were housed in pathogen-free conditions. Rodent food and water were provided ad libitum. The animals were treated according to the guidelines of the Ministère de la Recherche et de la Technologie (Paris, France). All experiments were approved by the Comité d'Ethique de l'Université de Bourgogne (Bourgogne, France). Procedures were performed with inhalation anaesthesia with isoflurane (TEM, Lormont, France). For intratracheal instillation, animals received 50 μL NaCl 0.9%, carbon 0.1 mg·mouse−1 (carbon black 101, 90 nm diameter; Degussa, Frankfurt, Germany), bleomycin 0.07 U·mouse−1 (blenoxane; Bristol-Myers-Squibb, Montreal, QC, Canada) or bleomycin+carbon via a 22G canula introduced within the trachea without surgery. Bleomycin and carbon were diluted in NaCl 0.9%. For intrapleural administration, animals received an intrapleural injection (100 μL) of NaCl 0.9%, carbon (0.1 mg·mouse−1), bleomycin (0.07 U·mouse−1 or 0.48 U·mouse−1) or bleomycin+carbon, as previously described 8.
Pleural lavage fluid (PLF), bronchoalveolar lavage fluid (BALF) and lungs were harvested as previously described 8. Briefly, after abdominal aortic bleeding and slight incision through the diaphragm, PLF was performed with 1.2 mL of NaCl 0.9% injected into the pleural space. A canula was placed into the trachea, lungs were removed and bronchoalveolar lavage was performed with 1 mL of NaCl 0.9%. Lungs were inflated with and placed in formalin for 24 h. PLF and BALF were centrifuged and supernatants were stored at -80°C. Cytospins of pellets were counted (Giemsa stain).
Pleurodesis score
The macroscopic pleurodesis was graded as follows 17: 0: normal pleura; 1: less than three adhesions; 2: more than three adhesions, but localised; 3: generalised scattered adhesions; 4: complete obliteration of pleural space.
TGF-β1 levels
Total human TGF-β1 was determined from PLF or culture supernatant using ELISA (R&D Systems, Lille, France), performed according to the recommendations of the manufacturer. The sensitivity of this assay was 7 pg·mL−1.
Histology
Transverse sections were paraffin embedded, 5-μm sectioned, and stained with Masson-Trichrome and Picrosirius Red. Heat shock protein (HSP)47 immunohistochemistry and dual staining immunofluorescence with antibodies to α-smooth muscle actin (α-SMA) and cytokeratin were performed as previously described 8.
The pleura thickness was determined by histomorphometric measurement on sections stained with Masson-Trichrome as previously described 8: 20 random measures per section per animal were obtained (Eclipse E600 microscope; Nikon, Champigny-sur-Marne, France). Images (colour camera 3 CCD; Sony, Nikon, Champigny-sur-Marne) were analysed using an analysing system (Archimed Instruments, Evry, France).
Collagen was analysed on sections stained with Picrosirius Red as previously described 8: 20 random fields per animal were digitised under polarised light. The percentage of emission was quantified (Histolab, Microvision Instruments, Evry) as a reflection of collagen content. Collagen intensity in the pleura was measured within a rectangle (constant length of 100 μm, width depending on pleura thickness). Collagen content was expressed as percentage of emission multiplied by the surface of each rectangle. Collagen within the subpleural zone was measured using circles (diameter 65 μm) randomly placed at a constant distance from the pleural surface. Collagen content within circles was expressed as percentage of emission.
Zymography
The gelatinolytic activity of matrix metalloproteinase (MMP)-2 and MMP-9 in PLF or cell culture supernatants was measured by zymography 8. Samples were separated by 10% sodium dodecylsulfate-polyacrylamide gel electrophoresis containing 0.1% gelatin (Sigma, St Quentin Fallavier, France). After electrophoresis, the gels were incubated in 2.5% Triton X-100 (Sigma) for 30 min and were then placed in the activating buffer (50 mM Tris-HCl pH 8, 10 mM CaCl2, 5 μM ZnSO4, 150 mM NaCl; Sigma) overnight at 37°C. The gels were stained with 0.1% Coomassie brilliant blue-250 solution (Sigma), for 30 min at 37°C. The gels were then destained with several changes of 40% methanol and 7% acetic acid. Zones of enzymatic activity were evident as clear bands against a blue background. Reference standards were MMP-2 and MMP-9 (Chemicon International, Paris, France).
Mesothelial cell culture
Primary culture of mesothelial cells was performed from female Sprague Dawley rats (Charles River). Rats were euthanised by abdominal aortic bleeding. After incision through the diaphragm, 10 mL trypsin -0.4% EDTA was injected into the pleural cavity. After 30 min, the fluid was aspirated and put into 15 mL fetal calf serum (FCS) and centrifuged at 340×g for 5 min. The pellet was re-suspended in Dulbecco's Modified Eagle's Medium (DMEM) with 1% l-glutamine, 1% penicillin-streptomycin and 15% FCS. The cells were incubated at 37°C with 5% CO2. To confirm the mesothelial cell phenotype, we performed immunofluorescence staining for calretinin (Santa Cruz, TebuBio, Le Peray en Yvelines, France). Briefly, cultured mesothelial cells fixed by 4% paraformaldehyde and made permeable by Triton X-100 (Sigma), were incubated with calretinin (1:100) overnight and then with secondary antibody (1:2000, biotinylated goat anti-mouse/rabbit immunoglobulin (Ig); Dakocytomation, Trappes, France).
4/4RM4 mesothelial cells were incubated in Ham's F10 with 1% l-glutamine, 1% penicillin-streptomycin and 15% FCS. The cells were incubated at 37°C with 5% CO2.
Mesothelial cells (primary and 4/4RM4) were cultured, without FCS, with carbon (250 μg·mL−1), bleomycin (25 mU·mL−1) or bleomycin+carbon for 48 h. Carbon was suspended in medium by ultrasound.
Immunofluorescence
Staining with antibodies to E-cadherin (BD System, Le Pont-de-Claix, France) and α-SMA (DakoCytomation) was performed using fluorescently labelled antibodies. Briefly, 4/4RM4 mesothelial cells (ECACC, Fisher rat visceral pleura mesothelium) were fixed by 4% para-formaldehyde and made permeable by Triton X-100 (Sigma), incubated with primary antibody (1:100) overnight and then with secondary antibody (1:2000, biotinylated goat anti-mouse/rabbit Ig; DakoCytomation).
Statistical analysis
Comparisons between groups were performed by the Mann–Whitney test and comparisons between animals in the same group by the Wilcoxon test.
RESULTS
Intratracheal administration of carbon black does not worsen bleomycin-induced pulmonary fibrosis
Intratracheal NaCl or carbon did not induce inflammatory or fibrotic responses by day 21. As expected, intratracheal bleomycin caused pulmonary fibrosis by day 21 with persistent inflammation (fig. 1⇓). Carbon co-administration did not alter bleomycin-induced inflammation or fibrosis. Collagen was significantly higher in both bleomycin groups, compared with saline and carbon, and was identical in bleomycin and bleomycin+carbon groups (fig. 1a–d⇓). Similar inflammation was observed in bleomycin and bleomycin+carbon groups (fig. 1a–d⇓). Total cell count was similarly increased in BALF from bleomycin and bleomycin+carbon groups compared with NaCl and carbon (p<0.01; data not shown). There was no difference between groups in differential cell count.

Histology (Masson Trichrome) of mouse lungs 21 days after intratracheal instillation with a) NaCl, b) carbon black (CB; 0.1 mg·mouse−1), c) bleomycin (BL; 0.07 U·mouse−1) or d) BL+CB. Scale bars = 100 μm. e) Histomorphometric quantification of collagen intensity on lung sections 21 days after intratracheal instillation with NaCl, CB, BL or BL+CB (100 random visual fields per mouse). f) Total transforming growth factor (TGF)-β1 in bronchoalveolar lavage fluid 21 days after intratracheal instillation with NaCl, CB, BL or BL+CB. n = 8 mice for NaCl and CB groups and n = 12 mice for BL and BL+CB groups. #: p<0.0001 versus NaCl and CB, no difference between BL and BL+CB; ¶: p<0.03 versus NaCL and CB.
Total TGF-β1 (fig. 1e⇑) was significantly increased in BALF from bleomycin and bleomycin+carbon animals (96±41 and 109±55 pg·mL−1, respectively). Total TGF-β1 was not detectable in BALF from animals treated with NaCl or carbon.
Intrapleural co-administration of bleomycin and carbon induces severe and progressive pleural and subpleural fibrosis
Mice receiving NaCl, carbon or bleomycin intrapleurally did not develop pleural fibrosis. There were some carbon spots on the pleural surface in the carbon group by day 7, which disappeared by day 21 (fig. 2a–d⇓). Microscopically, the pleura was a thin monolayer without collagen accumulation in NaCl and carbon groups. In the bleomycin group, there was a slight inflammatory response by day 7 (fig. 2e–l⇓) that had resolved by day 21. The total cell number was similarly increased in PLF from the carbon, bleomycin and bleomycin+carbon groups compared with NaCl. Neutrophils were increased in the bleomycin and bleomycin+carbon groups (table 1⇓). Mice receiving bleomycin+carbon developed severe and progressive pleural fibrosis (fig. 2a–d⇓). Pleural inflammation was apparent by day 7. By day 21, it was progressively increased and fibrosis was a major feature (fig. 2e–l⇓). There were abundant adhesions which complicated lung extraction; retrieval PLF was impossible. Histomorphometry showed increasing pleural thickness and collagen accumulation up to day 21 (fig. 2m⇓). By day 21, the pleurodesis score (table 2⇓) after intrapleural injection of bleomycin+carbon was significantly higher (3.8±0.3) compared with the other groups (NaCl 0±0, carbon 0.3±0.3 and bleomycin: 0.4±0.4).

Intrapleural co-administration of bleomycin (BL) and carbon black (CB) induced severe and progressive pleural and subpleural fibrosis. a–d) Macroscopic view of the pleura 21 days after intrapleural injection of a) NaCl, b) CB (0.1 mg·mouse−1), c) BL (0.48 U·mouse−1) or d) BL+CB. e–l) Histology of mouse pleura and subpleural lung parenchyma e–h) 7 days and i–l) 21 days after intrapleural injection (Masson Trichrome) with e, i) NaCl, f, j) CB, g, k) BL and h, l) BL+CB. Scale bars = 50 μm. m) Histomorphometric quantification of pleural thickness (○) and collagen intensity (bars) on lung sections 2, 7 and 21 days after intrapleural (i.p.) injection with NaCl (░), CB, (□) BL (▪) or BL+CB (▓). 20 random visual fields per mouse. #: p<0.0001 compared with all other groups. n) Histomorphometric quantification of collagen on lung sections in subpleural parenchyma using two diameter-constant circles disposed below the pleura 2, 7 and 21 days after intrapleural injection. ▓; circle 1; ▪: circle 2. 20 random visual fields per mouse. n = 8 mice for the NaCl and CB groups and n = 12 mice for the BL and BL+CB groups. #: p<0.0001 versus NaCl and CB. o, p) Collagen within the subpleural zone was measured using two diameter-constant circles (diameter 65 μm). Circle 1: white; circle 2: green.
Total and differential cell count in bronchoalveolar lavage fluid(BALF) and pleural lavage fluid (PLF) 7 days after intrapleural injection
Pleurodesis scores 7 and 21 days after intrapleural injection
Interestingly, the fibrotic changes in bleomycin+carbon-treated animals were not restricted to the pleura, but extended into the lung parenchyma adjacent to the pleural surface (fig. 2n⇑). Collagen was quantified within this subpleural area by morphometry and progressive accumulation deep in the lung parenchyma was found (fig. 2n⇑). Collagen was most prominent underneath the mesothelial cell layer (circle 1) and decreased with increasing distance from the surface (circle 2).
To our surprise, intraperitoneal or intranasal bleomycin concomitant with intrapleural carbon induced a strong fibrotic pleural response by day 21 (fig. 3⇓), similar to combined intrapleural bleomycin+carbon administration.
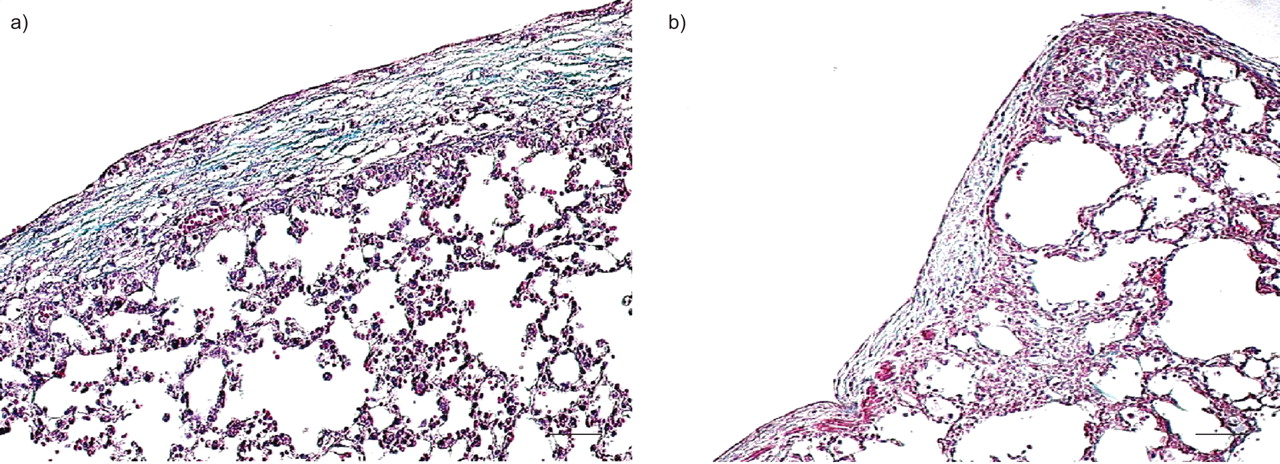
Systemic a) intraperitoneal (0.48 U·mouse−1) or b) intranasal (0.08 U·mouse−1) bleomycin combined with intrapleural carbon black induced pleural fibrosis by day 21 (Masson Trichrome). n = 5 mice for each group. Scale bars = 100 μm.
In contrast to extracellular collagen, the collagen chaperon HSP47 18 was induced in mesothelial cells in carbon, bleomycin and bleomycin+carbon by day 7, compared with NaCl. HSP47 was seen in the subpleural area only in bleomycin+carbon-treated mice along with collagen (fig 4⇓).

Immunohistochemistry showing heat shock protein (HSP)-47 (brown stain) was slightly over expressed in mesothelial cells 7 days after b) carbon black (CB) or c) bleomycin (BL) intrapleural injection compared with a) NaCl-treated animals. d) HSP-47 was strongly over expressed within the pleura and the subpleural area in the BL+CB group. n = 8 mice for NaCl and CB groups and n = 12 mice for the BL and BL+CB groups. Scale bar = 20 μm.
The tissue was analysed for evidence of mesothelial-fibroblastoid transformation, a process involving mesothelial cells and similar to epithelial-to-mesenchymal transition (EMT) 8. Zymography on PLF at day 7 showed increased MMP-9 activity in the bleomycin+carbon group (fig. 5a⇓). The mesothelial layer from bleomycin+carbon-treated animals on day 7 contained a substantial number of cells positive for the epithelial marker cytokeratin and the myofibroblast marker α-SMA (dual staining immunofluorescence). By day 21, these dual-labelled cells persisted and numerous α-SMA-positive cells were present in the thickened pleura (fig. 5b–i⇓), strongly suggesting transition of mesothelial cells to a myofibroblast phenotype. The bleomycin-treated mice had a smaller amount of dual-labelled cells within the pleura by day 7, which vanished by day 21 without evidence for increased persistence of α-SMA-positive cells. The NaCl or carbon groups were normal at any time-point. TGF-β1 is known to induce EMT and was significantly increased in PLF at day 7 in bleomycin and bleomycin+carbon groups (fig. 5j⇓).

Mesothelial cells show features of mesothelial-fibroblastoid transformation, a process similar to epithelial-to-mesenchymal transition. a) Gelatin zymography from pleural lavage fluid (PLF) 7 days after intrapleural injection with NaCl, carbon black (CB), bleomycin (BL) or BL+CB. Matrix metalloproteinase (MMP)-9 activity was only increased in BL+CB-treated mice. Std: standard. b-i) Immunofluorescence of mouse pleura b–e) 7 days and f–i) 21 days after intrapleural injection with b, f) NaCl, c, g) CB, d, h) BL and e, i) BL+CB (cytokeratin stained green, α-smooth muscle actin stained red and DAPI stained blue). Immunofluorescence showed few dual-positive cells in BL mice and frequent dual-labelled cells in BL+CB-animals (solid arrows) 7 days after intrapleural injection. These cells were persistent by day 21 and were only associated with α-smooth muscle actin-positive cells (dotted arrow) within the thickened pleura animals in BL+CB mice. Scale bars = 20 μm. j) Total transforming growth factor (TGF)-β1 in PLF 7 days after intrapleural injection with NaCl, CB, BL or BL+CB. n = 8 mice for NaCl and CB groups and n = 12 mice for the BL and BL+CB groups. **: p<0.01 versus NaCl; #: p<0.003 versus NaCl; ¶: p<0.03 versus CB.
Mesothelial cells are active in this process
Primary rat mesothelial cells were exposed to carbon suspended in DMEM. After 48 h, agglomerates of carbon were seen inside mesothelial cells, suggesting active internalisation of these particles (fig. 6a⇓ and b). This was also observed in mesothelial cells in the animal model (fig. 6a⇓ and b). Gelatin zymography on culture supernatants of mesothelial cells confirmed increased MMP-9 activity in both carbon groups (fig. 6c⇓), similar to PLF.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mesothelial cell activity in the process of pleural fibrosis. Internalisation of carbon nanoparticles by mesothelial cells a) in vitro (calretinin stained green and DAPI stained blue) and b) in vivo (stained by Masson-Trichrome), 21 days after intrapleural injection with carbon black (CB). Scale bars = 20 μm. c) Gelatin zymography from culture supernatants after mesothelial cell incubation with NaCl, CB (250 μg·mL−1), bleomycin (BL; 25mU·mL−1) or BL+CB for 48 h. Matrix metalloproteinase (MMP)-9 activity was increased after treatment with CB (with or without BL). This experiment was repeated three times. Std: standard. d–k) Immunofluorescence on 4/4RM4 mesothelial cells with d–g) α-smooth muscle actin (SMA) (green staining) and h–k) E-cadherin (green staining, DAPI in blue, 630X) showing an increase in α-SMA and a decrease in E-cadherin after 48 h treatment with d, h) NaCl, e, i) CB, f, j) BL or g, k) BL+CB. Scale bars = 10 μm.
4/4RM4 mesothelial cells exposed to bleomycin or to bleomycin+carbon showed an increase in α-SMA and a decrease in E-cadherin expression after 48 h (fig. 6d–k⇑).
DISCUSSION
The pleura is a biologically highly active structure. The mechanisms of pleural fibrosis are not completely understood, and even more mysterious are the mechanisms of subpleural lung fibrosis, one of the hallmarks of IPF in humans. Our study provides an insight into the pathogenesis of both these fibrotic conditions and highlights a potential and, as yet, unexplored role of the pleural mesothelium. Our study also describes a novel rodent model of pleural fibrosis, which may have considerable impact on future research in the area of pleural fibrosis and pleurodesis.
The subpleural predominance of fibrosis is a characteristic feature of IPF, especially in the earlier disease stages 19. To date, there is no clear explanation for this. Being aware of the relationship between smoking and IPF, we hypothesised that cigarette smoke could be involved not only in susceptibility to pulmonary and pleural fibrosis, but specifically in the development of the subpleural fibrosis pattern in IPF/ usual interstitial pneumonia (UIP), possibly through carbon particles.
There are different models of lung fibrosis, all of which have certain advantages and limitations depending on the underlying questions 20. However, none of these experimental approaches truly models the subpleural fibrosis seen in IPF in humans 21. Intratracheal injection of bleomycin is the most common method for induction of pulmonary fibrosis in animals. It consistently causes inflammation and fibrosis in rodent lungs, although the changes are limited and partially reversible 21. Due to the mode of administration, fibrosis is predominantly peribronchial and central. Systemic bleomycin, e.g. delivered via osmotic mini-pumps or through repeated intraperitoneal injection, causes a more diffuse pattern of fibrosis in the lung, but again without the typical subpleural pathology of UIP 22. In contrast to the parenchymal effect, bleomycin instilled in the pleural space rarely causes pleural fibrosis in rodents, although it can have some pleurodesis effects in humans 23.
Most research related to pleural scarring concerns the induction of pleurodesis as a therapeutic approach to treat chronic effusion associated with metastasised cancer. Rabbit or sheep models are typically employed for this purpose. Rodents, especially mice, are preferred in experimental medicine, largely due to the availability of genetically manipulated strains that allow researchers to dissect mechanistic questions in detail. In the context of experimental pleural fibrosis, there is a dramatic shortfall of rodent models, apart from TGF-β over expression with recombinant protein administration 24 or gene transfer 8. Inflammation can be induced in mouse or rat pleura, but usually resolves over time and chronic scarring and fibrosis rarely develops. The trigger for switching a resolving to a progressively fibrotic tissue response in the pleural cavity has not yet been identified. In this context, our new model should be of considerable interest.
Carbon particles are a component of cigarette smoke and a strong association between smoking and IPF/UIP has been described previously 12. The role of cigarette smoke in pleural fibrosis is not as clear, but some studies have suggested a higher rate of asbestos related pleural fibrosis in smokers while others have not been able to confirm this finding 25. Ultrafine or nanoparticles (<100 nm in size) can enter the body through any exposed surface including lungs, skin and the gastrointestinal tract 26. Carbon black are nanoparticles believed to be inert and harmless to the lungs, although they can induce early and moderate, albeit transient, inflammation 27. There is a large database in the lung particle toxicology literature supporting significant toxic effects of a variety of nanoparticles 28. Amongst all inhaled particles, asbestos fibres are probably best known for having the ability to migrate from the parenchyma to the pleura 29. This particle migration can occur through macrophage uptake or directly without involving cellular vehicles. Holt et al. 16 demonstrated in guinea pigs that macrophages are able to carry carbon dust through the parenchyma towards the pleura. In our study, we administered carbon black together with bleomycin into the trachea or the pleural space of mice. We observed very severe pleural and subpleural fibrotic changes in mice receiving combined intrapleural bleomycin plus carbon, while neither bleomycin nor carbon alone caused fibrosis. As expected, intratracheal bleomycin resulted in inflammation and fibrosis, but this was not exaggerated by the addition of carbon. This is similar to the study by Adamson et al. 30 describing that carbon nanoparticles were retained in rodent lung when administered 4 days after bleomycin, but did not result in an increase of fibrosis. Interestingly, we found that bleomycin does not need to be directly injected into the pleural space: severe pleural fibrosis also developed in mice exposed to intrapleural carbon when bleomycin was administered by the intranasal or intraperitoneal route. Taken together, this suggests that carbon might act as a precipitating or cofactor for bleomycin and, thus, promote the development of pleural fibrosis. Even more interesting is the observation that this “dual hit” experiment resulted in subpleural fibrosis in the lung parenchyma, very similar to early fibrotic changes in IPF/UIP in humans.
The detailed mechanisms for this phenomenon are unclear and need to be thoroughly examined. We have previously shown that TGF-β1 over expression by mesothelial cells can induce subpleural fibrosis together with progressive pleural scarring, suggesting that the mesothelium might be involved in the pathogenesis of pulmonary fibrosis 8. In these experiments, we provided evidence that matrix accumulation and fibrosis within the subpleural area evolved through a process involving mesothelial-fibroblastoid transformation 8. This is a phenomenon where mesothelial cells acquire myofibroblast characteristics, almost identical to EMT 31. Mesothelial cells are versatile elements and can exert multiple biological functions 32. Our study confirms the results from the TGF-β1 over expression experiments and shows phenotypical changes of mesothelial cells during the development of bleomycin plus carbon-induced subpleural fibrosis. We found an increase of dual-labelled cytokeratin/α-SMA positive cells in the mesothelial layer by day 7 and 21. In contrast, mice exposed to intrapleural bleomycin only had these dual-labelled cells until day 7 and their lungs appeared otherwise normal. These results were supported in vitro by demonstrating that mesothelial cells treated with bleomycin acquired myofibroblast characteristics and lost the mesothelial marker E-cadherin. This suggests that bleomycin is able to induce transient mesothelial-fibroblastoid transformation in the pleura, but without a second hit (e.g. by carbon particles in our study) this cellular transformation does not persist and does not lead to progressive fibrosis. A recent report by Cool et al. 33 about the fibroblastic reticulum in human IPF, which suggested that fibroblastic foci are organised in a network fashion rather than being isolated areas of cell accumulation, is in general support of our hypothesis. However, one would ideally demonstrate mesothelial markers in fibroblast foci from IPF patients to confirm this challenging experimental finding.
The increased levels of TGF-β in the PLF of mice exposed to bleomycin plus carbon indicates that endogenous induction of this profibrotic cytokine might be involved in the progressive fibrotic phenotype. MMP-9 activity was enhanced in the pleural space of these animals, suggesting that the presence of active MMPs may be a critical factor for mesothelial-fibroblastoid transformation to be maintained or to progress and result in “invasive” fibrosis in the pleura and adjacent subpleural areas 8. There was no correlation between MMP activity and the inflammatory response. Liu et al. 34 have previously demonstrated that mesothelial cells secrete MMPs in response to phagocytosis of small particles, which would explain why MMP-9 is induced in both carbon black and carbon/bleomycin exposed cells in vitro after 48 h (fig. 6c⇑). In the in vivo system, MMP-9 was only increased in the group that received both carbon black particles and bleomycin (fig. 4d⇑). Our hypothesis states that only the concomitant inflammation (caused by co-administration of bleomycin) induces sustained and mesothelial cell derived MMP-9 activity and, thus, facilitates invasive mesothelial-fibroblastoid transformation.
In summary, our study shows that intrapleural carbon black is a critical cofactor for the development of bleomycin-induced pleural fibrosis in mice. Experimental pleural fibrosis using this approach is extraordinarily severe and is characterised by major collagen accumulation not only within the pleura but also in the adjacent subpleural area. Mesothelial cells appear to be important contributors to this fibrotic process, possibly through induction of MMPs and TGF-β. These cells have to be considered as targets for further treatment development, both for pleural as well as pulmonary fibrosis.
Support statement
The research leading to these results has received funding from the European Community 7th Framework Programme (FP7/2007–2013) under grant agreement number HEALTH-F2-2007-202224 eurIPFnet. N. Decologne is supported by the Société de Pneumologie de Langue Française, the Ligue Bourguignonne contre le cancer, and the Association pour la Recherche sur le Cancer. G. Wettstein is supported by the EU 7th Framework Programme (2007–2013 agreement number HEALTH-F2-2007-202224 eurIPFnet.). M. Kolb has a New Investigator Award from the Canadian Institutes of Health Research. C. Garrido is supported by grants from the Ligue Nationale Contre le Cancer and its committees in the Nièvre and Saône-et-Loire.
Statement of interest
None declared.
Acknowledgments
The authors would like to thank V. San Giorgio (animal quarter technical manager) and all the team from the animal quarter of the University of Burgundy (France) for their invaluable and professional help.
- Received December 1, 2008.
- Accepted June 23, 2009.
- © ERS Journals Ltd
References










