



The mechanism of deep inspiration-induced bronchoprotection: evidence from a mouse model
- Russell S. Wong*,#,
- Alexander N. Larcombe*,#,
- Lynette B. Fernandes¶,
- Graeme R. Zosky*,# and
- Peter B. Noble*,#,+,§,f⇓
- *Division of Clinical Sciences, Telethon Institute for Child Health Research, Subiaco
- #Centre for Child Health Research, University of Western Australia
- ¶Pharmacology and Anaesthesiology Unit, School of Medicine and Pharmacology, University of Western Australia
- +School of Women’s and Infants’ Health, University of Western Australia
- §Centre for Neonatal Research and Education, University of Western Australia
- fPhysiology, School of Biomedical, Biomolecular and Chemical Sciences, University of Western Australia, Crawley, Australia
- P.B. Noble, The University of Western Australia, 35 Stirling Hwy, Crawley, WA 6009, Australia. E-mail: Peter.Noble{at}uwa.edu.au
Abstract
In healthy individuals, deep inspirations (DIs) taken prior to a bronchial challenge reduce the bronchoconstrictor response, which is termed “bronchoprotection”. The mechanism(s) of DI-induced bronchoprotection is unclear.
The forced oscillation technique was used to assess the effect of prior DI on subsequent bronchoconstriction to methacholine (MCh) in BALB/c mice. We assessed likely mechanisms for the bronchoprotective effects of DI including reduced airway narrowing (from changes in airway resistance) and/or closure (changes in tissue elastance) and enhanced bronchodilation to a subsequent DI (% reversal in airway narrowing).
DI prior to MCh challenge: 1) did not reduce but instead enhanced airway narrowing (p<0.05); 2) increased ventilation heterogeneity (p<0.05); 3) enhanced the subsequent bronchodilatory response to DI (p<0.05); and 4) reduced tissue elastance (p<0.05), suggesting opening of closed airways or alveoli units.
Our findings suggest that DI prior to MCh challenge may elicit a series of changes, some of which are beneficial to respiratory function (enhanced bronchodilation), while others place greater load on the system (enhanced bronchoconstriction and ventilation heterogeneity). It is proposed that the relative magnitudes of these opposing physiological and mechanical effects will determine the net effect on respiratory function in health and disease.
- Airway hyperresponsiveness
- asthma
- bronchodilation
- bronchoprotection
- deep inspiration
Airway hyperresponsiveness (AHR) is a primary characteristic of asthma whereby the airways narrow excessively to bronchoconstrictor stimuli [1]. While the mechanism underlying AHR is unclear, there are a number of physiological determinants of airway calibre that may be disrupted by disease processes. Regular airway stretching by deep inspiration (DI) is believed to play a major role in maintaining airway calibre and impairment of this innate protective mechanism is proposed as a precursor to AHR [2–4].
In healthy humans, DI produces bronchodilation in previously contracted airways [5–8], a beneficial effect that maintains airway patency. In addition to these well-established bronchodilatory effects of DI, when bronchial challenge is preceded by DI the evoked bronchoconstrictor response is attenuated [6, 9, 10]. The effect of DI in reducing subsequent bronchoconstriction, dubbed “bronchoprotection” [10], is absent in asthmatic individuals [10, 11], suggesting that impairment in this normal physiological response to DI may be involved in AHR [11].
The mechanism(s) behind DI-induced bronchoprotection is not well understood and is an important area of study if the abnormalities producing AHR are to be revealed. The bronchoprotective effects of DI have typically been assessed from the change in forced expiratory volume in 1 s (FEV1) in response to a bronchial challenge, where DI prior to challenge reduces the fall in FEV1 [6, 9–11]. It is intuitive to then consider the different factors that influence FEV1, which provides insight into the physiology of DI-induced bronchoprotection. First, DI taken prior to challenge may directly reduce airway narrowing. Evidence from isolated airway smooth muscle (ASM) studies in vitro suggest that mechanical stretching of the airway wall during DI reduces subsequent force development [12, 13]. Secondly, DI prior to challenge may prevent airway closure. In a recent study by Chapman et al. [14], bronchoprotective effects of DI were primarily mediated in peripheral airways and it was concluded that the predominant effect was in preventing airway closure. A third and final possibility is that the DI that precedes the maximal expiratory manoeuvre (FEV1) itself produces bronchodilation and the magnitude of this response is enhanced when DIs are taken before a bronchial challenge. Notably, when manoeuvres other than maximal expiration are used to assess the effects of DI prior to challenge; for example, in expiration from a submaximal lung volume (i.e. partial exhalation), bronchoprotection is not observed [7, 15].
The purpose of the present study was to examine the possible mechanism(s) responsible for DI-induced bronchoprotection. Using a mouse model, we assessed whether DI prior to bronchial challenge modified airway narrowing capacity, airway closure and/or the bronchodilatory response to DI. Airway resistance, tissue elastance, tissue damping and ventilation heterogeneity were assessed using the forced oscillation technique (FOT), which allowed us to discriminate between effects mediated centrally and in the lung periphery. Most importantly, unlike spirometry (i.e. FEV1), FOT does not require a DI which allowed us to determine, in a controlled manner, whether DI prior to a bronchial challenge enhances subsequent bronchodilatory responses to DI.
METHODS
Animal handling and preparation
All procedures were approved by the Telethon Institute for Child Health Research (TICHR) Animal Ethics Committee (Subiaco, Australia) and conformed to National Health and Medical Research Council of Australia guidelines. Naïve female BALB/c mice (8 weeks of age, ∼20 g) were housed at TICHR and maintained on a 12/12-h light/dark cycle. Food and water were provided ad libitum. Prior to in vivo measurements of lung mechanics, all mice were anaesthetised by intraperitoneal injection of ketamine (0.4 mg·g−1; Troy Laboratories, Glendenning, Australia) and xylazine (0.02 mg·g−1; Troy Laboratories). Supplemental doses of anaesthetic (50% of the original dose) were administered via i.p. injection every 40 min, as required. Once surgical anaesthesia was induced, a polyethylene cannula (length 10 mm, internal diameter 0.86 mm) was inserted into the trachea and secured with surgical silk. The mouse was connected to an animal ventilator (Flexivent; Scireq, Montreal, Canada) and allowed to stabilise for 5 min prior to commencement of study protocols. Mice were ventilated at a frequency of 450 breaths per minute with a tidal volume of 8 mL·kg−1 and positive end-expiratory pressure (PEEP) of 2 cmH2O. All respiratory measurements described below were also performed under a 2-cmH2O PEEP.
Measurement of lung mechanics by FOT
This study adopted a modification of the FOT [16] to assess changes in lung mechanics in vivo. Briefly, respiratory system impedance (Zrs) was measured during pauses in ventilation by applying an oscillatory signal generated by a loudspeaker via the tracheal cannula. The applied signal comprised nine frequencies: 4, 6, 10, 14, 18, 22, 26, 34 and 38 Hz. A four-parameter model with constant-phase tissue impedance was fit to the Zrs:
where Raw is the Newtonian airway resistance (predominantly airway resistance since contributions from the compliant chest wall are small), j = √-1, ω is the angular frequency, α = (2/π) arctan (G/H), H is the coefficient of tissue elastance and G the coefficient of tissue damping. Inertance (I) is not reported since it is contained almost entirely in the tracheal cannula and is negligible once taken into account. Raw was used to assess airway narrowing and/or bronchodilation. H, which reflects tissue elastance but is sensitive to decreases in lung volume and, hence, increases with airway closure [17], was used to assess potential closure events. The effects of prior DI on ventilation heterogeneities developed after methacholine (MCh) challenge were assessed by hysteresivity, i.e. the ratio of G/H [18, 19]. FOT measurements (6 s) were performed immediately before and after MCh, as described in the Experimental protocols.
In some animals, we also determined whether MCh or DI modified the distensibility of airways. Airway distensibility was measured based on a previous approach [20] where distensibility was related to the slope of lung volume and airway conductance. To determine airway distensibility, airway conductance (1/Raw) was measured during stepwise increases in transrespiratory pressure (the difference in the pressure at the airway opening and the chest wall) (2, 4, 6, 8 and 10 cmH2O). Airway distensibility was determined from the slope of the relationship between airway conductance and transrespiratory pressure.
Simulation of DI
To simulate DI in the anaesthetised mouse, the ventilator expelled a volume of air to inflate/deflate the lung to a specified transrespiratory pressure (fig. 1a). Each DI consisted of an increase in transrespiratory pressure from 2 to 30 cmH2O in 1 s, followed by a 1-s hold and a deflation back to 2 cmH2O in a further 1 s.

a) Schematic indicating the changes in transrespiratory pressure (Prs) during a simulated deep inspiration (DI) which was driven by the mechanical ventilator. b) Schematic of a two-round single-dose methacholine (MCh) protocol, which was used to assess the effects of prior DI. Changes in respiratory mechanics produced by MCh and/or DI were assessed by the forced oscillation technique (FOT). Between challenges, recruitment manoeuvres (RMs) were performed to aid recovery from bronchoconstriction and to minimise ventilator related airway/alveoli collapse. The order of the challenge protocols was randomised.
Experimental protocols
As outlined in figure 1b, each mouse received two single-dose MCh (β-methacholine chloride; Sigma-Aldrich, St Louis, MO, USA) challenges to assess the effects of prior DI (or no DI, i.e. control) on changes in lung mechanics induced by MCh. The aerosolised MCh challenge (30 mg·mL−1 for 15 s) was generated by an ultrasonic nebuliser (UltraNeb; DeVilbiss, Somerset, PA, USA) and delivered into the ventilator circuit.
Once mice were anaesthetised and under mechanical ventilation, a slow inflationary manoeuvre (30 s) to 30 cmH2O (recruitment manoeuvre; RM) was performed to open up lung units that may have closed during surgery. After a further 5 min, a DI was performed to establish a relevant lung volume history. Mice (n=8) subsequently underwent a 20-min period of tidal mechanical ventilation in the absence of DI. At the end of this period, a “prior DI” or “control” challenge was performed in a randomised order. The prior DI challenge comprised five DIs induced immediately before MCh challenge whereas the control challenge did not receive any DI prior to challenge. FOT measurements were performed immediately before and after MCh challenge (30-s intervals for 5.5 min post-MCh) that was either preceded or not by five DIs (fig. 1b). At the end of the first challenge, a further two RMs were performed (10 min apart) to aid recovery from MCh-induced bronchoconstriction and to minimise ventilator-related airway/alveoli collapse. A further 10 min later, the second challenge protocol began, which included one further RM, and other than the presence or not of DI before challenge, the protocols were identical.
In a separate group of mice (n=8), a modified approach was used to determine whether the bronchodilatory response to DI after MCh challenge was influenced by DI taken prior to challenge. Bronchodilatory responses to DI were compared at the same time-point (similar to human lung function studies) [10, 11] instead of the peak of airway narrowing, which differed between groups (see results). Bronchodilation to DI was assessed 1.5 min after the initiation of bronchial challenge. FOT measurements after DI were performed continuously during a brief period of apnoea (∼30 s).
Statistics and analysis
Changes in Raw, H, G and G/H relative to baseline were compared by repeat measures one-way ANOVA and Newman–Keuls post hoc analysis. The effect of prior DI on each parameter before and after MCh was assessed by two-way ANOVA and Newman–Keuls post hoc analysis with prior DI (or otherwise, i.e. control) and time as repeat measure variables. The effects of prior DI on bronchodilation to DI were assessed from % reversal to DI (see later) and were similarly analysed using two-way ANOVA and Newman–Keuls post hoc analysis with prior DI (or control) and time as repeat measure variables.
The magnitude of bronchodilation to DI (post MCh challenge) was assessed from the % reversal in airway narrowing [21, 22], using the formula:
where 100% indicates full reversal of airway narrowing by DI. The rate of airway re-narrowing after DI was assessed by fitting an exponential curve to the % reversal-time data:
where A and B are fitted constants, and computation of rate constant k.
The index of airway distensibility was calculated as the linear least-squares slope of change in 1/Raw over the change in transrespiratory pressure between 2 and 10 cmH2O. Graphical analysis and statistical tests were performed using GraphPad Prism (v4.03; GraphPad Software, San Diego, CA, USA) and Statistica (99 Edn; StatSoft Inc., Tulsa, OK, USA), respectively. All data are presented as mean±sd. p-values of <0.05 were regarded as statistically significant.
RESULTS
Baseline FOT measurements were performed immediately before MCh challenge (i.e. at 0 min) with each mouse receiving two MCh challenges. In preliminary experiments, we established that in the absence of DI, there was no difference in the magnitude of the bronchoconstrictor response produced by the first and the second MCh challenge (n=4; ns).
Effects of prior DI on MCh-induced airway narrowing and closure
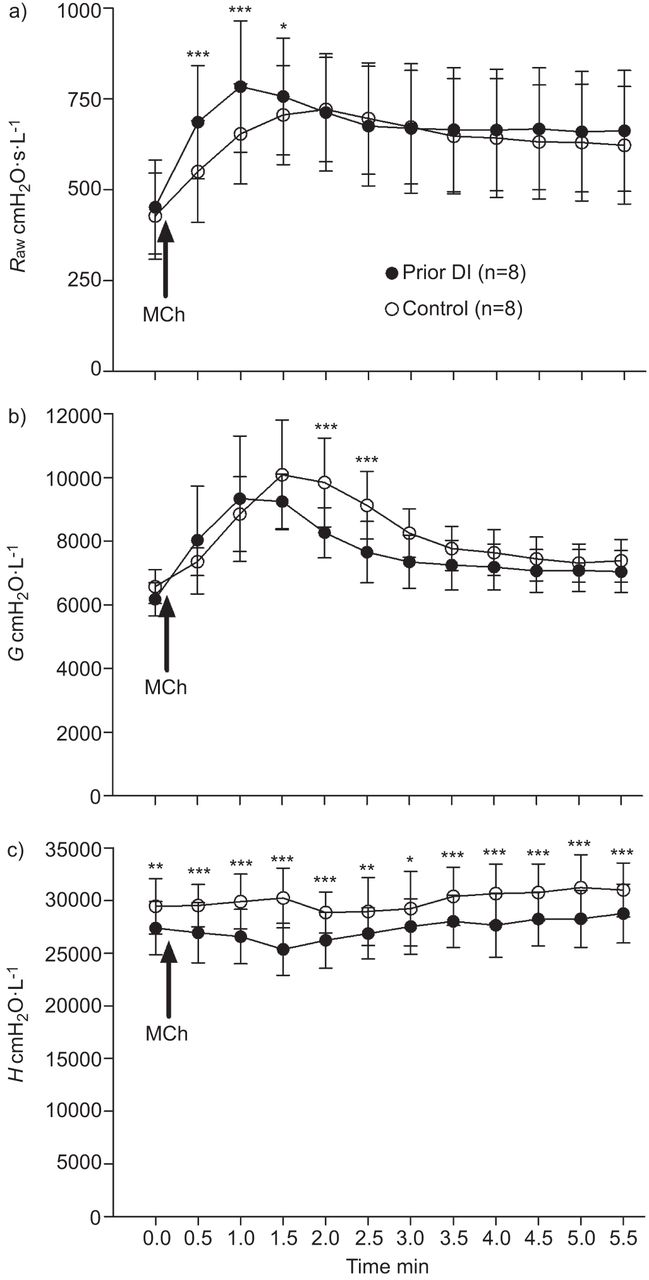
To assess the effects of prior DI on MCh-induced airway narrowing and closure, the full time-course of response was assessed, with measurements taken every 30 s for 5.5 min post-MCh challenge (fig. 2). Before MCh (i.e. baseline), there was no difference in Raw or G between the control and prior DI challenges. In comparison, in the prior DI challenge, H was lower compared with the control challenge (p<0.01).
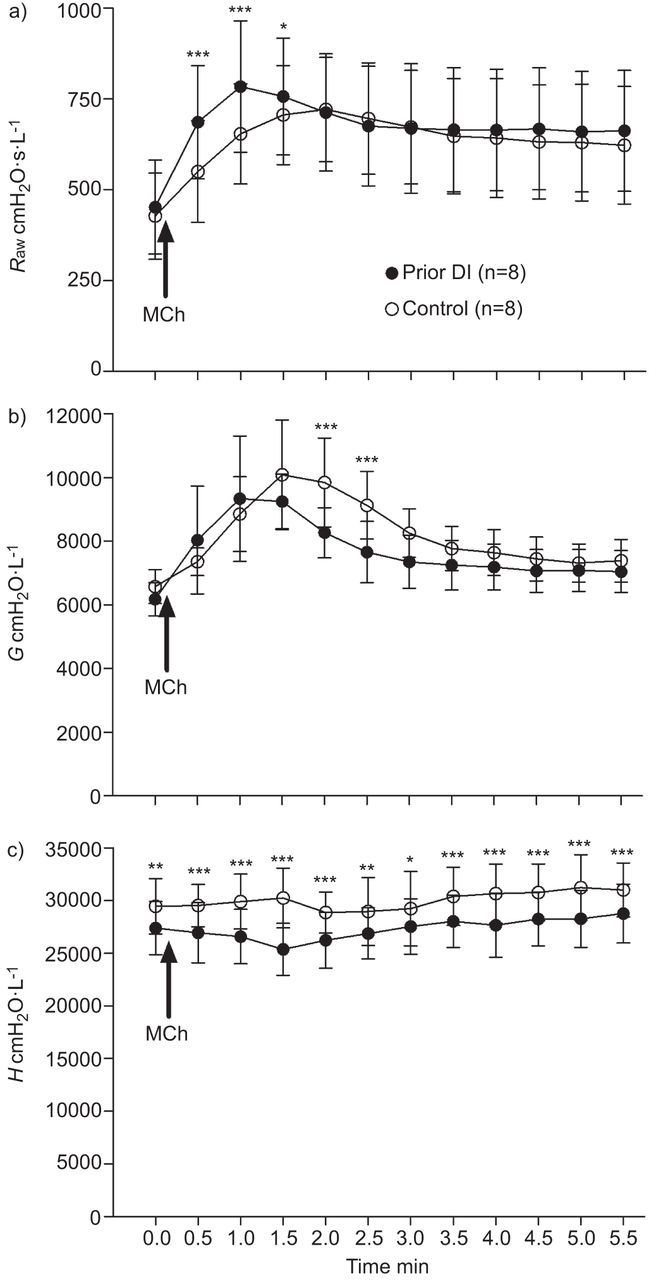
Effects of prior deep inspiration (DI) on methacholine (MCh)-induced changes in airway and lung mechanics in mice. 0 min indicates baseline measurements (i.e. pre-MCh challenge). Immediately after the baseline measurement, a MCh challenge was performed (indicated by arrow), which lasted for 15 s. The first post-challenge measurement was 15 s from the end of the challenge (i.e. 30 s from the beginning of the challenge). Measurements were subsequently taken every 30 s. a) airway resistance (Raw); b) tissue damping (G); c) tissue elastance (H). Data are presented as mean±sd. *: p<0.05 for prior DI versus control challenge protocols; **: p<0.01 for prior DI versus control challenge protocols; ***: p<0.001 for prior DI versus control challenge protocols.
MCh challenge increased Raw (p<0.001) at all time-points compared with baseline in both the prior DI and control challenges (fig. 2a). During the control challenge, G was elevated from 1.0 min (p<0.001) compared with baseline, returning to baseline by 5.5 min (fig. 2b). During the prior DI challenge, G was elevated from 30 s after MCh challenge (p<0.001) and remained elevated at 5.5 min (p<0.05). In general, MCh had little to no effect on H, indicating that airway closure had not occurred (fig. 2c).
The increase in Raw produced by MCh was greater and more rapid in the prior DI challenge (fig. 2a). Raw was elevated in the prior DI challenge compared with the control challenge at 0.5, 1.0 (p<0.001 in both cases) and 1.5 min (p<0.05). There was no difference in Raw between groups after 1.5 min. Maximum narrowing typically occurred at 1.0 min (mode) in the prior DI challenge and at 1.5 min (mode) in the control challenge. Maximum airway narrowing was greater in the prior DI challenge (p<0.05).
G was lower in the prior DI challenge at 2.0 and 2.5 min (p<0.001 in both cases) compared with the control challenge (fig. 2b). There were no other differences in G between the two challenge protocols. The fact that H differed at baseline between challenge protocols (fig. 2c) made it difficult to determine the effects of prior DI on MCh-induced changes. H post-MCh challenge was therefore normalised to baseline and compared by two-way ANOVA. The results of this analysis showed that only at one time-point (1.5 min) was H proportionally lower in the prior DI challenge (p<0.001).
The effects of prior DI on ventilation heterogeneities developed post-MCh were assessed by hysteresivity, i.e. the ratio of G/H (fig. 3). Hysteresivity was elevated after MCh from 1 to 3 min in the control challenge and from 0.5 to 2.5 min in the prior DI challenge (p<0.001 to p<0.01). There was an increase in hysteresivity in the prior DI compared with the control challenge at 0.5 min (p<0.05) and 1.0 min (p<0.01) after MCh. There was no difference in hysteresivity at any other time-point.

Effects of prior deep inspiration (DI) on methacholine (MCh)-induced changes in hysteresivity (G/H) in mice. 0 min indicates baseline measurements (pre-MCh challenge). Data are presented as mean±sd. *: p<0.05 for prior DI versus control challenge protocols; **: p<0.01 for prior DI versus control challenge protocols.
Effects of prior DI on DI-induced bronchodilation
Bronchodilation to DI was assessed 1.5 min after MCh challenge in both protocols (fig. 4a) and tracked continuously for 30 s. Before bronchodilation to DI, Raw was enhanced in the prior DI challenge compared with the control challenge at 0.5, 1.0 and 1.5 min (p<0.001, p<0.001 and p<0.01, respectively). This was consistent with the previous experiment (fig. 2a), demonstrating that this finding was reproducible.

Effects of prior deep inspiration (DI) on subsequent DI-induced bronchodilation. a) airway resistance (Raw) before and after DI (indicated by arrow); b) % reversal in airway narrowing produced by the DI. Data are presented as mean±sd. *: p<0.05 for prior DI versus control challenge protocols; **: p<0.01 for prior DI versus control challenge protocols; ***: p<0.001 for prior DI versus control challenge protocols. MCh: methacholine.
After DI was induced post-MCh, levels of Raw decreased substantially in both challenge protocols, indicating bronchodilation had occurred (fig. 4a). This was followed by rapid airway re-narrowing whereby Raw returned to pre-DI levels by 3 and 13 s after post-MCh DI in the control and prior DI challenge protocols, respectively. Notably, in the control group, Raw was increased above pre-DI levels 14 s after DI (p<0.01), and continued to rise thereafter. Raw after DI was identical between the control and prior DI challenges. The rate of airway re-narrowing post-MCh DI was similar between control and prior DI challenges (mean±sdrate constants were 0.186±0.08 s−1 and 0.184±0.06 s−1 for control and prior DI challenge protocols, respectively; ns).
To determine whether prior DI altered the bronchodilatory response to DI, we computed the % reversal of airway narrowing to DI in each challenge protocol for up to 30 s after the post-MCh DI (fig. 4b). In the prior DI challenge, the % reversal to post-MCh DI was increased and persisted longer compared with the control challenge (p<0.05), which as described above promoted a constrictor response. Immediately after post-MCh DI, there was a 44.5±22% and 61.2±15% reversal in airway narrowing in the control and prior DI challenges, respectively. The absolute change in Raw was also more prominent in challenges preceded by DI (p<0.05). The absolute change in Raw immediately after the post-MCh DI was 165±84 cmH2O·s·mL−1 in the control challenge and 290±177 cmH2O·s·mL−1 in the prior DI challenge.
In view of the results demonstrating an enhanced bronchodilatory response to DI in MCh challenges preceded by DI, in six additional mice, we assessed whether prior DI increased airway distensibility. Figure 5a and b shows the relationship between 1/Raw and transrespiratory pressure under baseline conditions and after MCh challenge for the control and prior DI challenges, respectively. The slope of this relationship was used as an index of airway distensibility. Airway distensibility (fig. 5c) was reduced after MCh challenge (p<0.001). However, there was no difference in airway distensibility between control and prior DI challenges.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relationship between airway conductance and transrespiratory pressure (Prs) (2–10 cmH2O) for a) control and b) prior deep inspiration (DI) challenges. c) Slopes of the linear relationships observed in a) and b) were used to provide an index of airway distensibility. Airway distensibility was reduced after methacholine (MCh) challenge (p<0.001). There was no difference in airway distensibility between challenge protocols. Data are presented as mean±sd.
DISCUSSION
The present study provides a systematic examination of the likely mechanisms mediating the regulatory effect of DI on bronchoconstriction when DI is taken prior to a bronchial challenge. By using a mouse model and FOT, we have shown that prior DI elicits a complicated series of changes, some of which are beneficial to respiratory function (enhanced bronchodilation), while others place greater load on the system (enhanced bronchoconstriction and increased ventilation heterogeneity). We propose that the relative magnitude and interaction between these opposing physiological and mechanical changes determines the net effect on respiratory function in health and disease.
Before discussing major findings, our methodology requires comment. The study design was heavily influenced by the known methodological dependency of responses to DI reported in human subjects. Indeed, the method used to assess changes in airway calibre during bronchoconstriction may influence the apparent effects of DI. Most notably, expiratory flow derived from maximal expiration (i.e. FEV1) is differentially altered compared with flow during expiration from a submaximal expiration (partial expiration) [7, 15]. The FEV1 necessitates a DI, which is significant in that potential bronchoprotective effects become intertwined with the bronchodilatory effects of DI. Experiments were therefore designed to assess the effects of DI prior to bronchial challenge on bronchoconstrictor responses measured before and after DI was induced after MCh challenge. The use of FOT ensured a relatively fixed lung volume during measurement (i.e. no DI required) while at the same time providing information on central and peripheral lung mechanics and ventilation heterogeneity. The FOT signal comprises a broad range of frequencies that allows us to reliably partition Raw from the overall respiratory impedance [16]. This is in contrast to many clinical human FOT studies [14, 23, 24] where the real part of the respiratory impedance, which contains contribution from both airway and tissue resistance, at two to three frequencies is reported. In the present study, changes in Raw were used as an index of bronchoconstriction or bronchodilation. Finally, the inbred murine model limits biological variability, providing greater statistical power and sensitivity to detect small but potentially physiologically meaningful effects of DI. The mouse is an emerging model for the study of DI and, like humans, exhibits a robust response to DI [25, 26], which in the present study was simulated by positive ventilator-driven pressures.
Of all the different factors that may be involved in the bronchoprotective response to DI, the simplest explanation is that DI reduces the narrowing capacity of an airway. There are compelling data from isolated ASM studies in vitro suggesting that mechanical stretch accompanying DI temporarily reduces ASM force generation. In the study by Wang et al. [12], prior length oscillation of isolated ASM strips reduced the subsequent ASM force development, data initially derived using porcine airway tissue [12] and later confirmed using human ASM [13]. However, to the best of our knowledge, no study in humans in vivo has reported a direct effect of DI in reducing airway narrowing, at least using global measures of airflow and Raw [7, 14, 15, 24], although there is some evidence for this in rabbits [27]. The results of the present study argue against a limitation of airway narrowing by prior DI, and instead show a transient increase in Raw to MCh in the prior DI challenge, suggesting that the dominant effect is one of enhanced airway narrowing. In human subjects, the majority of studies showed no effect of prior DI on airway narrowing [7, 14, 24], while one study reported an enhanced bronchoconstrictor response in both healthy and asthmatic subjects [15], in agreement with our present findings. Taken as a whole, these observations fail to provide a convincing case for an association between reduced airway narrowing and bronchoprotective effects of DI.
The reason for an increase in airway narrowing following periods of DI is unknown. Large inflationary pressures have been shown to increase epithelial barrier permeability [28, 29]. An increase in epithelial barrier permeability after DI would favour greater airway narrowing in challenges preceded by DI. Changes in permeability would also explain the observed differences in the rate of airway narrowing, where peak airway narrowing occurred ≥30 s earlier in the prior DI challenge. An alternate explanation for an increase in airway narrowing after DI is that the airway wall exhibits an intrinsic contractile response to mechanical stretch, which may be activated by DI. In naïve airway tissue from both bovine and porcine models [30, 31], simulated DI was shown to enhance contractile responses. Whether a myogenic response to mechanical stretch exists in the uninflamed mouse airway remains to be determined. It is also possible that the mechanical load on the airway wall was reduced by the actions of DI. Reduced airway distending pressure as a result of parenchymal hysteresis has previously been evoked to explain contractile responses to DI in some subjects [32]. However, this may not necessarily apply in our study since Raw was measured at the same transrespiratory pressure, and therefore at the same airway transmural pressure, although local variations in pressure at the peribronchial level are possible. Finally, while H was decreased in the prior DI challenge, there was no change in G, which argues against an alteration in tissue rheological properties and, therefore, the load associated with distortion of tissue attachments. As will be discussed, reduced elastance is more likely explained by the opening of airway passages.
As a direct consequence of increased airway narrowing in challenges preceded by DI, ventilation heterogeneity also increased. Hysteresivity, the ratio of G/H, is an index of ventilation heterogeneity [18, 19] and, in the present study, was elevated in the prior DI challenge. The increase in hysteresivity during the prior DI challenge was apparent early into bronchoconstriction reflecting the greater kinetics of the response. Ventilation heterogeneity adversely affects lung function and in asthma may be involved in the development of AHR [33, 34].
A primary goal of the present study was to determine if DI prior to bronchial challenge enhanced subsequent bronchodilation to DI. This possibility is supported by two independent studies by Crimi and co-workers [7, 15], which showed that the bronchoprotective effects of DI were only apparent when assessed using spirometric manoeuvres involving full lung inflation (i.e. FEV1). Conversely, bronchoprotective effects of DI were not observed when assessed using manoeuvres devoid of a DI (i.e. partial expiration). In the present study, we measured bronchodilation (% reversal) [21, 22] to DI in the presence of MCh-induced bronchoconstriction and determined whether the magnitude of bronchodilation was altered if challenge was preceded by DI. As previously demonstrated in mice [25], there was a potent and transient bronchodilatory response to DI (i.e. dilation of narrowed conducting airways), but importantly, the present results indicate that when a DI is also administered prior to challenge, the subsequent bronchodilatory response is enhanced. Remarkably, while in the prior DI challenge Raw was increased before the initiation of a post-MCh DI (as discussed), after the bronchodilatory effects of DI, Raw was identical between the challenge protocols. Hence, any difference in Raw (narrowing) between the challenge protocols was abolished by the bronchodilatory effects of DI. One other study [35] has previously questioned whether DI prior to challenge enhances subsequent bronchodilation by assessing changes in the maximal to partial flow ratio in human subjects (an increase in this ratio indicates enhanced bronchodilation). While the conclusion drawn was that bronchodilation to DI was unaffected by DI prior to challenge, there was a reasonable trend towards enhanced bronchodilation when DI preceded challenge. From the present data and with supporting evidence from the literature, findings suggest that the bronchodilatory response to DI may be influenced by previous DI and contribute to the apparent bronchoprotective effects of DI.
In an attempt to explain why bronchodilatory responses to DI may be enhanced when preceded by a period of DI, we assessed airway distensibility. Our rationale was that prior DI may increase airway wall compliance and, thus, increase distension of the airway to a subsequent DI. Indeed, the magnitude of airway wall stretch is positively correlated with reduced ASM force, which produces bronchodilation [21, 36, 37]. The assessment of airway wall compliance in vivo is difficult, so we followed a previous approach where airway distensibility was quantified from the slope of the relationship between airway conductance and lung volume [20] or in our study transrespiratory pressure. Airway distensibility was reduced after MCh, which can be explained by contractile-associated stiffening of the airway wall [37, 38] and at least in part due to a geometric effect of a reduced airway calibre. Airway distensibility tended to be greater in the prior DI group before MCh challenge (i.e. relaxed airway), although this was not statistically significant. Moreover, any potential difference between the challenge protocols was no longer apparent after MCh-induced stiffening, which is the more relevant measure as this reflects airway distensibility prior to the induction of DI. Changes in airway distensibility in challenges preceded by DI cannot, therefore, explain the subsequent enhancement in bronchodilatory response to DI.
In this study, we used H as a means of assessing whether MCh produced differential airway closure in challenges preceded by DI. While elastance, by definition, describes the elastic properties of lung tissue, it is sensitive to airway closure and conversely airway opening events [17]. Even before the induction of bronchoconstriction, tissue elastance was lowered in the prior DI challenge and without an accompanying shift in tissue damping (i.e. no change in tissue properties). These data are suggestive of lung recruitment as a result of the opening of closed or near-closed airway passages by DI. Subsequent bronchoconstriction to MCh did not produce airway closure in either challenge protocol. By assessing changes in forced vital capacity (FVC), a previous study determined that the predominant effect of prior DI was in preventing or reducing airway closure and, thus, attenuating the fall in FVC [14]. The authors speculated that the bronchoprotective effects of DI could be attributed to reduced surface tension. Our present results, to an extent, support these findings in that there was reduced airway closure (or alveoli collapse) in the prior DI challenge; however, this was apparent before the induction of bronchoconstriction. It is possible that, even in the absence of induced bronchoconstriction, removal of the beneficial effects of DI may result in airway instability as demonstrated in a recent follow-up study by the same group [23]. Although our results are in agreement with the aforementioned data from spontaneously breathing human subjects [14, 23], in the scenario of mechanical ventilation, alveolar collapse and oedema are common and re-recruitment of these lung regions seems the most likely explanation for the fall in lung elastance after DI. Mechanical ventilation is a necessary intervention in studies using mouse models and this was a limitation of the present study.
The results of the present study suggest that DI prior to a bronchial challenge provokes a complicated series of physiological and mechanical changes. The beneficial effects of DI, which may partly involve enhanced bronchodilation, are offset by opposing and mechanically disadvantageous effects including enhanced airway narrowing and increased ventilation heterogeneity. These findings, to a large degree, mirror contrasting reports in the literature, some of which may be explained by methodological differences, but not all. In consideration of the multiscale and complex design of the respiratory system, there should be no reasonable expectation that DI-induced bronchoprotection in healthy subjects and its absence in asthma, can be traced to a single mechanical and/or biological mechanism. The effect of DI may instead be a balance of favourable and adverse effects. A shift in this balance with disease may facilitate an abnormal response to DI, limiting the beneficial effects of DI and perhaps even promoting bronchoconstriction [39].
Acknowledgments
We would like to thank Z. Hantos (Dept of Medical Physics and Informatics, University of Szeged, Hungary) for critical discussion on the experimental data.
Footnotes
Support Statement
This study was supported by a National Health and Medical Research Council Biomedical Fellowship (number 513921) to P.B. Noble and a University of Western Australia Research Development Award.
Statement of Interest
None declared.
- Received November 22, 2011.
- Accepted February 12, 2012.
- ©ERS 2012