



Regulation of cytokine expression in human plasmacytoid dendritic cells by prostaglandin I2 analogues
- C-H. Hung 1 , 2 , 3 ,
- Y-T. Chu 3 ,
- J-L. Suen 1 , 4 ,
- M-S. Lee 3 ,
- H-W. Chang 3 ,
- Y-C. Lo 5 and
- Y-J. Jong 1 , 2 , 3 , 6
- 1Graduate Institute of Medicine, College of Medicine, 2Dept of Paediatrics, Faculty of Paediatrics, 4Dept of Microbiology, Faculty of Medicine, College of Medicine, 3Dept of Paediatrics, 6Dept of Laboratory Medicine, all Kaohsiung Medical University Hospital, and 5Dept of Pharmacology, Institute of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan.
- Y-J. Jong, Dept of Paediatrics, Kaohsiung Medical University Hospital, No. 100, Tz-You 1st Road, Kaohsiung 807, Taiwan. Fax: 886 73213931. E-mail: yjjong{at}kmu.edu.tw
Abstract
Plasmacytoid dendritic cells (pDCs) are critical in controlling adaptive immunity, but the mechanisms governing cytokine expression remain incompletely defined. Analogues of prostaglandin (PG)I2, such as iloprost, can modulate functions of myeloid dendritic cells, but their involvement in the regulation of human pDCs remains unknown. To this end, the regulatory role of PGI2 analogues on cytokine expression in pDCs was investigated.
Circulating pDCs were magnetically sorted with BDCA-4 cell isolation kits from human peripheral blood mononuclear cells and treated with varying concentrations of iloprost with or without the addition of Toll-like receptor agonists, or an I prostanoid (IP) receptor antagonist, CAY10449. The levels of tumour necrosis factor (TNF)-α, interferon (IFN)-α and interleukin (IL)-10 were measured by ELISA.
Iloprost induced IL-10 expression, but suppressed CpG oligodeoxynucleotide- (or imiquimod-) induced TNF-α and IFN-α production in pDCs. This effect was reversed by the addition of CAY10449. Forskolin, a cyclic adenosine monophosphate activator, conferred a similar modulating effect to that noted in iloprost-treated pDCs, although a higher concentration of forskolin was required to exert the same effect.
Iloprost enhanced interleukin-10 and suppressed Toll-like receptor-mediated tumour necrosis factor-α and interferon-α production of human plasmacytoid dendritic cells via the I prostanoid receptor and, in part, the cyclic adenosine monophosphate pathway.
- Dendritic cell
- iloprost
- interferon-α
- interleukin-10
- prostaglandin
- tumour necrosis factor-α
Dendritic cells (DCs) are highly heterogeneous in terms of their phenotypes and function, and are known to play a major role in initiation and regulation of adaptive immune responses 1–3. The seemingly diverse function of DCs is, in part, dependent on the maturation stage and nature of the stimulating antigen and, importantly, the cytokines secreted upon activation. Notably, activated plasmacytoid DCs (pDCs) are capable of secreting type-I interferon (IFN), such as IFN-α, and are critical in host immunity against viral infection. Also, the role of pDCs in the regulation of various diseases, including allergic, autoimmune diseases and cancer, has been documented. For example, IFN-α is elicited in virus-activated pDCs and participates in the induction of potent T-helper cell (Th) type 1 responses 4. In contrast, tumour-infiltrating pDCs from patients with ovarian cancer promote the induction of regulatory T-cells secreting interleukin (IL)-10, an anti-inflammatory cytokine 5. Interestingly, the products of tumour cells have been shown to inhibit IFN-α production and antigen-presenting capacity by pDCs 6. In a mouse model of asthma, depletion of pDCs led to a polarised Th2 response and the cardinal features of asthma, while transfer of pDCs in naïve mice before sensitisation prevented the occurrence of pulmonary allergic response, suggesting their protective role in lung inflammation 7. It has also been noted that the number of pDCs increased significantly in bronchoalveolar lavage fluid 24 h after allergen challenge in patients with asthma 8. Thus, accumulated evidence to date has indicated the importance of pDCs in the regulation of disease expression and progression and also, depending on the stimulus and the cytokines produced, pDCs can be either immunostimulatory or tolerogenic. While these functional features have been demonstrated, the exact mechanisms by which pDCs are regulated are as yet undefined.
In this regard, it is of interest to note that while prostaglandins (PGs) are initially thought of as pro-inflammatory molecules, prostacyclin (PGI2) has been shown to possess anti-inflammatory activity via the cyclic adenosine monophosphate (cAMP) pathway 9. Because PGI2 is very unstable, PGI2 analogues with a higher chemical stability have been used in clinical application, of which iloprost, a stable PGI2 analogue, is a well-accepted medication for pulmonary arterial hypertension, acting as a vasodilator by binding to the I prostanoid (IP) receptor. Recent evidence from the use of animal models suggests that signalling via this receptor also suppresses the features of asthma via inhibition of airway myeloid DC (mDC) function in vivo 10. It is at present unknown, however, as to whether IP receptor activation and/or iloprost have any functional effect on pDCs. Considering the importance of pDCs in various disease contexts, the current authors have examined the in vitro effects of two PGI2 analogues, iloprost and treprostinil, on the generation of three regulatory cytokines in human pDCs and have provided, for the first time, functional evidence supporting a novel role of PGI2 analogues in the regulation of pDC function.
METHODS
Isolation and analysis of pDCs
Peripheral blood mononuclear cells were isolated from five healthy individuals with no history of systemic disease, after obtaining informed consent. Circulating pDCs were magnetically sorted by centrifugation with BDCA-4 cell isolation kits (Miltenyi Biotec, Bergisch Gladbach, Germany), following the manufacturer’s instructions. Each experiment was repeated three times. In all cases, the purity of isolated pDCs was >92% (see online supplementary material), as judged by their expression of CD123 but not CD14 markers on their surface by flow cytometry (data not shown). Purified pDCs (2×105 cells·condition−1) were treated with varying doses of IP receptor agonists, iloprost or treprostinil, or a cAMP activator, forskolin, for 2 h and then stimulated with 10 μg·mL−1 of CpG oligodeoxynucleotide (ODN)-2216 or imiquimod as previously described 11, 12, for 48 or 72 h. For analysis of IFN-α, pDCs were pre-treated with IL-3 (10 ng·mL−1) 13 for 30 min prior to the treatment with iloprost, followed by the stimulation of the cells with CpG or imiquimod. To examine the involvement of the iloprost–IP receptor axis, pDCs were pre-treated with an IP receptor antagonist, CAY10449, 1 h before the treatment of the cells with iloprost. The production of cytokines in the culture supernatants was determined by ELISA for IL-10, IFN-α and TNF-α (R&D Systems Inc., Minneapolis, MN, USA).
Statistical analysis
All data are presented as mean±sd. Change in cytokines at different doses of iloprost and treprostinil were analysed using the Wilcoxon signed rank test. Differences between experimental and control groups were analysed using the Mann–Whitney U-test. A p-value <0.05 was considered indicative of significant between-group differences.
RESULTS
Iloprost induced IL-10 production, but inhibited Toll-like receptor-mediated expression of TNF-α and IFN-α
To examine the potential effect of PGI2 analogues on the expression of cytokines in pDCs, pDCs from a total of five nonallergic subjects were treated with varying doses of iloprost (10−9–10−7 M) either alone or in combination with Toll-like receptor (TLR) agonists, CpG ODNs or imiquimod, or with varying concentrations of treprostinil. The results showed that iloprost was capable of inducing, in a dose-dependent manner, IL-10 production in pDCs (fig. 1a⇓), whereas no significant enhancement of IL-10 in pDCs was noted when another PGI2 analogue, treprostinil (10−9 and 10−7 M), was used as a stimulus for 48 h (fig. 1b⇓). Interestingly, while iloprost alone was unable to elicit TNF-α expression, iloprost could suppress CpG-induced TNF-α production (fig. 1c⇓). To determine whether iloprost downregulation of CpG-stimulated TNF-α was IL-10 dependent, anti-IL-10 antibody (0.1 μg·mL−1; R&D Systems Inc.) was used for neutralisation of IL-10 bioactivity during the experiment. Neutralised anti-IL-10 antibody could not influence the suppressive effect of iloprost on CpG-stimulated TNF-α expression in pDCs (fig. 1d⇓). Additionally, treprostinil was not effective in modulating the expression of CpG- (or imiquimod-) induced TNF-α production in pDCs (fig. 1e⇓). To investigate the ability of PGI2 analogues to affect IFN-α expression, pDCs were first pre-treated with IL-3, followed by the treatment of the cells with PGI2 analogues and TLR agonists since, in the current authors’ experience, CpG ODNs were unable to induce IFN-α production in pDCs without a brief “priming” by IL-3 13. As shown in figure 2⇓, iloprost (fig. 2a⇓) and treprostinil (fig. 2b⇓) could suppress CpG- or imiquimod-induced IFN-α production in IL-3-primed pDCs. Note that, during this culture period, there was no significant difference in the expression levels of DC maturation markers, including CD86, CD80 and HLA-DR expression, as judged by flow cytometry (data not shown).

Effects of prostaglandin I2 analogues on interleukin (IL)-10 and tumour necrosis factor (TNF)-α expression in plasmacytoid dendritic cells (pDCs). BDCA-4-positive pDCs magnetically sorted from peripheral blood mononuclear cells were treated with varying doses of I prostanoid receptor agonists, iloprost or treprostinil, for 2 h and then stimulated with 10 μg·mL−1 of CpG oligodeoxynucleotide or imiquimod for 48 or 72 h. IL-10 and TNF-α were assessed by ELISA. a) After 48 h (▓) and 72 h (□) of treatment, iloprost enhanced production of IL-10 by pDCs. b) Treprostinil had no effect on IL-10 production after 48 h of treatment. c) Iloprost decreased CpG-induced TNF-α production after 48 h (▓) and 72 h (□), d) even in the presence of an IL-10-neutralising antibody at 0.1 μg·mL−1 for 48 h. e) Treprostinil could not suppress CpG- (░) or imiquimod (▪)-induced TNF-α production after 48 h of treatment. All data are presented in pg·mL−1 per 10−6 cells. #: no CpG; ¶: anti-IL-10 antibody; +: no CpG or imiquimod. *: p<0.05.

Effects of prostaglandin I2 analogues on interferon (IFN)-α expression in plasmacytoid dendritic cells (pDCs). BDCA-4-positive pDCs magnetically sorted from peripheral blood mononuclear cells were treated with varying doses of I prostanoid receptor agonists, iloprost or treprostinil, for 2 h and then stimulated with 10 μg·mL−1 of CpG oligodeoxynucleotide or imiquimod for 48 h. IFN-α was assessed by ELISA. a) Iloprost and b) treprostinil could suppress CpG- (░) or imiquimod (▪)-induced IFN-α production by interleukin (IL)-3-primed pDCs. All data are presented in pg·mL−1 per 10−6 cells. #: no CpG or imiquimod or IL-3 priming. *: p<0.05.
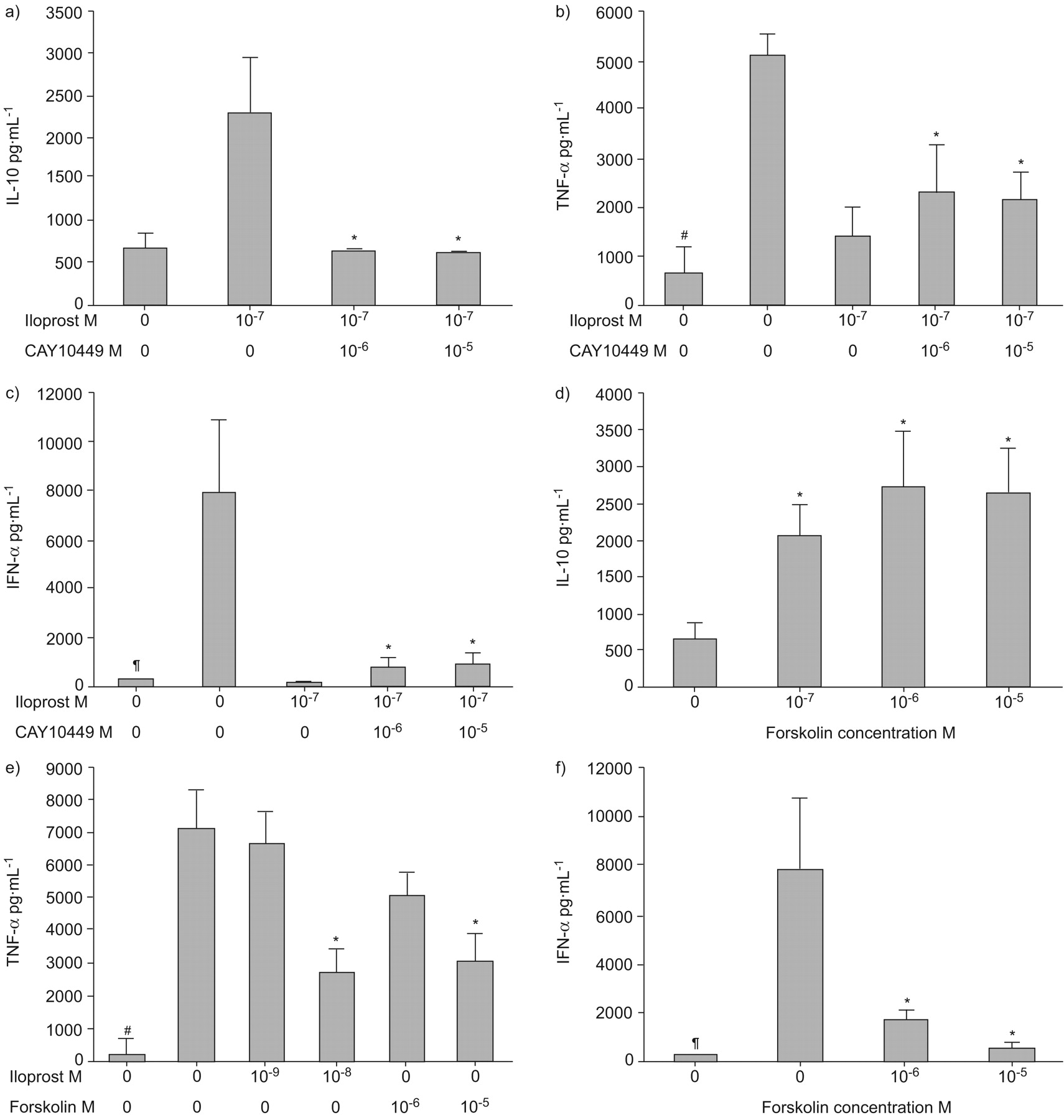
IP receptor antagonist could reverse the effects of iloprost on pDCs
It is known that PGI2 analogues, such as iloprost, exert their function through the IP receptor, leading to increased levels of intracellular cAMP 9. To examine whether, indeed, the effect of PGI2 analogues was mediated through the IP receptor, pDCs were pre-treated with an IP receptor antagonist, CAY10449. The effects of iloprost on the cytokine expression in pDCs were then analysed. As shown in figure 3⇓, the addition of CAY10449 was able to reverse iloprost-enhanced IL-10 expression (fig. 3a⇓), and the inhibitory effect of iloprost on TLR-mediated TNF-α and IFN-α production was abrogated (fig. 3b⇓ and c). Next, it was examined whether increasing intracellular cAMP in pDCs could confer the modulatory effect of iloprost, using a cAMP activator, forskolin. The results showed that treatment of the cells with forskolin showed similar modulatory effects on cytokine expression to those found in iloprost-treated cells. As seen in figure 3d⇓, forskolin was able to enhance IL-10 production, and an inhibitory activity of forskolin on TLR-mediated induction of TNF-α and IFN-α was revealed (fig. 3e⇓ and f). However, on a molar basis, iloprost was shown to be more potent than forskolin on the regulation of cytokine expression in pDCs. These data suggest the involvement of the IP receptor and cAMP pathway in conferring the modulatory effects of iloprost on the expression of cytokines in pDCs.
{kind=link}
{kind=link}
{kind=link}
The effect of iloprost on plasmacytoid dendritic cells (pDCs) was via the I prostanoid (IP) receptor and cyclic adenosine monophosphate (cAMP) pathway. To determine whether the effect of iloprost on interleukin (IL)-10, tumour necrosis factor (TNF)-α and interferon (IFN)-α expression in pDCs was via the IP receptor–cAMP pathway, pDCs were pre-treated with an IP receptor antagonist, CAY 10449, for 1 h before treatment with iloprost or a cAMP activator, forskolin, for 2 h then stimulation with 10 μg·mL−1 of CpG oligodeoxynucleotide for 48 h. For analysis of IFN-α, pDCs were pre-treated with IL-3. CAY10449 could reverse iloprost-enhanced a) IL-10 expression and iloprost-reduced b) TNF-α and c) IFN-α production by pDCs. High doses of forskolin d) enhanced IL-10, e) decreased CpG-induced TNF-α and f) decreased CpG-induced IFN-α production by pDCs, in a similar manner to iloprost. All data are presented in pg·mL−1 per 10−6 cells. #: no CpG; ¶: no CpG or IL-3 priming.*: p<0.05.
DISCUSSION
DCs are emerging as chief orchestrators of immune responses, and two DC subsets, mDCs and pDCs 14, are known to exert their regulatory functions through the release of cytokines following activation and/or inflammatory insults; however, the exact regulatory mechanisms remain to be elucidated. Thus, understanding of the regulatory pathway controlling the expression of cytokines in pDCs would help in advancing the knowledge about the function of pDCs and their involvement in adaptive immunity. Evidence is provided herein supporting a role of the IP receptor and PGI2 analogues in the regulation of pDC function through their modulatory activities on the expression of cytokines, adding a new dimension to the existing regulatory network in pDCs.
PGs are generated by stepwise conversion of arachidonic acid into a series of products, including PGG, PGH, PGD2, PGE2, PGF2α and PGI2, via the action of the cyclooxygenases and other specific enzymes 15, 16. PGs are generally regarded as pro-inflammatory molecules, but PGE2 can have anti-inflammatory and anti-asthmatic effects via the E prostanoid receptor 3 17. PGI2 is a lipid mediator with vasodilatory and anti-thrombotic effects, used in the treatment of vasoconstrictive/ischaemic diseases, including pulmonary arterial hypertension. Inhaled iloprost, a stable PGI2 analogue, is used clinically as a drug treatment for patients with pulmonary arterial hypertension. The present results showed that treatment of pDCs with iloprost resulted in differential effects on cytokine expression. Iloprost inhibited the production of TNF-α, but increased IL-10 production by pDCs. TNF-α may induce acute phase response protein and also play an important role in asthma 18. IL-10 plays an important role in suppression of inflammatory responses by inhibiting multiple pro-inflammatory cytokine expression. Therefore, the functional effect of iloprost in suppressing TLR-mediated pro-inflammatory cytokine expression while increasing IL-10 production in pDCs suggests its anti-inflammatory function.
The findings that CAY10449, an IP receptor antagonist, could reverse, at least in part, the effects of iloprost suggest the involvement of the IP receptor and cAMP pathway in mediating the effect of iloprost. As a corollary, iloprost has been shown to modulate cytokine response via the IP receptor, cAMP and nuclear factor-κB pathways in mouse bone marrow-derived myeloid-type DCs 9. However, treprostinil, another IP receptor antagonist, could not enhance IL-10 production in pDCs. Indeed, different PGI2 analogues have been shown to exert differential effects involving different PG receptors. For example, it is noted that iloprost failed to suppress alveolar macrophages, while treprostinil, a longer-acting PGI2 analogue, inhibited phagocytosis, bacterial killing and cytokine generation in alveolar macrophages to a much greater degree than the other PGI2 analogues and more closely resembled the effects of PGE2 19. Furthermore, iloprost is able to bind with higher affinity to the IP receptor 20. Thus, the differential activity of iloprost versus treprostinil analogues seen in the present study may reflect their difference in the nature and type of receptor occupancy. It is of note that, when the levels of TLR-mediated TNF-α and IFN-α production in pDCs were analysed, iloprost and treprostinil revealed difference, both quantitatively and qualitatively, in their capacities to modulate these two cytokines. It is also noteworthy that, unlike in the case of viral infection, CpG ODN-mediated IFN-α production in pDCs requires an initial priming of the cells with IL-3 11, 13, although the molecular basis for this requirement remains to be elucidated. The current results demonstrated that, in the presence of iloprost (or treprostinil), TLR-mediated IFN-α expression was inhibited in IL-3-primed pDCs. While, at present, it cannot be conclusively discerned whether the suppressive effect of PGI2 analogues on IFN-α production in pDCs is, in part, via its ability to downregulate IL-3 signalling, preliminary data suggest an alteration of IL-3-mediated activation of STAT5 (signal transducer and activator of transcription) in iloprost-treated pDCs (C-H. Hung; personal communication). These results suggest, therefore, the existence of a complex regulatory pathway involved in cytokine regulation in pDCs, and the detailed signalling events associated with differential cytokine responses await further investigation.
It is of interest to note that, in mice depleted of pDCs, iloprost suppressed the cardinal features of asthma via inhibition of airway mDC function through, in part, the induction of IL-10 10. The fact that the role of pDCs in the regulation of pulmonary allergic responses has been shown 7, and that iloprost also targets pDCs and induces IL-10 production, suggests a possible involvement of pDCs in contributing to the regulatory function of iloprost seen in the lungs. Therefore, iloprost, a well-accepted medication, may enhance tolerogenic function of both pDCs and mDCs by enhancing IL-10 and suppressing TNF-α production, suggesting its potential utility in asthma therapy. It is noted, however, that iloprost was able to suppress TLR-mediated induction of another regulatory cytokine, IFN-α, in pDCs. IFN-α from pDCs has been suggested to play a role in conferring the protective effect in pulmonary allergic inflammation and, in fact, significantly lower amounts of virus-induced IFN-α is seen in allergic asthmatic patients when compared with healthy children or those with intrinsic asthma 21. Further work is, thus, required to evaluate the potential use of iloprost and/or IP agonists in asthma. Interestingly, type-I interferons, including IFN-α, and TNF-α are critically involved in the pathogenesis of autoimmune diseases and may be a target of treatment in systemic lupus erythematosus and rheumatoid arthritis 22, 23. The results of the present study showed that iloprost could significantly suppress TNF-α and IFN-α and induce IL-10 expression in pDCs, suggesting the candidacy of iloprost as a modulatory agent in these diseases.
Thus, how I prostanoid receptor signalling integrates in the Toll-like receptor-mediated activation of plasmacytoid dendritic cells may represent a novel mechanism in plasmacytoid dendritic cell function, and the current results would offer an experimental basis for prostaglandin I2 analogues and the I prostanoid receptor signalling pathway as potential targets for modulation.
Support statement
Supported by a grant from the Centre of Excellence, Environmental Medicine, Kaohsiung Medical University Research Foundation, No. KMU-EM-97-2.2a, and a grant from Kaohsiung Medical University Hospital, No. KMUH96-6R03 (both Kaohsiung, Taiwan).
Statement of interest
None declared.
Footnotes
-
This article has supplementary material accessible from www.erj.ersjournals.com
- Received May 6, 2008.
- Accepted September 27, 2008.
- © ERS Journals Ltd