



Lysosomal cysteine proteases in the lung: role in protein processing and immunoregulation
- 1Institute of Immunology, and 2Dept of Pneumology and Critical Care, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany
- T. Welte, Dept of Pneumology and Critical Care, Otto-von-Guericke-University Magdeburg, Leipziger-Str. 44, 39120 Magdeburg, Germany. Fax: 49 39167190466. E‐mail: tobias.welte@medizin.uni-magdeburg.de
Abstract
Lysosomal cysteine proteases are a family comprising >10 enzymes. For many years it was believed that these enzymes catalyse protein breakdown unselectively, are highly redundant in their substrate specificity and are also expressed ubiquitously.
This view has changed dramatically since a number of new lysosomal cysteine proteases with restricted expression and outstanding enzymatic activity have been described. In addition, knockout mice and selective protease inhibitors have been used to characterise specific functions of single proteases.
In this review, some of these functions are discussed in relation to the lungs, especially the role of lysosomal cysteine proteases in matrix remodelling, immunoregulation and surfactant protein processing.
- Cathepsin
- fibrosis
- lung
- protease
- surfactant
Under normal conditions, up to 50% of proteins are degraded within the lysosomal compartment 1. This finding, along with the short intralysosomal half-life of proteins, demonstrates the high activity of lysosomal proteases 2. Lysosomal proteolysis is limited by the activity of endoproteases that degrade internalised proteins into polypeptides. The terminal processing of these polypeptides is carried out by exopeptidases 3. For many years it was believed that the lysosomal breakdown was a rather unselective process carried out by a number of highly redundant proteases that are expressed ubiquitously. However, this view has recently changed dramatically as knockout mice and selective protease inhibitors have been used to characterise specific functions of single proteases 4.
Proteases are classified, based on their catalytic mechanisms, into serin, aspartic, metallo, threonin and cysteine proteases. This overview focuses on cysteine proteases. In general, the term “protease” is used, which is synonymous with “peptidase”, because it is commonly used in the medical literature. Proteases are enzymes that hydrolyse peptide bonds. They can be subdivided into exopeptidases, enzymes which act near a terminus of the polypeptide chain, and endopeptidases, which cleave internal peptide bonds. The old term “proteinase” carries the same meaning as “endopeptidase”. The most comprehensive overview concerning the classification of proteases and their inhibitors has been compiled in the MEROPS database 5.
The term “cathepsin”, which was introduced in 1920 6, stands for “lysosomal proteolytic enzyme”, regardless of the enzyme class. Therefore, in addition to cysteine proteases, this term also includes serine proteases (cathepsins A and G) and aspartic proteases (cathepsins D and E). Sequence homologies of the cysteine proteases (cathepsins B, C, H, L and S) indicate that these enzymes diverged early during eukaryotic evolution 7. Some of the genes encoding novel enzymes, e.g. cathepsins F, K, W and X, may be the result of relatively recent gene duplication events, as suggested by three gene pairs with common chromosomal localisation and high sequence homologies 8.
Cathepsins are synthesised as inactive preproenzymes and are glycosylated post-translationally. They are directed towards the lysosomal compartment using cellular mannose‐6‐phosphate receptors. Some cells can secrete significant amounts of mature and procathepsins 9, 10.
Cathepsins are mostly monomeric proteins. Several three-dimensional structures of cysteine proteases are available and demonstrate that enzymes of the papain family share a common fold 4, 11, 12. The main structural features of a papain-like two-domain catalytic platform are depicted in figure 1⇓ using a model of mature cathepsin S; the active site is located at the interface between the two domains. Most cysteine proteases of the papain family are endopeptidases “by design”. Exopeptidases, such as cathepsins B, C, H and X, are likely to have evolved from the endopeptidase template by addition of structural elements capable of interacting with the C‐ or N‐terminus of a substrate 13–15.

Secondary structure representation of mature cathepsin S. Catalytic residues are highlighted as capped sticks. Figure kindly provided by W. Brandt, Martin-Luther-University Halle, Halle, Germany.
The substrate-binding region of proteases is defined as an arrangement of binding pockets or subsites (S) for peptide substrate amino acids (P) on both sites of the scissile bond. To date, seven such sites have been defined for papain-like cysteine proteases. The primary substrate specificity of cathepsins is defined by the S2 binding pocket 16, 17. However, recent studies support the importance of S3 and S4 sites for the determination of substrate specificity 18, 19. Most lysosomal cysteine proteases accept hydrophobic amino acid residues, such as phenylalanine and leucine, at the S2 position. In contrast to other cathepsins, cathepsin B also accepts arginine in this position.
The exceptional enzymatic activity of cathepsin K is based on its ability to cleave protein substrates with proline residues in the P2 position 20. Thus, cathepsin K cleaves collagen I and II fibres at different sites in both the telopeptide and the helical regions 21. In contrast, cathepsin L catalyses the hydrolysis of collagen in the telopeptide region only, whereas matrix metalloproteases (MMPs) mediate the cleavage at a single site within the helical region. Before the discovery of cathepsin K, it was suggested that collaboration of different proteases is a precondition for collagen degradation. It was thought that cathepsin L‐like proteases were involved in destabilising the collagen fibres and making them accessible for further breakdown by MMPs 22. However, now it is clear that cathepsin K is the first mammalian protease to be described that is capable of independent collagen breakdown. Similar activities have previously only been described for bacterial proteases. The importance of cathepsin K in matrix metabolism in vivo is supported by the finding that inactivating mutations in the cathepsin K gene render osteoclasts, which express large amounts of cathepsin K, dysfunctional and lead to the hereditary disease pycnodysostosis 23. The functional relevance of cathepsin K at other expression sites is less well understood.
A number of studies have investigated elastin degradation by lysosomal cysteine proteases and showed that cathepsin K is one of the most potent elastases described to date 24. In addition, the elastinolytic activities of cathepsins L and S are, at optimal pH, higher than the activities of neutrophil elastase and metalloelastase 25. Both the collagenolytic and elasinolytic activities of cathepsins appear to be important for matrix remodelling in the lungs.
Regulation of enzymatic activity
The activity, and thus the functional importance of cathepsins, is regulated at different levels (fig. 2⇓). Initially, it was believed that cathepsins were “housekeeping” proteins and therefore ubiquitously expressed. This view has evolved as cathepsins with distinct expression patterns have been described. Examples include: cathepsin K, which is found in large amounts in osteoclasts, activated macrophages and some epithelial cell–s 26; cathepsin S, found in antigen-presenting cells 31, 32; and cathepsin W, which has been detected in cytotoxic lymphocytes 33–35. The same view is also reflected in the lungs (fig. 3⇓), where some cathepsins are expressed more or less ubiquitously, for example cathepsins B, H and L, whereas others are expressed either in macrophages (cathepsin S) or in bronchial epithelial cells (cathepsin K). Of note are type II pneumocytes, which express significant amounts of cathepsin H, but low levels of the other cathepsins (table 1⇓) 28, 36, 37.
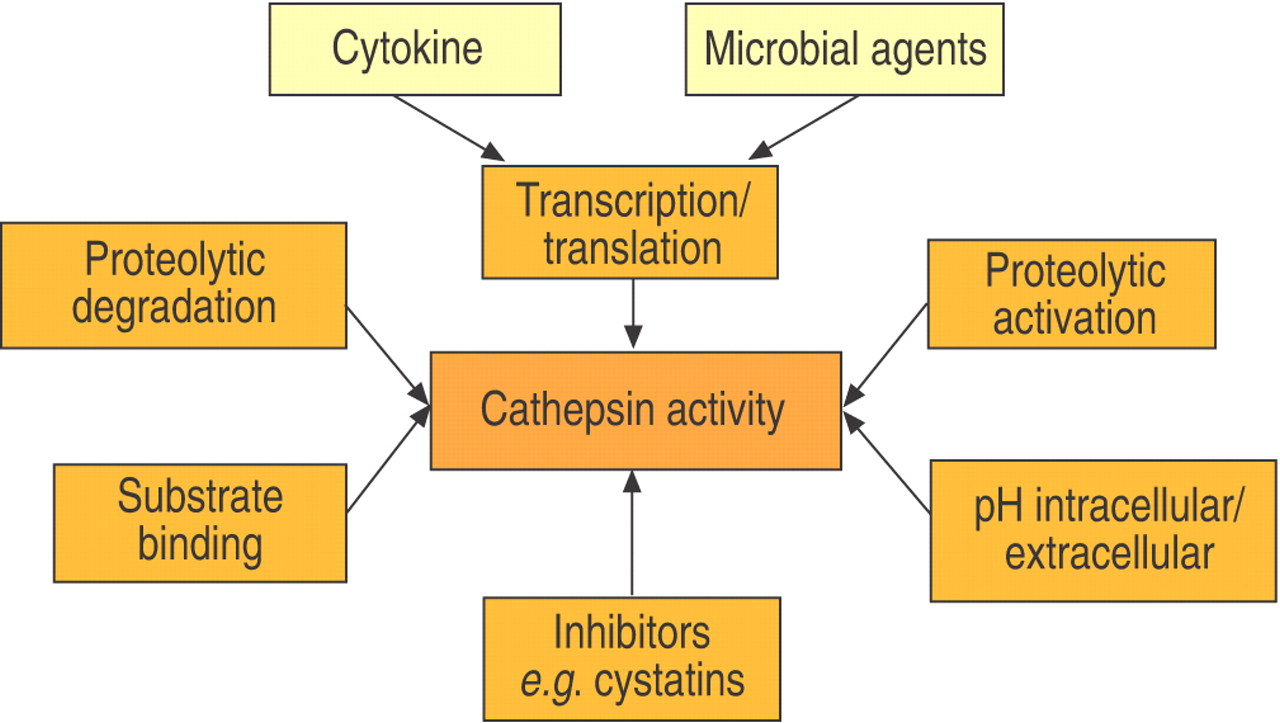
Regulation of enzymatic cathepsin activity.

Cathepsin expression in adult human lung tissue. a) Cathepsin B was ubiquitously expressed. b) Cathepsin H was expressed in macrophages (AM) and type‐II‐pneumocytes (TIIP). c) Cathepsin K immunoreactivity was found predominantly in bronchial epithelial cells (BE). d) Cathepsin L was expressed in BE and in AM. e) and f) Cathepsin S and cathepsin W were found in AM (kindly provided by S. Krüger, Institute of Pathology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany). g) Cathepsin X was ubiquitously expressed. Anti-cathepsin L, S and W antibodies were kindly provided by E. Weber (Institute of Physiological Chemistry, Martin-Luther-University Halle, Halle, Germany) and T. Wex (Dept of Gastroenterology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany).
Immunoreactivity in lung tissue and enzymatic activity of cathepsins
Furthermore, cathepsin expression and activity are regulated by cytokines and microbial agents. Interferon (IFN)‐γ and interleukin (IL)‐6 stimulate expression of cathepsins B and L, whereas transforming growth factor‐β1 and IL‐10 inhibit enzymatic activity in most cells 38–41. Stimulation of macrophages by bacterial lipopolysaccharide also leads to increased cathepsin expression 42, 43.
The majority of enzymes, including cathepsins, are synthesised as inactive precursors. In addition to being selective competitive inhibitors of the parent enzyme, the proregions appear to be important for the correct folding of the newly synthesised polypeptide chain and for stabilising the protein against denaturation caused by neutral or slightly alkaline pH 44. The removal of propeptides is a precondition for enzymatic activation 13, 45, 46–49. The enzymatic activation of cathepsins is achieved by other cathepsins or by autocatalysis. Exopeptidases, such as cathepsin C or X, are activated exclusively by other lysosomal endoproteases, and proteolytic activation depends on pH and the concentration of glucosaminoglycans 15, 50, 51.
Furthermore, cathepsins are controlled by proteolytic degradation and inactivation of the proteases themselves. For example, cathepsin L is inactivated by cathepsin S 52 and the inhibition of lysosomal cysteine proteases leads to delayed cathepsin D processing 53.
Cathepsins have an acidic pH optimum. In general, environments with neutral or alkaline pH lead to inactivation of cathepsins. However, the rate of inactivation varies among the different members of the protease family. Cathepsin L is inactivated rapidly, whereas cathepsin S retains most of its activity for >1 h. Cathepsins K and V have an intermediate sensitivity to increasing pH 27, 54, 55. In addition, recent investigations have shown that cathepsins are stabilised after formation of complexes with potential substrates. This is illustrated by the findings that inactivation of cathepsin L at neutral pH is prevented by binding of the p41 protein and that cathepsin B is stabilised extracellularly by high protein concentrations 56, 57.
Finally, because of the exceptional proteolytic activity and the high concentration of cathepsins, up to 1 mM in lysosomes, their activity is controlled by a network of endogenous inhibitors. The activity is mostly controlled by cystatins, which are expressed intracellularly (cystatins A and B) or extracellularly (cystatins C, D, and F) 58. Other endogenous cathepsin inhibitors include kininogens, serpins and α2‐macroglobulin 59–61.
In vivo functions of cathepsins
Cathepsin knockout mice
During the last 8 yrs the phenotypes of seven different cathepsin knockout mice have been described. In addition to the aspartic proteases, cathepsins D and E, the genes of the lysosmal cysteine proteases of the papain family, cathepsins C, B, L, K and S, have been inactivated 62–69.
In one study, cathepsin B knockout mice displayed no obvious phenotype 70, but detailed analyses of thyroid function in cathepsin B‐deficient mice revealed defects in thyroglobulin processing 71. In addition, after caerulein challenge, these mice were less susceptible to the induction of acute pancreatitis; pancreatic damage was 50% lower in cathepsin B‐deficient animals compared with control animals 63. Furthermore, recent investigations have demonstrated that cathepsin B is involved in the induction of hepatocyte apoptosis; genetic or pharmacological inhibition of cathepsin B reduced the amount of apoptotic hepatocytes, and thus inflammation and fibrosis after bile duct ligation 72.
Cathepsin C (dipeptidyl peptidase I) knockout mice have displayed severe defects in the induction of target cell apoptosis by cytotoxic T‐cells. As cathepsin C plays an essential role in the in vivo processing and activation of granzymes A and B, which are required for cytotoxic lymphocyte granule-mediated apoptosis, these defects are comparable with those detected in perforin- or granzyme A‐/B‐deficient cytotoxic lymphocytes 73. Furthermore, defects in cathepsin C activity may have severe consequences for antiviral protection and the removal of tumour cells. More recently, it was shown that cathepsin C knockout mice are protected against acute arthritis; this protective function was linked to defects in the proteolytic activation of cathepsin G, neutrophil elastase and proteinase 3 62. The question of whether this incomplete activation, especially that of leukocyte elastase, is associated with defects in the antibacterial response remains to be resolved 74, 75. In addition, cathepsin C activates mast cell chymases in vivo 76 and is therefore involved in the regulation of allergic reactions.
Cathepsin K‐deficient mice display an osteopetrotic phenotype, with excessive trabeculation of the bone marrow space. This phenotype could be related to alterations in the function of osteoclasts 67. Accordingly, histomorphometric analyses of cathepsin K‐overexpressing mice revealed significantly decreased trabecular bone volume and acceleration of bone turnover 77. However, the phenotype displayed by cathepsin K knockout mice is mild compared with the human cathepsin K deficiency, pycnodysostosis. Pycnodysostosis is associated with severe skeletal dysplasia, including short stature and dysplasia of cranial bones. Therefore, it was suggested that there is only a partial functional overlap in the functions of human and mouse cathepsin K.
The most prominent phenotype was found after inactivation of cathepsin L, which underlines the well-known high endoproteolytic potential of this ubiquitous protease. Cathepsin L gene (CTSL)‐deficient mice develop periodic hair loss and epidermal hyperplasia, acanthosis and hyperkeratosis. The hair loss is due to alterations in hair follicle morphogenesis and cycling, dilatation of hair follicle canals, and disturbed club hair formation. Hyperproliferation of hair follicle epithelial cells and basal epidermal keratinocytes are the primary characteristics underlying the mutant phenotype 64. The phenotype of CTSL‐deficient mice is reminiscent of the spontaneous mouse mutants furless and nackt 78, 79, which also involve the cathepsin L gene. Recently, it has been found that older CTSL knockout mice develop a dilated cardiomyopathy associated with interstitial fibrosis in the myocardium 80. In addition to these findings, recent publications have documented the function of CTSL in prohormone processing. In thyroid and neuroendocrine cells of cathepsin L‐deficient mice there was a distinct accumulation of the prohormones thyroglobulin and proenkephalin, respectively 71, 81.
In vivo, the crucial role of cathepsins in immunoregulation was first demonstrated in cathepsin L knockout mice 82.
Cathepsins in antigen presentation
Extracellular antigens, which are taken up by professional and nonprofessional antigen-presenting cells, are mostly degraded in the endosomal/lysosomal compartment. The resulting peptides are displayed on the cell surface after formation of major histocompatability complex (MHC) class II‐peptide complexes. MHC class II αβ heterodimers are synthesised in the endoplasmic reticulum. During the early stages of biosynthesis, the αβ heterodimers associate with a type II membrane protein, the invariant chain (Ii), to form a nonameric complex of αβIi. Ii associates with class II molecules via direct interaction of its luminal domain with the antigen-binding groove of class II molecules. The αβIi complex traverses the Golgi apparatus and is delivered to the endosomal/lysosomal compartment were Ii is degraded and liberated from the αβIi complex. This allows the class II molecules to encounter and bind the antigenic peptides. Proteolysis participates in generation of class II‐peptide complexes at two critical points: proteolytic degradation of Ii, which is a prerequisite of peptide loading; and generation of the antigenic peptides from larger proteins.
Cathepsin L knockout mice show reduced positive selection of CD4+ T‐cells, which is caused in part by a block in late Ii processing steps, and, therefore, decreased loading and presentation of class II complexes with self peptides 82. Additionally, evidence was found in cortical thymic epithelial cells that lack of cathepsin L activity shifts the pattern of self antigens presented by the class II complexes 83. Therefore, T‐cells that have been positively selected against peptides presented by cortical thymic epithelial cells fail to pass the negative selection against the peptides presented by dendritic cells. This is because the latter employ cathepsin S for antigen processing and therefore present the common pattern of self antigens. The finding that different antigen-presenting cells use different cathepsins for processing of Ii and antigens was later supported by data derived from cathepsin S knockout mice, indicating that cathepsin S is a major Ii-processing enzyme in B‐cells and dendritic cells 66. The finding that mouse macrophages are using predominantly cathepsin F for Ii processing and class II peptide loading is not yet proven in knockout mice 84.
The immunoregulatory function of defined cathepsins in mice is not necessarily identical to the function of similar enzymes in humans. For example, it was suggested that cathepsin V replaces cathepsin L in human cortical thymic epithelial cells 55. Cathepsin V is highly expressed in human thymus, testis and corneal epithelium 55, 85. However, the mouse genome does not contain a cathepsin V gene. In addition, by exploring human epithelial cells overexpressing the proteins of the MHC class II complex, it was shown that only human cathepsin S, and not human cathepsin L, was able to process Ii and thus to play a pivotal role in antigen presentation by epithelial cells 32.
The suppression of cathepsin F, but not of cathepsin S, in the lungs prevents eosinophilia and immunoglobulin E response after ovalbumin challenge 84. In mice, alveolar macrophages express large amounts of cathepsin F, which suggests a pivotal role for these cells in immunoregulation.
Further studies should characterise the expression and function of additional cysteine proteases, e.g. cathepsins B, K and L, in antigen-presenting cells of the lungs and also explore in more detail the effects of selective inhibition of these enzymes on the development of allergic reactions.
Cysteine proteases in matrix remodelling of the lung
The extracellular matrix (ECM) of most tissues contains a complex mixture of collagens, proteoglycans and elastin. The pulmonary ECM is subjected to a continuous turnover of >10% of total ECM per day 86. For complete degradation, the concerted action of several hydrolases, both inside and outside cells, is necessary. Proteolytic processing is needed for maturation and controlled deposition of ECM. Extracellularly, collagens are degraded by collagenases, gelatinases and stromelysin. These proteases cleave collagen helices at a defined site, leaving 2/3 and 1/3 fragments to be phagocytosed by cells such as fibroblasts and macrophages 22. Within these cells, the matrix-containing phagosomes fuse with lysosomes in which cathepsins complete the degradation.
In addition to intracellular degradation of collagens, cathepsins can also degrade matrix proteins extracellularly. Cells found to release cathepsins include macrophages (cathepsins B, K, L and S) 87, mast cells (cathepsins C and L) 88, smooth muscle cells (cathepsins K and S) 89, fibroblasts (cathepsins B and K) and tumour cells (cathepsins B, L and S) 90–92. Most cathepsins have optimal activity at an acidic pH. Some cathepsins, e.g. cathepsin L, lose their activity quickly after incubation at a neutral pH. However, several recently described cathepsins maintain their enzymatic activity even after incubation at neutral pH and thus can degrade matrix proteins under physiological conditions. Cathepsins C and S have been found to have the highest pH stability 54, 88, while cathepsins K and V display intermediate pH stability 27, 55. In vitro investigations have shown that cathepsin K can degrade fibril proteins at neutral pH 93.
Furthermore, cells releasing cathepsins might promote extracellular proteolysis by direct acidification of the pericellular space. For example, osteoclasts release their lysosomal contents into the resorption lacunae 94. In addition, Punturieri et al. 87 have reported the acidification of the pericellular environment by macrophages. In vitro, monocyte-derived macrophages adhere tightly to elastin particles and form a sequestrated environment, which is then acidified by using a vacuolar type H+‐ATPase to pump protons from the cytoplasmic to the extracellular space. Similar mechanisms may be involved in matrix remodelling by lung macrophages and may also contribute to the pathogenesis of granulomatous lung diseases, which are characterised by the appearance of multinucleated giant cells (MGC). Recently it was shown that MGC express and release large amounts of cathepsin K 29, 93, 95. Conversely, it was shown that MGC can enclose foreign materials, micro-organisms or other extracellular components. Therefore, it is likely that MGC are also generating an acidified micro-environment and use cathepsins for extracellular degradation.
Chronic obstructive lung disease (COPD) is a generic term covering several clinical entities, including emphysema and chronic bronchitis. Emphysema is characterised by abnormal permanent enlargement of the air space distal to the terminal bronchiole accompanied by destruction of alveolar walls. It has been shown recently that lymphocytes and activated macrophages are involved in the regulation of COPD and the development of emphysema.
Initiated by the finding that intratracheal instillation of cathepsin B in an animal model caused a phenotype that resembles human emphysema 96, a number of studies were performed to characterise the putative role of lysosomal cysteine proteases like cathepsin B and L in the development of emphysema 97, 98. These proteases were found to be strongly expressed in activated macrophages. It was shown that the concentration of cathepsin L was significantly increased in the bronchoalveolar lavage fluid of smokers with emphysema 36, 99.
Based on the hypothesis that the cytokine-regulated activation of macrophages leads to cathepsin release, Wang et al. 100 and Zheng et al. 101 established transgenic murine models employing the inducible expression of cytokines along the alveolar surface; the overproduction of IL‐13 or IFN‐γ caused a phenotype that mirrors human COPD, with emphysema, enlarged lungs and enhanced pulmonary compliance. These changes were accompanied by increased expression of MMPs and cathepsins in the lungs. When mice were given the cysteine protease inhibitor E64, they showed markedly attenuated emphysematous changes, implying that cysteine proteases are important in the development of cytokine-induced emphysema 101. Further investigations should determine whether induction of cytokines on a cathepsin-deficient background attenuates the development of emphysema.
A further question to be addressed is whether cathepsins influence the development of lung fibrosis. The current authors have previously shown that activated pulmonary fibroblasts express large amounts of cathepsin K. The cathepsin K expression in fibroblasts derived from patients with interstitial pulmonary fibrosis or bronchiolitis obliterans obstructive pneumonia was increased compared with fibroblasts from normal lung tissues. Using the model of bleomycin-induced lung fibrosis, an induction of cathepsin expression was detected after bleomycin challenge. Using cathepsin K knockout mice, increased lung fibrosis was found after bleomycin challenge. From these results, the authors concluded that cathepsin K may be involved in matrix degradation and thus may counteract the increased matrix deposition responsible for the development of lung fibrosis.
Lung cancer cells can secrete cathepsins L and B in vitro 90, 92 and anti-cathepsin L antibodies protect against the formation of solid tumours by implanted myeloma cells in vivo 102. Therefore, it was suggested that cathepsins may play a critical role in tumour invasion and progression. In addition, it was shown in a number of in vitro models using cells of different origins that inhibition of cathepsin B or cathepsin L by specific inhibitors, or suppression of protease expression using antisense technologies, decreases the invasion of tumour cells into the ECM 103–105. Conversely, the present authors' recent investigations indicate that cathepsin K does not play a role in the progression of lung tumours 106. It was found that the expression of cathepsin K is downregulated in lung tumours on a per cell basis when compared with differentiated bronchial epithelial cells. Therefore, future studies exploring tumour models in cathepsin knockout mice should provide more definite answers to the question of which cathepsins are really involved in the progression of lung tumours.
Processing of surfactant proteins by cysteine proteases
Pulmonary surfactant is a mixture of lipids and specific proteins, which is secreted into the alveolar space by the type‐II pneumocytes. Its main function is the reduction of surface tension at the air-liquid interface, thus preventing alveolar collapse at end-expiration. The surfactant proteins (SP)‐B and ‐C play an essential role in the metabolism and dynamics of the pulmonary surfactant lipids (reviewed in 107). Hereditary SP‐B deficiency in infants or mice leads to respiratory failure at birth 108–111. Mutations in the human SP‐C gene are associated with interstitial lung disease 112, 113. In a defined genetic background, SP‐C knockout mice develop chronic pneumonitis and emphysema 114. In addition, both hereditary alveolar proteinosis in babies without detectable mutations in the SP‐B or the SP‐C gene and acquired pulmonary alveolar proteinosis in children and adults are characterised by intra-alveolar accumulation of mature surfactant proteins and abnormal SP‐B precursors. SP‐B precursors only are detected in babies with congenital surfactant defects characterised by the absence of lamellar bodies in type‐II pneumocytes (F. Brasch, Institute of Pathology, University Hospital “Bergmannsheil”, Bochum, Germany, personal communication). Therefore, insufficient processing of proSP‐B or proSP‐C due to a lack or dysfunction of one or more proteases involved in surfactant protein processing might be another, yet undiscovered, cause of surfactant dysfunction in pulmonary diseases in babies, children and adults.
The hydrophobic surfactant proteins SP‐B and ‐C are transported via the endoplasmic reticulum, the Golgi system and multivesicular bodies from their site of synthesis at the ribosomes to the lamellar bodies. On this route, the primary translation products, proproteins (proSP‐B, proSP‐C) with molecular masses of 42 and 21 kDa, undergo extensive C‐ and N‐terminal processing in multivesicular bodies and lamellar bodies 115–118. Although various processing steps have been described so far 119, 120, nothing is known about the identity of the proteases involved. As a result of the acidic pH in multivesicular and lamellar bodies, cathepsins are potential candidates for involvement in surfactant protein processing. It has been shown in vitro that cathepsin D or a “cathepsin D‐like protease” catalyses the post-translational processing of the hydrophobic proSP‐B 121, but cathepsin D itself was not detected in type‐II pneumocytes and no specific activity was found in isolated lamellar bodies 122, 123. In contrast, the lysosomal protease cathepsin H was localised in lamellar bodies of type‐II pneumocytes of the rat lung 124. Immunoelectronmicroscopic investigations have shown that cathepsin H, proSP‐B and proSP‐C are colocalised in multivesicular and lamellar bodies of type‐II pneumocytes. Isolated rat lamellar bodies were also shown to express enzymatic cathepsin H activity. In human foetal type‐II pneumocytes, the expression and enzymatic cathepsin H activity was upregulated during in vitro differentiation in parallel with surfactant protein secretion. Incubation of differentiated type‐II pneumocytes with E64, a potent inhibitor of cysteine proteases, interrupts SP‐B processing and leads to additional features of SP‐B deficiency, including abnormal SP‐C processing and aberrant lamellar bodies 125.
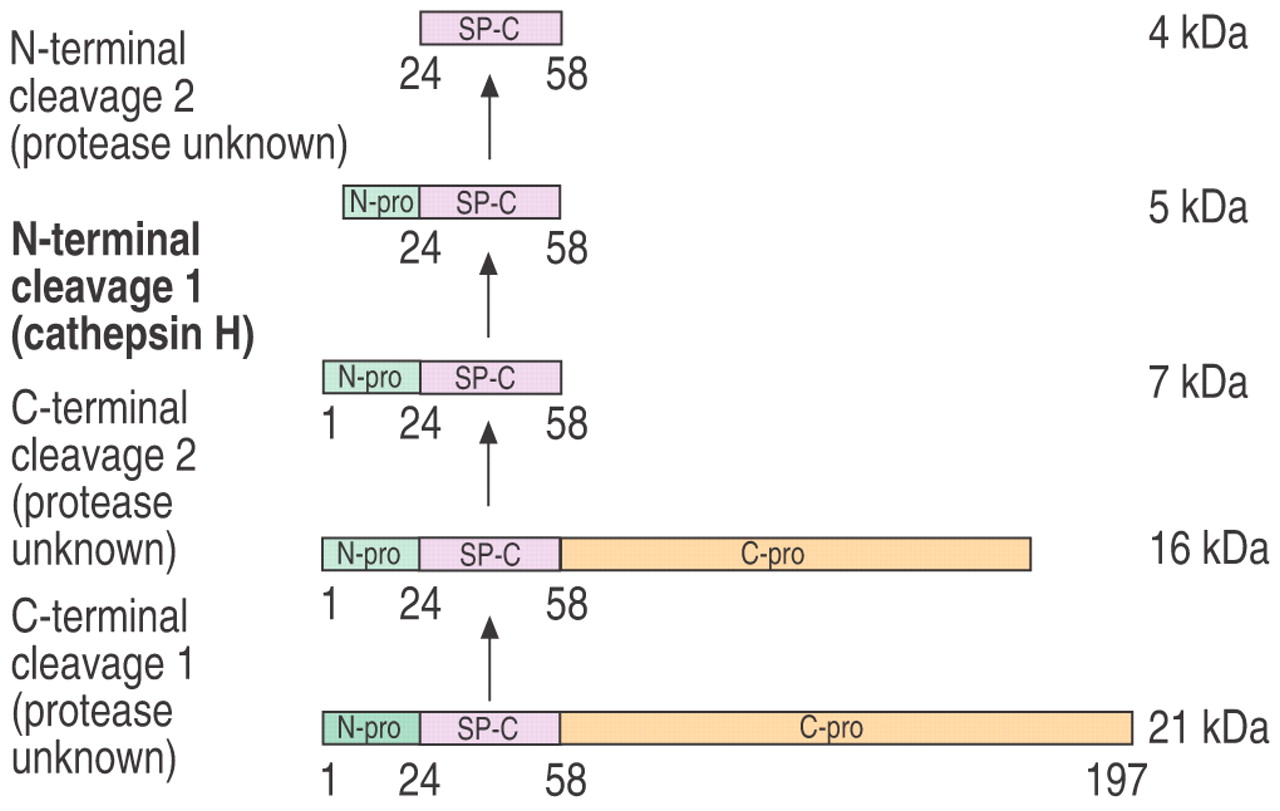
The SP‐C processing by cathepsin H was investigated in more detail in vitro. It was suggested that cathepsin H is involved in the N‐terminal processing of proSP‐C in electron dense multivesicular bodies (fig. 4⇓) 126. It should to be pointed out that, in these studies, the processing of proSP‐C by cathepsin H alone did not result in the mature surfactant proteins. Therefore, additional enzymes also seem to be involved in this complex processing cascade. Studies using knockout mice for different combinations of lysosomal proteases should be initiated to characterise other enzymes involved in surfactant protein processing.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Processing of surfactant protein (SP)‐C precursor by cathepsin H.
Conclusion
Using molecular biological methods, such as knockout mice and antisense approaches, a number of specific functions of cathepsins have been highlighted during recent years. Without doubt, it has been shown that cathepsins play a crucial role in matrix remodelling, immunoregulation and surfactant protein processing. On-going research with animal models and the detailed investigation of human lung diseases will clarify their role in the pathogenesis of different lung diseases.
- Received September 16, 2003.
- Accepted January 6, 2004.
- © ERS Journals Ltd