



The bidirectional capacity of bacterial antigens to modulate allergy and asthma
- H. Renz and
- U. Herz
- H. Renz, Central Laboratory, Hospital of the Philipps University, Baldingerstr., D-35033, Marburg, Germany. Fax: 49 64212865594
Abstract
In recent decades, the prevalence of allergic diseases including bronchial asthma, hay fever and atopic dermatitis, has risen steadily in high-income countries. The underlying mechanisms for this phenomenon remain largely unknown. Since the natural mutation rate is low, altered environmental and lifestyle conditions are thought to play an important role.
Epidemiological and clinical studies have provided indirect evidence that infections may prevent the development of atopy and atopic disease. This is referred to as the “hygiene hypothesis”. According to the hygiene hypothesis, viral and/or bacterial infections could inhibit the T-helper (Th)-2 immune response associated with atopic reactions by stimulating a Th-1 response involved in defence of bacterial infections and delayed-type hypersensitivity reactions.
In particular, the prenatal period and early childhood are considered to be critical for the establishment and maintenance of a normal Th-1/Th-2 balance. On the other hand, several studies suggested that infections exacerbate established allergic diseases, e.g. bronchial asthma, airway hyperresponsiveness and atopic dermatitis. Therefore, viral and/or microbial infections and/or their products may have bidirectional effects on the development of allergy and asthma.
This review will focus on recent findings related to the interaction between allergic disorders and infectious diseases, with the main emphasis on bacterial infections.
- Allergy
- asthma
- atopy
- bacterial antigens
- T-helper 1/T-helper 2 balance
It is well established that allergy and asthma result from a complex interaction between genetic and environmental factors. Although there is no doubt that genetic polymorphisms at multiple sites in the genome contribute to the predisposition for the development of these conditions, the immediate environment also plays a crucial role in determining the time of onset, maintenance and severity of allergy and asthma. The contribution of the environment is underscored by the overwhelming evidence regarding the dramatic rise in allergy and asthma prevalence and incidence particularly in high-income countries around the world 1, 2. Numerous studies have shown a several fold increase in allergic diseases during the last few decades. This increase cannot solely be explained by genetic factors since the natural mutation rate is low and substantial shifts in the genome do not occur over a few generations. Therefore, research has recently focused on the identification and isolation of environmental factors which contribute to this phenomenon. More recently, the so called “hygiene hypothesis” has been put forward to explain the epidemiological data 3, 4.
Allergy and asthma are associated with a profound immunological deregulation characterized by an overproduction of T-helper (Th)-2 T-cells. It is well established that Th-2 effector function is negatively regulated by Th-1 T-cells. Th-1 cells seem to play a critical role in defence against various microbial antigens including bacterial, viral and fungal infections. The hygenie hypothesis suggests that a reduced frequency of infections, less severe infection and prevention of infection by e.g. frequent use of antibiotics would prevent maturation of Th-1 immunity and, therefore, would give rise to a allergen-specific Th-2 immune response following natural exposure to allergens.
It is the purpose of this review to discuss the evidence both in favour and against the hygiene hypothesis, focusing on bacterial infections and, to a lesser extent, on viral antigen exposure.
The concept of T-helper 1, T-helper 2 and T-helper 3 immune deviation
During the last two decades, the cellular and molecular mechanisms of allergic inflammation have been revealed to a large extent. A breakthrough in understanding the regulation of allergic immune responses came with the discovery of different T-cell effector subsets termed T-helper (Th)-1 and Th-2 5, 6. Both T-cell subsets are characterized by a unique pattern of cytokine production. Th-1 cells produce interferon (IFN)-γ, interleukin (IL)-2 and tumour necrosis factor (TNF)-β. In contrast, Th-2 cells are defined by the exclusive production of IL-4, IL-5, IL-6, and IL-10 5, 6 (fig. 1⇓). Th-1 cells play an important role in host defence against a wide range of microbes including viral, bacterial, and fungal antigens, contribute to the production of immunoglobulin-(Ig)M and IgG antibodies, the formation of granuloma and the development of cytotoxic T-cell responses. Recently, the Th1/Th2 concept which was originally exclusively used to describe the CD4 T-cell subset was extended to include CD8 T-cells as well 7, 8. In contrast, Th-2 T-cells play a critical role in the defence of helminthic infections. This type of infection requires recruitment and activation of eosinophils as well as formation of IgE antibodies 9, 10.
Further studies have addressed the question whether deregulation in T-cell subset formation may play an important pathophysiological role in certain diseases. Since allergic diseases are characterized by production of IgE and local recruitment of eosinophils, it was no surprise that extensive investigations carried out in patients suffering from respiratory and skin allergies revealed the importance of the Th-2 T-cell subset in allergic inflammation. These cells control regulation of IgE production via IL-4 and IL-13 release. Furthermore, certain aspects of eosinophil and mast cell biology are also closely monitored by Th-2 cells including maturation, recruitment, and activation. In addition, IL-5 represents an important survival factor for the eosinophils via prevention of apoptosis 11, 12.
Recently, a third T-cell subset has been discovered termed Th-3 (fig. 1⇓). This T-cell subset is defined by the production of transforming growth factor (TGF)-β in addition to IL-4 and IL-10 13–15. The TGF-β-superfamily consists of a number of structurally related regulatory proteins which include besides TGF-β1-TGF-β5 the inhibins, activins, bone morphogenic and related morphogenic proteins, all of which are of significant importance in many areas of cell biology 16. The five isoforms of TGF-β share biological activities. The prototype of this family is TGF-β1 a cytokine that was originally purified from human platelets. Interestingly, human platelets represent the most concentrated natural source of TGF-β 17. In mammalian species only TGF-β1, TGF-β2, and TGF-β3 have been identified and cloned.
TFG-β is produced by every cell lineage and exhibits a wide variety of partially antagonistic and diverse functions (table 1⇓) 16. TGF-β plays an important role in immune activation (upregulation of adhesion molecules, enhanced chemotaxis and enhanced production of IgA) as well as in immune deactivation. Under certain conditions, TGF-β inhibits proliferation of CD4 as well as CD8 T-cells, down-regulates several B-cell functions, including immunoglobulin production, and natural killer (NK)-cell activities 18–24. Furthermore, cytotoxic T-cells are also downregulated in the presence of TGF-β. Resolution of inflammation, particularly during chronic inflammation, results in fibrogenesis and angiogenesis. Both are also under close control of TGF-β. TGF-β expression has been observed in mucosal tissues. The expression pattern in the mucosal tissues of the gut and nose initiated a number of investigations in the field of mucosal tolerance. There is ample evidence that particularly on the mucosal site TGF-β is directly involved in the regulation and maintenance of oral tolerance 25–27.
Formation and maintenance of T-cell effectors do not represent a static and fixed function of the immune system, but rather demonstrate a great extent of dynamic plasticity operating on several regulatory levels. Firstly, at the level of T-cell development, the development of T-cell effector subtypes depends on a number of signals and factors delivered both in a cognate as well as in a non-cognate fashion. Antigen-presenting cells phagocytose and process antigen and present antigenic peptides via major histocompatibility (MHC) class-II molecules. These antigenic peptides are bound to the MHC binding groove in the endoplasmic reticulum followed by transport to the cell surface. This complex of MHC plus peptide engages with the α/β-T-cell receptor. In addition to this cognate signal, several noncognate signals have been identified which contribute to Th-subtype development (fig. 1⇓). For example, the development of Th-1 T-cells is triggered by IL-12 and IFN-α, whereas the development of Th-2 T-cells requires initial IL-4 28. However, cellular sources ofthis initial IL-4 have not been convincingly identified. Mast cells and basophils have been proposed as possible sources 29, 30. Most recently a novel concept has been put forward proposing that the dichotomy of T-cell effectors is also reflected on the level of antigen presenting cells. Most prominently Kaliniski et al. 32 have proposed the development of analogous types of dendritic cells (DC) termed DC-1 and DC-2 which promote the development of Th-1 versus Th-2, respectively 31, 32.
The second level of plasticity is exhibited on the level of pre-existing Th-cell subsets (fig. 1⇓). For example, the Th-1 derived cytokine IFN-γ is the most potent inhibitor of Th-2 immune cells, and vice versa IL-4, IL-10, and to a lesser extent TGF-β, suppress Th-1 T-cell functions. Furthermore, IL-12 and IFN-γ have been shown to inhibit Th-3 effector T-cells. It is of great importance that naive T-cells are not preprogrammed to develop into either subset. However, the microenvironment at the site of antigen presenting cell (APC)/T-cell interaction is decisive for the development of these T-cell effectors. Once a single T-cell has been fated for a certain Th-phenotype, this decision is fixed. A redirection from one phenotype into another T-cell phenotype has not been shown so far on the single cell level.
How is the decision made about the development of T-cell effectors, and what are the factors determining the outcome of the response? So far, individual components of the microenvironment have been identified which play a role in the development of certain T-cell effector phenotypes. They include: 1) type of APC (e.g. B-cells versus dendritic cells); 2) nature of the antigen; 3) dose of the antigen (low dose versus high dose); 4) site of antigen encounter (mucosal versus systemic); and 5) age of initial antigen priming (pre-, postnatal, adult) and others. This concept is of great practical importance for the development of naturally occurring immune responses. It implies that each time an antigen is delivered to T-cells a correct recognition and decision has to be made in terms of whether this is a harmful antigen or a harmless one and, as a consequence, which type of T-cell effector response represents the most appropriate answer.
Concept: atopic infants start out as T-helper 1 weaklings
The concept of Th-1, Th-2 immune deviation also applies to the situation of pregnancy. It has been revealed during the last few years that the foeto-maternal interface represents an immunologically highly active site of regulation. Since foetal tissue is regarded as a classical allo-transplant expressing histocompatibility antigens (HLA) from father and mother, rejection of this allo-transplant must be avoided. The immune system has developed several strategies to prevent recognition of paternal HLA antigen, followed by subsequent development of cytotoxic T-cell responses against these cells. One major strategy to accomplish this relates to the Th-1/Th-2 concept. Since it is the task of both CD4 and CD8 Th-1 T-cells to mediate transplant rejection, development of such dangerous T-cell responses must be suppressed. This is, at least, in part accomplished by the profound upregulation of cytokines belonging to the Th-2 and Th-3 phenotype. In fact, high level expression of IL-4, IL-10 and TGF-β1 has been demonstrated at the foeto-maternal interface 33. Both, foetal and maternal cells contribute to these regulatory events.
At the same time, development of the foetal immune system takes place and, at least, the T- and B-cell compartments are functionally set up around weeks 17 to 20 of pregnancy. These lymphocytes are then ready to encounter antigens and to develop into mature T-cell effectors. Evidence for this active immune maturation process has been provided by several investigators. It is now well accepted that antigen specific T-cells are present in cord blood (CB) 34, 35. These cells are indeed of foetal and rather than maternal origin. T-cell responses against nutritional antigens including casein, lactoglobolin, lactalbumin and ovalbumin as well as T-cell responses against harmless environmental antigens including house dust mite and certain pollen allergens have been demonstrated. Active intra-uterine development of the B-cell compartment is indicated by the presence of antigen-specific secretory IgA antibodies (sIgA) in newborns. Again cow's milk and hen's egg sIgA antibodies have been detected 35.
Such T-cell maturation occurs intra-uterinally in a Th-2 and Th-3 biased environment. This environment apparently has a marked impact on the development of Th-1 and Th-2 phenotypes in late foetal and early post-natal life.
Several studies indicate that the Th-1 capacity as assessed by the amount of IFN-γ production is even further reduced in cases at risk, as compared to non-risk neonates (table 2⇓). The studies which address this issue differ in several major methodological aspects. Warner et al. 36 and Kondo et al. 37 assessed cytokine production in allergen stimulated CB mononuclear cells and followed infants for up to one year and six years, respectively. Outcome parameters were atopic eczema and skin test reactivity. In contrast, Tang et al. 38 and Martinez et al. 39 employed mitogen stimulation of CB-cells and followed their cohort for one and six years, respectively. The cytokine pattern of allergen specific T-cell clones was assessed in the study by Piccinni et al. 40 who reported a follow-up period of three years. Prescott and co-workers 35, 41 studied allergen specific T-cell responses and provided a thorough follow-up for two years. They determined cytokine production on both transcriptional and translational levels. Regardless of whether CB-cells were stimulated with allergen or mitogen, a further reduced capacity to produce IFN-γ in each case has been noted as compared to nonrisk controls. Although the data are quite consistent with respect to suppressed Th-1 immunity, the pattern of Th-2 immunity is still a matter of debate. In most studies Th-2 cytokine production has not been investigated with the exception of the studies by Prescott and co-workers 35, 41. These reports provide initial evidence that reduced IFN-γ production might be associated with enhanced and even persistently elevated Th-2 cytokines production in such infants. However, it is important to note that the size of the cohorts was small and the period of follow-up was rather short. Further longitudinal studies need to be conducted with well defined cohorts in order to accumulate sufficient data allowing prospective immunomodulatory strategies with the aim of readjusting the Th1/Th2 balance in infants at risk.
The first experimental evidence that maternal sensitization favours the development of allergy in offspring was obtained from animal studies (fig. 2⇓). These studies showed that offspring from normal healthy mice were characterized by impaired IFN-γ production at birth which was even further lowered in offspring from allergen-sensitized mothers. After exposure to a heterologous allergen, the offspring derived from allergen-sensitized mice developed an accelerated Th-2-driven immune response compared to offspring from nonsensitized mice 42. These data suggest that Th-2/Th-0 immunity present during pregnancy has a decisive impact on shaping the Th-1/Th-2 T-cell profile in response to post-natal allergen exposure.
A reliable biomarker to assess the presence of Th-1 immunity on the level of antigen-specific responses is the development of delayed type hypersensitivity (DTH) responses. It is a well known phenomenon that, for example, infants with atopic dermatitis express suppressed responses to recall antigens in this test. This test cannot only be performed in vivo, but also in vitro by assessing lympho-proliferative responses to antigens. The useful prototypic antigen in this regard is the proliferative response to tetanus toxoid (TT) following dipheria-pertussis-tetanus (DPT) vaccination. Recently Prescott et al. 43 published an elegant study in which lympho-proliferative responses to TT were assessed in 2-yr-old infants following DPT vaccination. The response pattern was correlated with the level of serum IgE. They found several fold higher levels of serum IgE in those infants exhibiting a lack of lympho-proliferative responses. In contrast, infants with a positive response to TT following DPT vaccination had reduced levels of serum IgE 43. Again, this observation further underscores the inverse relationship between Th-1 immunity (as assessed by the presence of DTH responses) and Th-2 immunity (as assessed by serum IgE antibody levels). However, the question concerning the causal relationship between these two events remains open. Whether the incapacity to mount appropriate Th-1 immune responses allows the uncontrolled development of Th-2 immunity, or whether infants with already elevated Th-2 immunity have an incapacity to mount appropriate DTH responses remains to be studied.
Relationship between infections and development/exacerbation of allergy and asthma
In numerous studies during the last decades, an inverse relationship between asthma and the overall burden of respiratory infections has been reported. These studies are summarized in table 3⇓. Although these studies relate to an interesting concept, the cause-effect relationship regarding the impact of viral infections on the prevention/suppression of allergy and asthma still remains an open issue.
There is no doubt that common colds are the most frequent cause of asthma exacerbation in both children and adults 58–62. The underlying mechanisms have been extensively analysed and reviewed 63. One mode of action might be an additive effect of airway inflammation resulting from the inflammatory response to viral infections in an already inflamed mucosal tissue. Several studies have found an increased incidence of asthma in children with a history of childhood bronchiolitis 51–53, 56, 57, 64–70. Virus infected patients with asthma have enhanced cytokine responses, apparently leading to prolonged lymphocytic and eosinophilic accumulation in the tissue 71. One of the major causes of early childhood bronchiolitis is respiratory syncytial virus infection, which will be extensively discussed by Schwarze and Gelfand 72 in this series.
The data from these studies and further experimental studies (discussed later) suggest that the timing between infection and allergen exposure may be of great importance in terms of the immunological consequences. In the case of a wide gap between infection and allergen exposure, infections may have a protective effect. In contrast, a closely timed relationship between infection and allergen exposure and infection of already allergic/asthmatic patients may have a disease aggravating effect. Although the evidence supporting the concept of asthma exacerbation in relationship to viral infections is relatively strong, the association of bacterial respiratory infections with exacerbations of wheezing remains relatively weak. An exception is the association between chronic sinusitis and asthmatic symptoms. The data linking infection of the para-nasal sinuses to asthma are mainly associative and have been reported by several investigators 73–78. However, causality has been difficult to prove. The mechanisms by which infection of the sinuses may contribute to acute exacerbation of asthma are known, but may include bacterial seeding from the upper to lower respiratory tract (the post-nasal trip theory), naso-sino-bronchial reflex or infection-induced enhancement of β-adrenergic blockade.
The issue of mycobacterial infection and Bacillus Calmette-Guérin (BCG) vaccination
The relationship between mycobacterial infection and Bacillus Calmette-Guérin (BCG) vaccination represents another model situation to assess the relationship between Th-1 and Th-2 immunity. Bacterial infections with, for example, Bordetella pertussis, Listeria monozytogenes, Mycobacterium bovis, leishmania and many others are associated with increased production of IFN-γ, reflecting the requirement of Th-1 immunity for defence and protection 79–82. Cytokines produced by the cells of the innate immune system including macrophages and NK cells in response to Mycobacterium tuberculosis and/or its components, were able to shift, at least in vitro, the development of allergen-specific T-cells from a Th-2/Th-0 to a Th-1 profile. These cytokines include IL-12 and IFN-α which play an important role in the development of Th-1 effector cells.
One of the first studies which addressed this issue, employing a cross-sectional epidemiological approach, was published by Shirakawa et al. 59. The authors reported that among BCG immunized Japanese school children, asthmatic symptoms and several other index factors of atopy, were significantly less likely in positive tuberculin responders compared to negative responders, and that remission of atopic symptoms between the ages of seven and 12 years was much more likely in positive tuberculin responders. The interpretation of these findings has been debated intensively. It is of importance that all infants enrolled in this study have undergone an extensive BCG vaccination schedule. At the age of six years tuberculin nonresponders have been revaccinated. The most likely interpretation of these results is that the inverse association between the atopic status and DTH responses may simply reflect the imbalance of Th-1/Th-2 responsiveness. It is the failure to respond appropriately after BCG vaccination that is associated with the development of atopic symptoms rather than the BCG vaccination per se. Such an interpretation is further supported by the findings of Swedish investigators, again indicating that a single immunization for BCG after birth does not affect the prevalence of atopic diseases at school age 83, 84.
In recent studies, however, an inverse relationship between BCG vaccination and atopy, as defined by skin-test reactivity to common allergens, has been reported in Guinea Bissau 85. However, in that study the BCG response pattern has not been assessed. In contrast, a study performed on unselected children in Sweden found no reduction in atopic diseases associated with BCG in infancy amongst children born in Sweden 84. In contrast, among immigrant children, many born in Asia and South America, BCG was associated with a lower prevalence of atopic diseases 86. This interesting observation may point to differences among ethnic groups.
It is important to distinguish between BCG vaccination and mycobacterial infection. Recently, von Mutius et al. 87 reported a significant inverse correlation between the prevalence of asthma and reported rates of tuberculosis. To further explore underlying mechanisms of this epidemiological phenomenon animal model systems have been employed by several investigators 88–90. In one such model IgE-high responder BALB/c mice were infected with a BCG vaccine prior to sensitization against ovalbumin. It is important to note that the BCG vaccine contains attenuated mycobacteria which results in severe generalized mycobacterial infection in such animals. Therefore, this reflects a model situation of generalized infection with a facultative intracellular organism. It was shown that such an infection prior to allergic sensitization prevented the development of IgE/IgG1 antibody responses, immediate positive-type skin-test responses and airway hyperresponsiveness together with prevention of allergic airway inflammation. In parallel, a marked rise in antiovalbumin specific IgG2a antibodies was found, reflecting the development of an OVA-specific Th-1 instead of Th-2 response 88. A further study revealed that this effect was mediated via IFN-γ since the protective effect of mycobacterial infection was lost in IFN-γ receptor knock-out mice 89. However, the exact mechanism of how IFN-γ mediates this effect is not yet clear. Interestingly, it appears that bacteria do not necessarily have to be alive in order to prevent allergic responses since killed Listeria monozytogenes, Mycobacterium vacae or Lactobacillus plantarum could also suppress allergic responses in mice 80, 91.
Bacterial components: deoxyribonucleic acid
It is of major interest to identify and characterize bacterial components which are able to influence the Th-1/Th-2 balance. In this context, bacterial deoxyribonucleic acid (DNA) has been shown to exert a dramatic effect on the immune system 92. Bacterial (but not mammalian) DNA has mitogenic properties for a number of cells of both the innate and adaptive immune system. These properties are due to unmethylated “CpG motifs” containing oligodeoxynucleotides (ODN) 93, 94. The key CpG motif influencing human immune responses is defined by the GACGTT sequence and in the mouse by GTCGTT. The phosphorothionate backbone seems to be an essential prerequisite of CpG-ODNs to exhibit their immunomodulatory properties.
The immunogenicity of such CpG-ODNs has been extensively studied, particularly in the murine system 95–99. They directly interact with certain subsets of APC including DC subpopulations, NK and B-cells 100. The activation of DC by CpG-ODN results in increased expression levels of costimulatory molecules and an increase in antigen presentation and priming 101, 102. Within a few hours, several chenokines and cytokines are being expressed, especially IL-12, IFN-α and TNF-α, all of them playing an important role in Th-1 promotion. Monocytes and macrophages exhibiting increased antibody-dependent cellular cytotoxicity. IL-12, TNF-α and IFN-α have direct effects on NK cell activity 103. They induce IFN-γ and increase the cytolytic activity of NK cells. To a lesser extend, CpG-ODN also affect B-cell activities 104. B-cells rapidly produce IL-6 and IL-10 and express increased levels of costimulatory molecules 105. Although T-cells are not directly activated by CpG-ODN, their response and activation pattern is indirectly affected via the enhanced response profile on APC. Thus, development of Th-1 effector cells on both the level of CD4 and CD8 T-cell subsets is a major result of exposure to CpG-ODN 106.
The therapeutic potential of bacteria and bacterial extracts in the treatment of particularly malignant disease has been known for more than a hundred years. In this respect, BCG extracts have been studied to a great extent. About sixty years ago, Freund's adjuvant, consisting of mycobacterial extract in oil immersion, was characterized as an immunomodulator with similar therapeutic activities. Defined as complete Freund's adjuvant this is one of the most potent modalities for generation of Th-1 immune responses. Yet, it was not until 1984 that Tokunaga et al. 107 found that it was the mycobacterial genomic-DNA activation of NK cells that was responsible for the potent anti-tumour activity of BCG 107. This observation stimulated further research on the immunogenicity of bacterial DNA 108–111. The potent immunological response suggests that bacterial DNA serves as an evolutionary conserved lesion for the vertebrate immune system which recognizes this structural pattern as a danger signal. More recently, synthetic ODNs containing immunostimulatory DNA sequences have been identified to exhibit adjuvant activity and to induce Th-1 like immune responses.
In addition to their anti-tumour and cytotoxic activity, Th-1 cells play an important role as immunoregulators by direct Th-2 inhibiting effects. Based on this capacity, it has recently been explored whether CpG-ODNs have antiallergic effects. This preventive and/or therapeutic potential has been extensively analysed, particularly using murine models of allergy and asthma 112. It has been shown that only one injection or mucosal administration of CpG-ODNs given before the allergen challenge to sensitized mice inhibited lung and blood eosinophilia better than dexamethasone 113. Levels of IL-3 and IL-5 and airway hyperreactivity were also reduced whereas IFN-γ levels were increased. This phenomenon of immediate inhibition of the allergic response has been recently confirmed in another model of ragweed sensitization 114. In that study it has been shown that the long-term Th-1 diversion of the allergen-specific response is stable for at least six weeks. This therapeutic potential of CpG-ODNs has been further explored in a well established animal model system of major birch pollen allergen which resembled a clinically highly relevant allergen model 115. Coadministration of CpG-ODNs with birch pollen allergen in aluminium hydroxide resulted in markedly increased Th-1 activities, resulting in markedly elevated IgG2a levels and subsequently reduced airway inflammation. Furthermore, systemic or local/mucosal application of CpG-ODNs after aerosol sensitization also induced this Th-1 dependent antibody profile. The authors conclude that such CpG-ODNs as adjuvants may have an important impact on new forms of specific immunotherapy in allergic disease. In this context, particularly the mucosal route of application might have further potential. Although both prophylactic and therapeutic effects have been clearly demonstrated in several animal model systems, further research is needed to analyse possible long-term and side-effects and its efficiency in man.
Bacterial components: lipopolysaccharides
Several cross-sectional and longitudinal epidemiological studies have revealed a reduction in allergen sensitization and atopic diseases in children of farmers, children with pigs, dogs, or cats in their homes, children raised in day-care centres from an early age, and children of large families 116–120.
An interesting “human model” that may provide further insight in this context is the recent observation reported by several authors that growing up on farms confers significant protection against the development of atopy. This has been clearly demonstrated in a Swiss population study of school children aged 6–15 yrs 121. Similarly in a large survey of Bavarian children entering school at ages 5–7 yrs old, the prevalence of hay fever among children raised on a farm was significantly lower than among their peers from the same villages who did not grow up on a farm 122. Furthermore, a recent Austrian survey of children aged 8–10 yrs has confirmed these findings 123. Interestingly, the development of atopic eczema was not effected by the farming activities of the parents.
What is the nature of the environmental factor present on farms which confers protection against allergy and asthma? The living conditions of farming families differ in many respects from the lifestyles of other families and include a larger family size, more pets, frequently heating with wood or coal, less maternal smoking, more dampness and unique dietary habits. In the study cited above none of these factors could so far explain the effect. In contrast, contact with livestock and poultry was found to explain much of the relationship between farming and atopy.
In all these environmental settings gram-negative bacteria and their cell-wall component endotoxin (lipopolysaccharide (LPS)) may exist in abundance. Indeed, occupational disease studies have reported high levels of endotoxin exposure in farm settings 124, 125. Since bacterial endotoxin is known to induce IFN-γ and IL-12 production, the hypothesis has been generated that chronic endotoxin exposure during early childhood and infancy may protect against the development of allergy and asthma.
This hypothesis was the basis for two recently published studies which aimed to compare indoor exposure to endotoxin and house dust with allergen sensitization in infants at high risk for developing asthma and/or wheezing. Gereda et al. 126 measured concentrations of house dust endotoxin and allergens in the homes of 61 infants, aged 9–24 months, with at least three physician documented episodes of wheezing. In parallel, skin-prick testing with a panel of common inhalant and food allergens was carried out and the cytokines reflecting Th-1 and Th-2 activities were measured. The homes of allergen-sensitized infants contained significantly lower concentrations of house dust endotoxin than those of nonsensitized infants. Increased house dust endotoxin concentrations correlated with increased proportions of IFN-γ producing CD4 T-cells.
However, another recent study did not confirm this association. Park et al. 127 examined the associations of house dust endotoxin, measured within the first three months after birth, with wheezing during the first year of life in a cohort of 499 children with one or both parents having a history of asthma or allergy. Their results suggest that endotoxin is associated with an increased risk of wheezing, and may promote persistent wheezing during the first year of life among children with a history of allergy or asthma. This occurred independent of the effect of lower respiratory infections, one of the strongest risk factor for wheezing in infancy. The association between early childhood infections and subsequent development of asthma were elegantly assessed in a birth cohort study by Illi et al. 120. The most important conclusion from this study was that the risk of asthma by age seven is reduced by ∼50% in children with two or more reported episodes of common colds during the first year of life. In contrast, children with wheezy lower respiratory illnesses in the first year of life tend to wheeze later on.
LPS serves as a potent inducer of cytokine-favouring Th-1 T-cell development. One mechanism through which LPS mediates this effect is by binding together with the LPS binding protein to the LPS receptor, CD14, which is expressed on a variety of APC, particularly monocytes and macrophages. Polymorphisms in the 5' linking region of a CD14 gene have recently been identified and were associated with the intensity of atopy 128.
These data from cross-sectional studies provide circumstantial evidence suggesting a protective role for LPS in the development of allergic disease. However, the underlying molecular and cellular mechanisms are still unknown and need to be assessed.
It needs to be emphasized that the immunological consequences of LPS-exposure depend on a variety of conditions. It would be an oversimplification to conclude that in all cases LPS-exposure protects against allergy and asthma. The source and route of exposure to LPS may also be of importance since LPS is produced by several bacterial pathogenes (e.g. Haemophilus and Salmonella spp.) and comensales colonizing the gut (e.g. Escherischia coli). In this context it has been reported that the colonization of the gastrointestinal tract from newborn babies who had Lactobacillus and Eubacteria spp. (compared with those who had Clustridium difficile) correlated with a decrease in atopic disorders later in life 129. Furthermore, it is well known that long-term high-dose exposure to LPS triggers a variety of adaptation processes which result in macrophage “desensitization”.
Animal model systems offer the opportunity to assess the immunobiological effects of LPS on allergy and asthma. In this regard, it is now clear that the sequence of events determines the outcome. When LPS exposure occurred before allergen sensitization/challenge, a preventive effect of LPS has been documented. In this regard intravenous LPS reduced antigen-induced bronchoconstriction in the guinea pig, and exposure early in sensitization prevented allergic sensitization in rats 130, 131.
In contrast, when LPS exposure occurred in already allergen-sensitized animals, LPS promoted the inflammatory response (fig. 3⇓). This has been shown in several studies including a more recent investigation indicating that exposure to LPS in sensitized rats exacerbated the allergic immune response 131, 132. To further complicate this issue, it has been also documented that intratracheal LPS induced bronchial hyperresponsiveness to serotonin and acetylcholine in the guinea pig even in the absence of allergen exposure and/or sensitization.
In conclusion, there is now circumstantial evidence suggesting that continuous LPS exposure in early life may have a preventive effect on allergy and asthma. This situation is closely reflected by the results in animal model system where LPS exposure prior to allergen sensitization has a well-documented preventive effect. In contrast, LPS exposure at a later stage, when allergen sensitization has already been established, may exacerbate and promote the inflammatory response. These experimental data may explain the clinical observation of asthma exacerbation during bacterial and viral infection. Furthermore, these biological effects also depend on the route of exposure since intestinal LPS exposure may have a different effect.
Bacterial components: superantigens
Just as gram-negative bacteria secrete LPS as a major immunomodulatory component, gram-positive bacteria provide, via secretion of bacterial superantigens (enterotoxins), a major immunomodulatory stimulus (fig. 4⇓). Bacterial superantigens act via a set of biological properties differing distinctly from conventional antigens. In contrast to conventional antigens which require uptake by APC, intracellular processing, presentation on MHC-class II molecules and recognition of the MHC-class II plus peptide complex by the T-cell receptor, bacterial superantigens function differentially. They bind outside the conventional T-cell epitope-binding-groove on MHC-class II molecules and crosslink the MHC-class II molecule with the T-cell receptor Vβ chain. In the case of a conventional antigen, this mode of APC cell interaction results in the stimulation of <0.1% of naive T-cells. In contrast, bacterial superantigens which recognize a defined set of T-cell receptor Vβ elements, stimulate up to 25% of naive T-cells. Such interaction results in vigorous T-cell activation and cytokine release, usually followed by deletion and apoptosis of the majority of T-cells.
One major source of such bacterial superantigens is S. aureus. In the case of atopic dermatitis, chronic colonization of the skin with toxin-producing S. aureus strains has already been well established. In a recent cross-sectional study ∼80% of children with atopic dermatitis (AD) show chronic colonization of the skin with S. aureus and >50% of these children were colonized with toxin-positive strains 134. It was then assessed whether colonization with toxin-positive or toxin-negative strain would have any correlation with clinical severity as assessed by SCORAD (severity scoring of AD). The results of this study revealed the most severe cases of atopic dermatitis in the patient group colonized with toxin producing S. aureus strains. These data resulted in the hypothesis that bacterial toxins have a major impact on clinical severity of atopic dermatitis.
To further explore this issue, the T-cell receptor Vβ repertoire of skin-infiltrating T-cells has been correlated with the pattern of toxins produced by the individual S. aureus strains 135. Preferential accumulation of superantigen-responsive T-cells was observed in patients with intense skin inflammation. Additional evidence for disease-modulating effect of superantigens has been provided by a study performed by Strickland et al. 136. The authors showed that, in vitro, superantigens have the capacity to activate and expand T-cells expressing specific T-cell receptor Vβ gene-segments and also to increase their skin-homing capacity via upregulation of this skin-homing receptor, cutaneous lymphocyte-associated antigen in vitro.
The disease-modulating effects of superantigens have also been extensively studied in the model of humanized severe combined immunodeficiency (SCID) mice 137. SCID mice exhibit a severe immunodeficiency syndrome due to a defect in the antigen-receptor recombinase. Due to this deficiency, mature T- and B-cells are absent in these mice and they accept xenotransplants without rejection. Transfer of atopic peripheral blood mononuclear cells from house-dust mite sensitized patients results in the development of house-dust mite specific T- and B-cell responses following antigen booster in these mice. When such reconstituted SCID mice were epicutaneously challenged either with the superantigen staphylococcal enterotoxin B (SEB) or house-dust mite allergen or a combination of both, an inflammatory response developed in the dermis and epidermis similar to skin lesions of atopic dermatitis. Most severe lesions were observed when mice were treated with a combination of SEB and house-dust mite allergen. In a recent study, SEB has been applied on normal and atopic skin, resulting in an influx of T-cells into the skin, leading to the induction of atopic dermatitis-like skin lesions 138.
At least in atopic dermatitis, S. aureus-derived superantigens can also function in an alternative fashion. Specific IgE antibodies directed against staphylococcal enterotoxin A (SEA) or SEB were detectable in a subgroup of atopic dermatitis patients 134, 139. The presence of these superantigen-specific IgE antibodies strongly correlates with disease severity and levels of total IgE. These results lead to the conclusion that these proteins can also function as conventional allergens resulting in an allergen-specific T- and B-cell response. In patients chronically colonized with superantigens secreting S. aureus strains, these toxins can easily penetrate through the disturbed epidermal barrier resulting in direct binding to IgE antibodies which are present on the surface of dermal mast cells. Through this mechanism, mast cells are chronically activated to release pro-inflammatory mediators perpetuating the itch-scratch cycle in these patients.
On the other hand, it has also been demonstrated that such bacterial superantigens can prevent and inhibit allergic responses in the mouse. When mice are cutaneously treated with SEB prior to allergen challenges (ovalbumin), the ovalbumin-induced allergic response is prevented. Although in this model, superantigen treatment most likely resulted in marked activation of the appropriate T-cells followed by deletion of the cells, the underlying immunological mechanisms explaining how superantigens prevent the development of the allergic response have not been explored in depth.
Conclusion
It is well accepted that the inflammatory response in allergic disease is characterized by the presence of a Th-2 mediated response. IgE antibodies represent important effector molecules and eosinophils and mast cells are predominant effector cells of the immediate hypersensitivity response. In (genetically) susceptible individuals with a unique type of immune response a pattern is initiated following the allergen exposure. Since indoor and outdoor allergens are ubiquitous, once the immune response is shifted towards the Th-2 phenotype, a chronic process of immunoderegulation results. Apparently, this process can already start in the intra-uterine environment. Since many, but not all, microbial antigens trigger a Th-1 immune response as a normal defence mechanism, a hypothesis has been put forward proposing that a negative correlation exists between the pattern and frequency of microbial infections and the onset and severity of allergy and asthma.
This hypothesis is supported by epidemiological data, indicating an inverse association between frequency, pattern and type of infection and the prevalence of allergy and asthma. In patients, however, a direct cause-effect relationship has not been demonstrated so far.
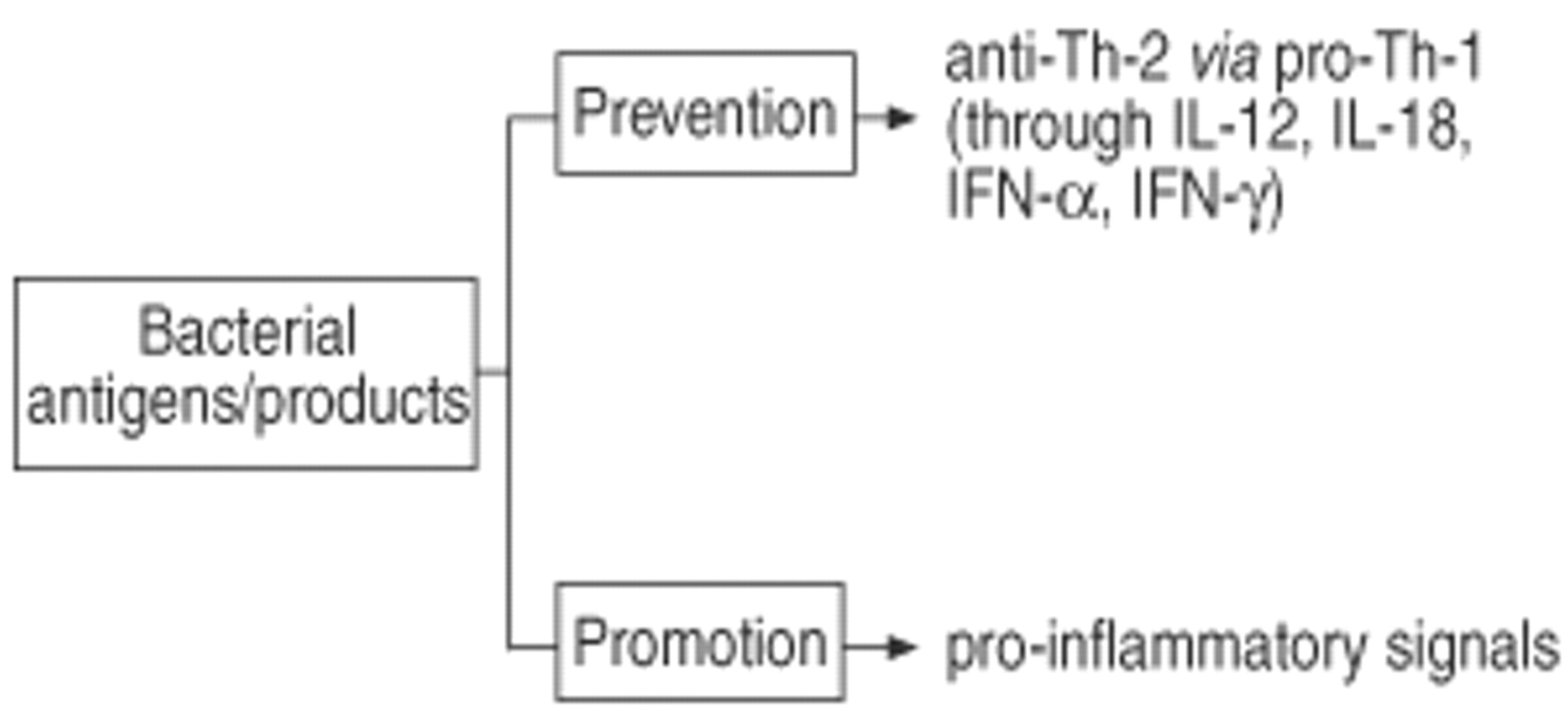
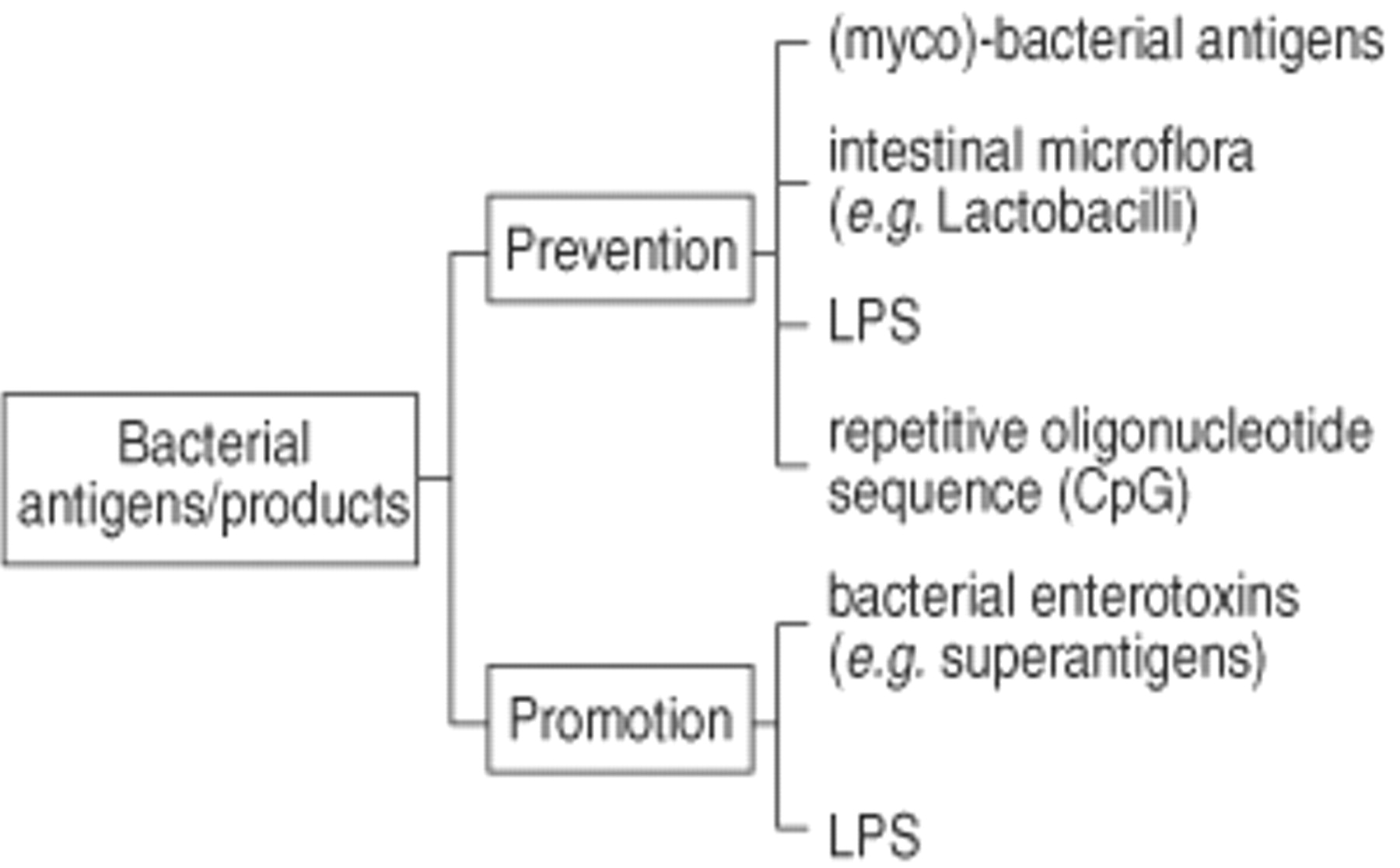
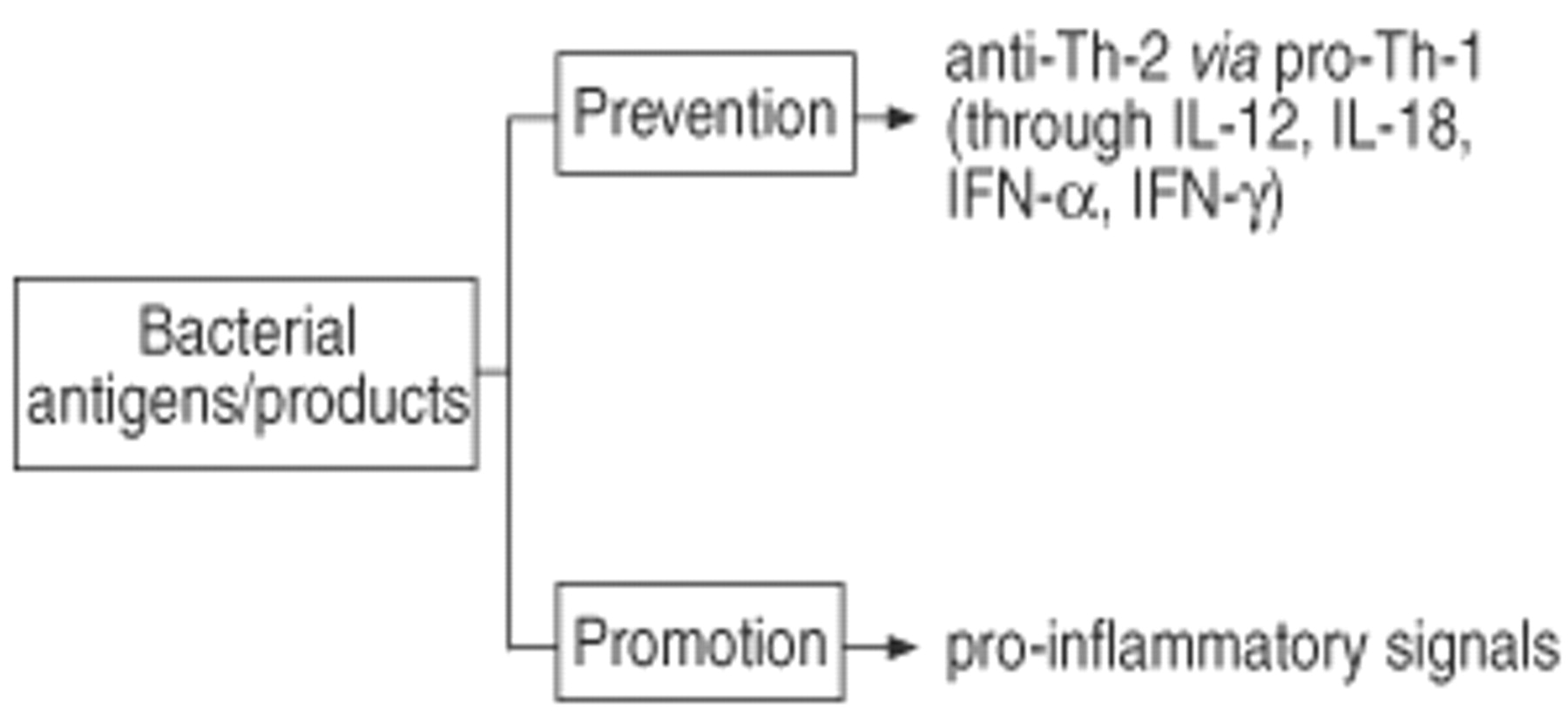
Experimental in vitro systems and animal models have been employed to further analyse the impact of microbial antigens on the development of allergic immune responses. The analysis of the available literature on this issue allows the following conclusion. Firstly, particular bacterial antigens have a capacity to dramatically modulate the immune response pattern (fig. 5⇓). This has been demonstrated in models of bacterial infections (e.g. mycobacterial infections and BCG) as well as in situations were bacterial components have been used (e.g. LPS, bacterial superantigens). Secondly, the same bacterial antigen can either promote or prevent the allergic immune response (fig. 6⇓). The outcome is critically dependent on the timing between exposure to microbial antigens and allergen sensitization. When microbial antigens are given the chance to develop a Th-1 immune response in a naive individual before the allergen can mount a Th-2 mediated inflammatory response, the Th-1 immune response is prevented. In contrast, acute or chronic encounter of bacterial antigens in an already Th-2 inflammed tissue usually aggravates the allergic condition. Thirdly, from current knowledge, a single transient infection or vaccination is not sufficient to direct the immune response towards such a strong Th-1 pattern that this could prevent the development of allergy and asthma later in life. In contrast, only frequent and repeated exposure to a variety of microbial antigens may lead to a long-lasting induction of Th-1 immunity which may, as a consequence, eventually prevent the development of Th-2 immunity and, therefore, may reduce the risk for the development of allergy and asthma.
This concept which is mainly based on experimental data, now requires testing in humans by the means of cohort and intervention studies. It needs to be explored how the immune system can be promoted to develop strong T-helper 1 immunity early in life. The experimental data further suggest that such an approach may only be useful if T-helper 1 immunity is installed before the immune system has the chance to mount a pathological T-helper 2 immune response against allergens in predisposed and susceptible individuals.

Positive and negative regulatory effects of cytokines in development and effector functions of T-helper (Th)-1, Th-2, and Th-3 T-cells. Positive effects: solid arrows; Negative effects: dashed arrows. APC: antigen presenting cell; IL: interleukin; IFN: interferon; TGF: transforming growth factor; TNF: tumour necrosis factor.

Maternal T-helper (Th)-2 immunity affects pre- and postnatal immune responses in an antigen-dependent and independent fashion. Th: T-helper cell; BLG: beta-lactoglobulin; OVA: ovalbumin; IL-4: interleukin-4; IFN-γ: interferon-gamma; TOVA: ova-specific T-cell; TIL-4: IL-4 producing T-cell; TIFN-γ: IFN-gamma producing T-cell. Arrows indicate increase or decrease of production.

Modifications of allergy and asthma by lipopolysaccharides (LPS). BALF: bronchoalveolar lavage fluid.

Bacterial toxins and their putative role in allergy and asthma. LPS: lipopolysaccharide; APC: antigen presenting cell; Th: T-helper cell; TCR: T-cell receptor.

Bidirectional effect of bacterial antigens and their products in allergy and asthma. LPS: lipopolysaccharide.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Prevention of allergy and asthma by microbial antigens; sequence of events and proposed mode of action. Th: T-helper cell; IL: interleukin; IFN: interferon.
Transforming growth factor-β and immune responses
T-helper (Th)-1/Th-2 imbalance at birth and development of atopy
Inverse relationship between viral infections/immunizations and respiratory allergy : clinical and epidemiological data
Footnotes
-
Previous articles in this series: No. 1: von Mutius E. Infection: friend or foe in the development of atopy and asthma? The epidemiological evidence. Eur Respir J 2001; 18: 872–881. No. 2: Wennergren G, Kristjánsson S. Relationship between respiratory syncytial virus bronchiolitis and future obstructive airway diseases. Eur Respir J 2001; 18: 1044–1058.
- Received March 20, 2001.
- Accepted June 19, 2001.
- © ERS Journals Ltd