





Abstract
Background and aims Neutrophilic inflammation is a hallmark of some specific asthma phenotypes; its aetiology is not yet fully understood. House dust mite (HDM) is the most common factor in the pathogenesis of airway inflammation. This study aims to elucidate the role of cross-antibodies against HDM-derived factors in the development of neutrophilic inflammation in the airway.
Methods Blood samples were collected from asthma patients with chronic neutrophilic asthma for analysis of HDM-specific cross-reactive antibodies. The role of an antibody against HDM-derived enolase (EnoAb) in the impairment of airway epithelial barrier function and induction of airway inflammation was assessed in a cell culture model and an animal model.
Results High similarity (72%) of the enolase gene sequences was identified between HDM and human. Serum EnoAb was detected in patients with chronic neutrophilic asthma. The EnoAb bound to airway epithelial cells to form complexes with enolase, which activated complement, impaired airway epithelial barrier functions and induced neutrophilic inflammation in the airway tissues.
Conclusions HDM-derived enolase can induce specific cross-antibodies in humans, which induce neutrophilic inflammation in the airway.
Abstract
House dust mite (HDM)-derived enolase induces cross-antibodies in the body, which recognise both HDM-derived enolase and the enolase in the airway epithelial cells, activate complement and induce neutrophilic inflammation in the airway tissues https://bit.ly/2XKSCgy
Introduction
Airway chronic inflammation generally features as profound inflammatory cell infiltration in the airway mucosa [1]. The inflammatory activities result in narrow airway calibration and patients experience difficulty in breathing [2]. The causative factors of chronic airway inflammation are various. Allergic responses are a major cause, in which house dust mite (HDM)-derived allergens are the canonical factors contributing to the pathogenesis of allergic responses in the airway mucosa [3]. Although research in airway allergic inflammation has advanced rapidly in the past a few decades, the initiating factors of airway chronic inflammation and the underlying mechanism remain to be further investigated [4]. The therapeutic effects of airway inflammation are not satisfactorily understood [5]. Thus, it is necessary to further investigate the mechanism, and invent novel and effective therapies for chronic airway inflammation.
The airway epithelium is an interface between the body and external environment. Some allergens, such as those derived from HDM, can be absorbed by epithelial cells. Although there is an enzyme system in epithelial cells that can degrade the absorbed protein antigens into small peptides or amino acids, some peptides with competent antigenicity may be transported to the subepithelial regions where the antigens contact immune cells and initiate immune responses [6]. The immune cells thereby produce antigen-specific immunoglobulin (Ig)E or/and antigen-specific IgG. IgE binds to the high-affinity receptors on mast cells, sensitising them. Re-exposure to a specific antigen activates sensitised mast cells. The mast cells release chemical mediators to evoke allergic responses [7]. The specific IgG can bind specific antigens to form complexes. The complexes may be cleaned by phagocytes, or activate the complement system to chemoattract neutrophils and induce inflammation in local tissues [8, 9]. Yet, whether such complexes are associated with the pathogenesis of airway chronic inflammation has not been defined.
It is recognised that the gene sequences of some extrinsic proteins, such as airborne allergens may be the same as that of some proteins in the human body. After getting into the body, the immune system may produce antibodies against the extrinsic proteins [10]. Apart from recognising the specific extrinsic proteins, the antibodies may also recognise proteins in the human body with similar gene sequences to the extrinsic proteins [11]. These antibodies are designated “cross-antibodies”. Homologous antibodies have cross-reactivity; for example, those against streptococcal-vimentin induce a microvascular cardiac endothelial proinflammatory phenotype in rheumatic heart disease [12]. The cross-antibodies may form complexes with specific constitutive homologous antigens. The complexes activate the complement system, chemoattract inflammatory cells and induce inflammation in the local tissues [13]. Previous studies indicate that enolase-specific antibodies are associated with the pathogenesis of mucosal inflammation, e.g. inflammatory bowel disease [14]. Enolase is a metalloenzyme involved in glycolysis and exists in all tissues and all organisms [15]. Our previous study found that the HDM-derived enolase gene had high similarity to human enolase [16]. Whether the enolase cross-antibodies are involved in the pathogenesis of airway chronic inflammation remains to be further investigated.
Neutrophilic asthma is a subtype of asthma, in which many neutrophils gather in the lung tissue as well as appear in the airway tract [17]. These neutrophils release inflammatory mediators to induce inflammation in the local tissues [17]. The causative factors of neutrophilic asthma remain largely unknown and need to be further investigated. As IgG autoantibody-induced cytotoxicity against airway epithelial cells plays a role in the pathogenesis of nonallergic asthma [18], neutrophilia is often accompanied with IgG-mediated immune inflammation [19] and HDM is the most common factor involved in the pathogenesis of human airway chronic inflammation [3], we hypothesise that HDM-derived factors may be associated with the pathogenesis of neutrophilic inflammation in the airway through inducing IgG-mediated immune responses. Therefore, in this study, the serum HDM-specific IgG levels in neutrophilic asthma patients were assessed, an antibody against HDM enolase (EnoAb) was generated and the role of EnoAb in the induction of airway neutrophilic inflammation was investigated.
Materials and methods
Collection of blood samples from human subjects
Neutrophilic asthma and eosinophilic asthma patients were recruited into the present study at the Department of Respirology, First Affiliated Hospital of Guangzhou Medical University (Guangzhou, China). The diagnosis of neutrophilic asthma and eosinophilic asthma referred to the literature [20], in which neutrophilic asthma patients had <3% eosinophils and >75% neutrophils in the sputum, while eosinophilic asthma patients had ≥3% eosinophils in the sputum. The diagnosis and management of asthma were carried out by our physicians following our routine procedures. Patients with any of the following conditions were excluded: cancer, other autoimmune diseases, allergic diseases, severe organ diseases or those under treatment with immune suppressors for any reason. Healthy control subjects were recruited. Blood samples were collected from each human subject through ulnar vein puncture. Sera were isolated from blood samples by centrifugation at 4°C and stored at −80°C until use. The demographic data of human subjects are presented in table 1.
Demographic and clinical characteristics of recruited subjects
Anti-Dermatophagoides farinae EnoAb recognises airway epithelial cells
16HBE cells (a human airway epithelial cell line) were fixed with 1% paraformaldehyde for 1 h, exposed to anti- Dermatophagoides farinae (Der f) EnoAb or isotype IgG in an Eppendorf tube for 1 h at room temperature. After washing with PBS three times, cells were exposed to the second antibody (labelled with fluorescein isothiocyanate) for 30 min and washed with PBS three times. The cells were then smeared onto a slide, mounted with cover slips and observed using a confocal microscope.
Statistics
Data were tested for normal distribution using the Kolmogorov–Smirnov test. The difference between two groups was determined by t-test or rank sum test. ANOVA followed by Dunnett's test or Bonferroni test was performed for multiple comparisons. The Spearman correlation assay was performed to show the correlation between data of the two groups. p<0.05 was set as a significant criterion. Some experimental procedures are presented in the supplementary materials.
Results
Serum HDM-specific IgG is correlated with neutrophil numbers in the sputum of patients with neutrophilic asthma
To determine the association between HDM-derived factors and the pathogenesis of neutrophilic asthma, we collected peripheral blood samples and sputum samples from neutrophilic asthma patients, eosinophilic asthma patients and healthy control subjects. Serum HDM-specific IgG levels (sIgG, in short), including total sIgG, sIgG1, sIgG2 and sIgG3 were higher in the asthma group than the healthy control group (figure 1a–d). Serum HDM sIgE levels in the neutrophilic asthma group were only slightly higher than in the healthy control group (figure 1e). In addition, significantly more enolase-specific IgG was detectable in the serum in the neutrophil group than that in the healthy control group or the eosinophil group (figure 1f). Frequency of neutrophils in the sputum was also higher in the asthma group than that in the healthy control group (table 1). Positive correlation was identified between sputum neutrophil frequency and serum sIgG levels (including the total sIgG, sIgG1, sIgG2 and sIgG3) in the neutrophilic asthma patients, but not in the eosinophilic asthma patients (figure 1g–j). No apparent correlation was found between serum sIgE levels and sputum neutrophil frequency (figure 1k). In addition, we found that sputum eosinophil frequency did not show correlation with or between serum IgG (supplementary figure S1a–d). A positive correlation was identified between serum IgE and sputum eosinophil frequency (figure S1e). The enolase-specific IgE was detectable in the serum, but there was no significant difference between groups (figure S1f). Serum HDM-specific IgG4 levels were higher in the eosinophil group and the neutrophil group as compared with the healthy control group (figure S1g), which did not have correlation with either sputum eosinophil counts or neutrophil counts (figure S1h and i). The results demonstrate that the neutrophilic asthma patients have HDM sIgG3 in the serum which positively correlates with the frequency of sputum neutrophilic frequency.

Sputum neutrophil number is positively correlated with serum house dust mite (HDM)-specific immunoglobulin (Ig)G levels in neutrophilic asthma patients. Samples of sputum and blood were collected from 41 neutrophilic asthma (NA) patients, 42 eosinophilic asthma (EA) patients and 30 healthy control (HC) subjects. a–f) Scatter plots show a–d) serum IgG, e) IgE and f) enolase-specific IgG levels. g–k) Scatter dot plots show g–j) correlation between serum IgG and sputum neutrophil number or k) serum IgE and sputum neutrophil number. Neutrophil data are presented in table 1. Data are presented as median (interquartile range). A positive part was defined as a level 3 standard error above the mean of HC subjects (the area above the shaded region). Statistics: a–f) ANOVA and Kruskal–Wallis and g–k) Spearman correlation assay. In panel f) recombinant Der f enolase was used in the ELISA experiment to detect the enolase-specific IgG.
A cross-antibody against HDM-derived enolase recognises proteins of airway epithelial cells
We next took a further insight into the role of HDM-derived factors in the pathogenesis of neutrophilic asthma. By whole-genome screening, we identified 16 376 genes in HDM in a previous study [16], in which 2.42–8.25% of the Der f genes had identical gene sequences to human genes varying from 50% to 100% (supplementary figure S2). By comparing the enolase gene sequences between Der f and human, we found that the enolase genes have 72% sequence identity (counting the blues) between Der f and human or 71% between Der f and mouse (supplementary figure S3). The similarity of the enolase gene sequences between Der f and human/mouse prompted us to search the probability that cross-antibodies produced by the human immune system originally against Der f-derived enolase might attack the homologous enolase protein in the human airway tissues. Through the pull-down approach with proteins extracted from 16HBE cells as baits, related proteins were precipitated and analysed by mass spectrometry (MS). The MS results showed that 89 proteins (including enolase) were captured by the IgG isolated from the sera of asthma patients sensitised to mite (supplementary figure S4). Previous reports also indicate that enolase is involved in the pathogenesis of severe asthma [21]. The 16HEB cell-derived enolase could be recognised by anti-Der f antibodies; this was verified by competitive ELISA, in which Der f enolase interfered with the binding between the antibody and human enolase and vice versa (supplementary figure S5). The serum enolase-specific IgG was higher in the neutrophil group than that in eosinophil group (figure 1f). We then assessed the expression of enolase in airway epithelial cells. We found the expression of enolase in both human and mouse airway epithelial cells (supplementary figure S6). Polyclonal antibodies against Der f-derived enolase were generated and designated as EnoAb. The EnoAb could specifically bind recombinant enolases (supplementary figure S5).
EnoAb binds airway epithelial cells to form a complex with enolase
The data in figure 1 implicate that EnoAb may be able to bind to the constitutive enolase in body cells. Thus, mice were treated with EnoAb by nasal drops for 3 days. The airway tissues were observed using immunohistochemistry. As expected, EnoAb specifically bound to the airway epithelial cells of mice (figure 2a). The results were reproduced in an in vitro experiment. By incubating 16HBE cells with EnoAb for 1 h, we found that EnoAb also bound to 16HBE cells (figure 2b). A complex of EnoAb and enolase was detected in the airway tissue protein extracts (figure 2c) and the 16HBE cells (figure 2d). The results demonstrate that EnoAb can bind to the constitutive enolase to form complexes in airway epithelial cells.

EnoAb (an antibody against house dust mite-derived enolase) binds airway epithelial cells to form a complex. A polyclonal antibody (EnoAb) against Dermatophagoides farinae enolase was generated. a) Naïve mice were treated with nasal drops containing EnoAb daily for 3 days. The representative immunohistochemistry images show that EnoAb (in green) binds to the surface of airway epithelial cells (original magnification ×400). b) 16HBE cells (a human airway epithelial cell line) were fixed with paraformaldehyde first, then incubated with EnoAb for 1 h. The representative immunocytochemistry images show that EnoAb (in green) binds to 16HBE cells (original magnification ×400). c) The immunoblots show an immune complex of EnoAb and enolase in mouse airway tissue. d) The immunoblots show an immune complex of EnoAb and enolase in 16HBE cells. The data represent three independent experiments. Scale bars=100 μm. Ig: immunoglobulin.
EnoAb activates the complement system in airway epithelial cells
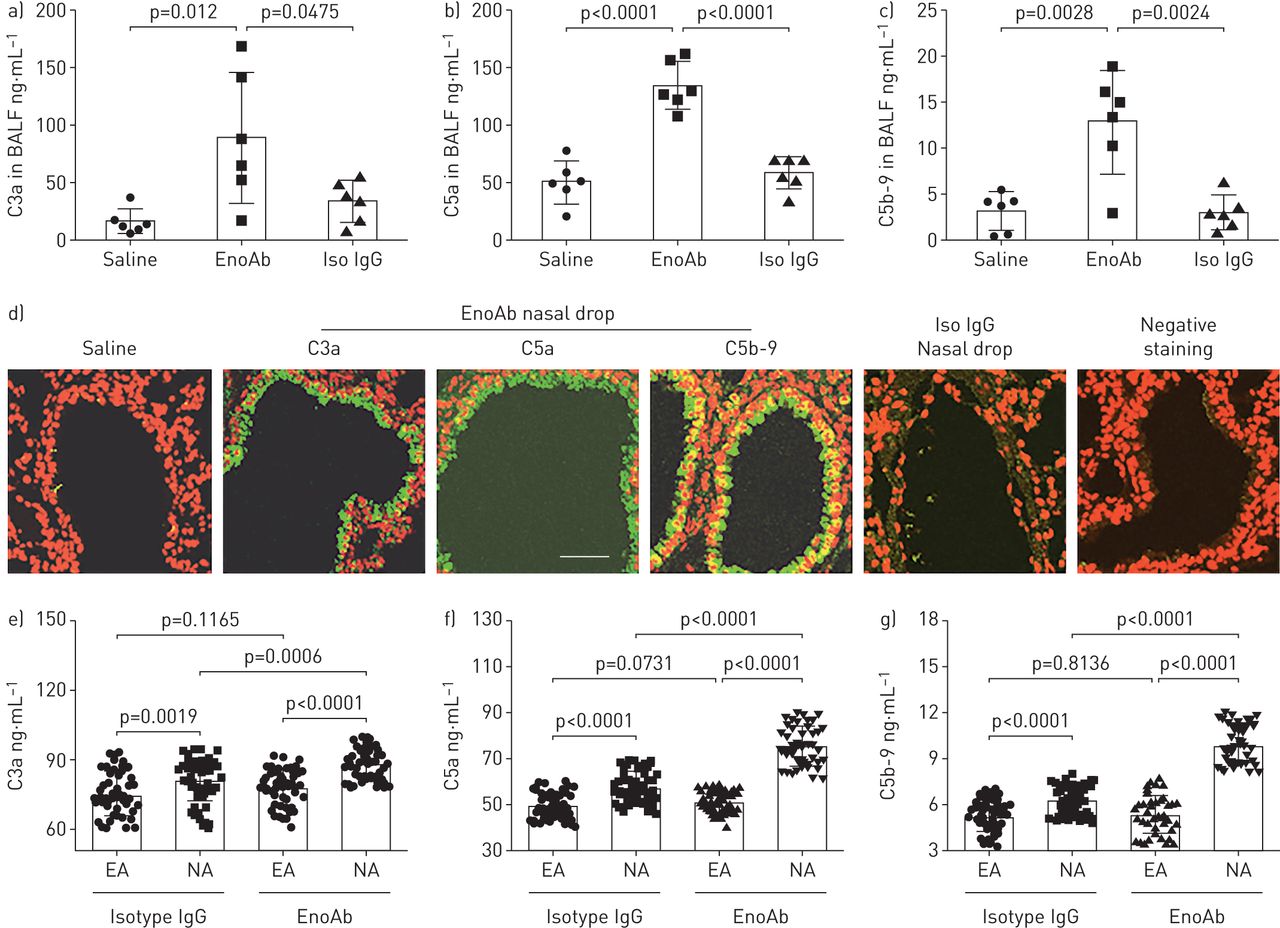
Complement can be produced by several cells, including epithelial cells [22]. The antigen–antibody response can activate the complement system [13]. We tested the effects of EnoAb interacting with enolase on the activation of complement in airway epithelial cells. Naïve BALB/c mice were treated with nasal drops containing EnoAb daily for 3 days. Bronchoalveolar lavage fluid (BALF) was collected; complement levels in BALF were analysed by ELISA. The results showed that the levels of C3a, C5a and C5b-9 were significantly higher in samples from mice that received EnoAb than those that received saline or isotype IgG (figure 3a–c). The results were verified by the immunohistochemistry data, in which the positive staining of C3a, C5a and C5b-9 was localised in airway epithelial cells of the lung tissues (figure 3d). The data indicate that EnoAb can form complexes with enolase and then activate complement on airway epithelial cells. The results were verified with human samples. We collected blood samples from patients with eosinophilic asthma and neutrophilic asthma. The plasma was isolated and incubated with either anti-human EnoAb or isotype IgG. The results showed that exposure to EnoAb markedly increased the levels of C3a, C5a and C5b-9 in the plasma in neutrophilic asthma samples, but not in eosinophilic asthma samples (figure 3e–g). Since activation of complement can promote inflammation [23], the data suggest that EnoAb may induce or exacerbate inflammation in the airway.

EnoAb (an antibody against house dust mite-derived enolase) induces complement activation in airway epithelial cells. Naïve BALB/c mice (n=6 per group) were treated with nasal drops containing EnoAb or isotype immunoglobulin (Ig)G daily for 3 days. a-c) Bronchoalveolar lavage fluid (BALF) was prepared from each mouse and analysed by ELISA. Bar graphs show the levels of a) C3a, b) C5a and c) C5b-9 in BALF. Lung tissue was examined using immunohistochemistry. d) Representative confocal images show the positive staining (in green) of C3a, C5a and C5b-9 in epithelial layers of the lung tissue. The treatment is denoted above each image. The original magnification was ×400. e–g) Blood samples were collected from 42 patients with eosinophil type asthma (EA) and 41 patients with neutrophil type asthma (NA). The plasma was isolated from the samples, incubated with anti-human EnoAb (1 µg·mL−1) or isotype immunoglobulin (Ig)G for 30 min at 37°C and analysed by ELISA. Bar graphs show the levels of e) C3a, f) C5a and g) C5b-9. Statistical method: ANOVA followed by a–c) Dunnett's test or e–f) Mann–Whitney test. Scale bars=100 μm.
EnoAb induces complement-dependent airway epithelial cell apoptosis
The data presented in figures 2 and 3 imply that EnoAb may damage airway epithelial cells. To test the inference, 16HBE cells were exposed to EnoAb in the culture in the presence of human serum (containing complement; supplementary figure S7) for 6 h. The cells were harvested and analysed using flow cytometry. The results showed that exposure to EnoAb slightly induced 16HBE cell apoptosis; the presence of complement significantly enhanced the number of apoptotic cells, which was abolished by heat-quenching complement in the serum. Exposure to EnoAb and recombinant C3a also induced cell apoptosis (figure 4a and b). Since dead cells were not detected in the control group (figure 4a and b), apoptosis is assumed to be the causative factor inducing cell death. Mice were treated with EnoAb by nasal drops daily for 6 days. The airway tissue was processed by TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labelling). The results showed that exposure to EnoAb induced airway epithelial cell apoptosis (figure 4c). The results demonstrate that exposure to EnoAb can induce airway epithelial cell apoptosis, which can be enhanced by the presence of complements.

Airway epithelial cell apoptosis after exposure to EnoAb (an antibody against house dust mite-derived enolase) and serum. a) 16HBE cells were exposed to EnoAb or/and serum (containing complement; collected from healthy control subjects) in culture overnight. The cells were harvested the following day and analysed by flow cytometry. The gated cells are apoptotic cells. b) The bars present the summarised data of apoptotic cells in panel a). c) Naïve BALB/c mice (n=6 per group) were treated with EnoAb or saline daily for 3 days. The lung tissues were examined by TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labelling) staining. The apoptotic cells were stained in green (original magnification ×400). Data are presented as mean±sem. *: p<0.01, compared with group I (ANOVA followed by Dunnett's test). rC3a: Recombinant C3a (100 ng·mL−1; referred to as C3a in the human BALF as presented in figure 3a). Scale bars=100 μm. #: heated serum; the complements were quenched.
Exposure to EnoAb impairs airway epithelial barrier integrity
To test the effects of EnoAb on the airway epithelial barrier integrity, 16HBE monolayers were prepared and exposed to EnoAb or/and complement-containing serum in the culture. The transepithelial electric resistance (TEER) and permeability to macromolecular tracers were assessed and used as indicators of epithelial barrier integrity. The results showed that exposure to EnoAb and complement-containing serum markedly impaired the epithelial monolayer integrity manifesting decrease in TEER (figure 5a). The results were verified using the calcium switch technique (figure 5b). In addition, we found that exposure to EnoAb and complement-containing serum significantly increased the 16HBE epithelial monolayer barrier permeability (figure 5c). It is noteworthy that the presence of heated serum (in which the compliments were quenched) markedly decreased the effects of EnoAb on impairing the epithelial barrier functions (figure 5a–c). After treating mice with nasal drops (containing EnoAb) daily for 6 days, mouse airway epithelial barrier permeability was markedly impaired, which could be blocked by quenching complements with cobra venom factor (CVF) (figure 5d). To elucidate whether exposure to EnoAb/complement alter the status of tight junction proteins in airway epithelial cells, 16HBE monolayers were collected after exposure to EnoAb/serum overnight and analysed using reverse transcriptase quantitative PCR and Western blotting. The results showed that exposure to EnoAb/complement did not apparently alter the levels of ZO-1 in 16HBE cells (supplementary figure S8). The results indicate that EnoAb together with complement can induce airway epithelial cell apoptosis which compromises the integrity of the epithelial barrier.

EnoAb (an antibody against house dust mite-derived enolase) induces airway epithelial barrier disintegration. a and b) 16HBE cell monolayers were prepared. After reaching confluence, the monolayers were treated with the procedures as denoted in the figures. The curves show the transepithelial electric resistance (TEER) at time points as denoted on the x-axis. c) The permeability of 16HBE monolayers was assessed in transwells. The curves show the permeability of the 16HBE epithelial monolayer barrier. d) Naïve BALB/c mice (n=6 per group) were treated with nasal drops containing EnoAb daily for 6 days. The permeability of the airway epithelial barrier was assessed by dextran-absorption. The bars show the permeability of the mouse airway epithelial barrier. The data represent three independent experiments and are presented as mean±sem. The data were analysed by ANOVA followed by Bonferroni test. Ig: immunoglobulin; D: day; FITC: fluorescein isothiocyanate; CVF: cobra venom factor (200 U·kg−1 intraperitoneally) 3 days prior to the beginning of nasal drops to deplete complement in mice. #: heated serum; the complements were quenched.
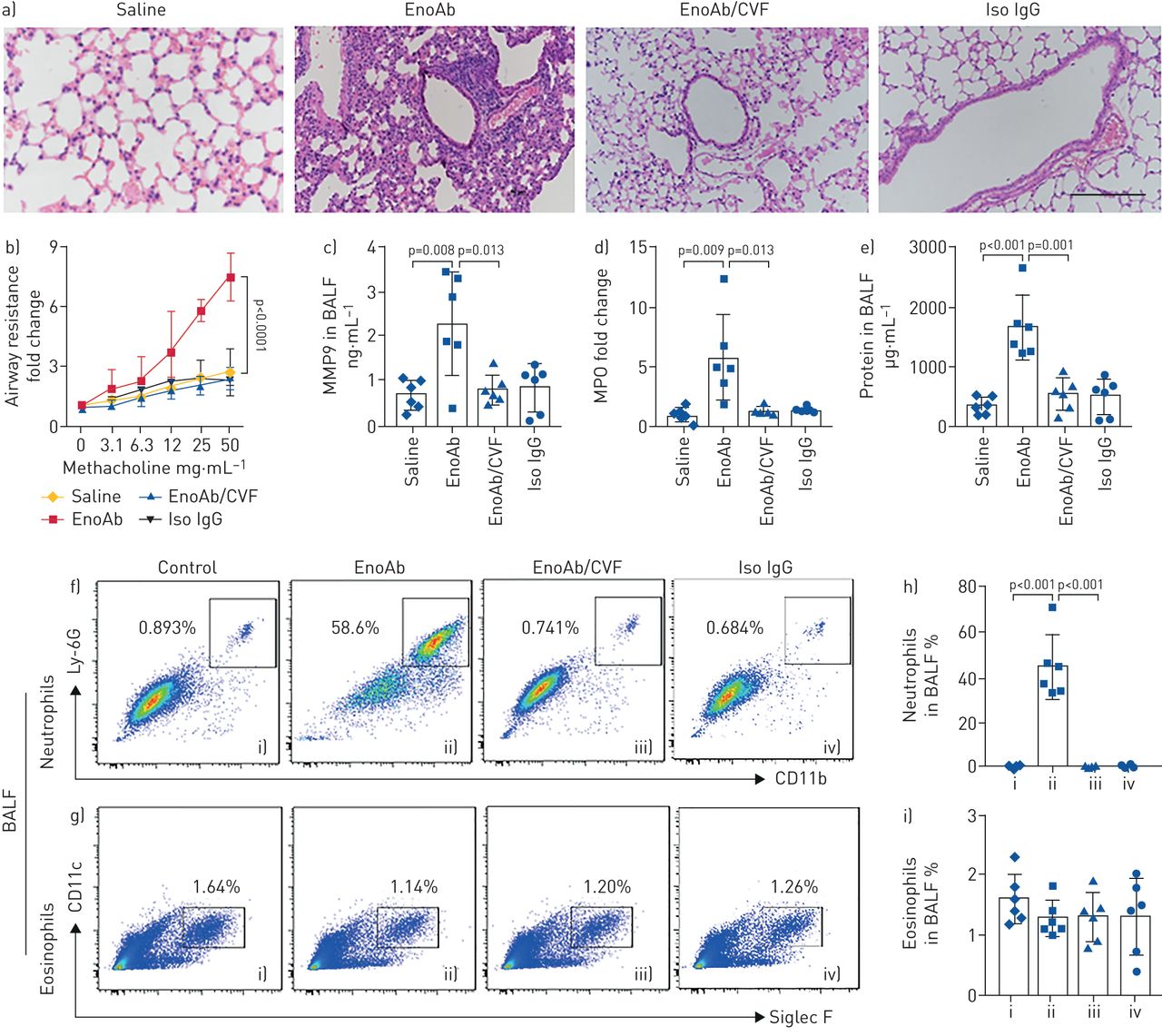
Exposure to EnoAb induces neutrophilic inflammation in the airway
To elucidate if the EnoAb can induce inflammation in the airway, naïve mice were treated with nasal drops containing EnoAb or saline for 6 days. In addition to inducing EnoAb/enolase complexes on the surface of airway epithelium, impairing epithelial barrier integrity, and activation of the complement system, as reported in figures 2–5, profound inflammatory cell infiltration in the lung tissues was observed (figure 6a); hypersensitivity was observed in mice (figure 6b); the levels of myeloperoxidase, matrix metallopeptidase-9 and total proteins in BALF were significantly higher in the EnoAb group than that in the saline group (figure 6c–e), and high frequency of neutrophils, but not eosinophils, was detected in BALF (figure 6f–i) as well as in the lung tissue (supplementary figure S9). Pre-treatment with CVF to quench complement abolished the EnoAb-induced inflammation in the lung (figure 6; supplementary figure S9). The results demonstrate that EnoAb can induce neutrophilic inflammation in the airway.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
EnoAb (an antibody against house dust mite-derived enolase) induces airway inflammation. Naïve BALB/c mice were treated with nasal drops containing saline or EnoAb or isotype immunoglobulin (Ig)G daily for 6 days. Mice received cobra venom factor (CVF; 200 U·kg−1, intraperitoneally) 3 days prior to the beginning of nasal drops. a) Representative lung histological images (×200) show inflammation in the lungs of mice treated with EnoAb. b) Airway resistance of mice. c–e) Levels of c) matrix metallopeptidase (MMP)9, d) myeloperoxidase (MPO) and e) total proteins in bronchoalveolar lavage fluid (BALF). f and g) Gated dot plots indicate f) neutrophils and g) eosinophils in BALF. h and i) Bars show the summarised data of h) neutrophils and i) eosinophils in BALF. Each group consists of six mice. Each data point inside bars present data from one mouse. Statistical method: ANOVA followed by Dunnett test. Data are presented as mean±sem. Scale bar=200 μm. Ig: immunoglobulin.
Discussion
The present data reveal a novel phenomenon that a cross-antibody, EnoAb, against HDM-derived enolase can initiate neutrophilic inflammation in the airway. Because HDM is the most prevalent allergen involved in the pathogenesis of airway allergic disorder, we investigated the role of EnoAb in the induction of airway inflammation. Most previous studies about HDM-induced airway inflammation focused on the role of IgE and HDM allergens in the pathogenesis of allergic asthma; the present data pinpoint another aspect of HDM: EnoAb, an IgG antibody, is also involved in the induction of airway inflammation. In line with previous studies, we also found eosinophil infiltration in the airway inflammation induced by EnoAb, while the most conspicuous finding of this study was that neutrophilic asthma was positively correlated with the serum HDM sIgG levels (p<0.01), although the correlation coefficient is rather low (∼0.3–0.4). The neutrophilic inflammation was reproduced in the airway tissues of mice by exposing mice to EnoAb, the cross-antibody induced by HDM-derived enolase.
The data show a conspicuous phenomenon that ∼50% of 16 376 mite genes are similar to human genes at a variety of ranges. This feature suggests that some proteins shared by mites and humans are partially similar in their molecular structure. It is known that a protein antigen molecule has many epitopes; each epitope may elicit the generation of a specific antibody [24]. Thus, it is probable that human immune system-producing antibodies against mite antigens may also attack human tissues. Mite-derived proteins are distributed extensively in the human living environment [25], and easily enter the human body through inhalation. Thus, mite-derived proteins contact human airway tissues constantly, and are the most common allergens of airway allergy [26]. Apart from HDM, there are many sources of enolase; for example, Candida albicans enolase has almost the same sequence identity as HDM and human enolase and, unsurprisingly, the conserved regions are very similar [27]; non-acarid sources, scabies or food contaminated by mites could be an important source of enolase that might cross-react with human tissues. Enolase also exists in fungus, e.g. Alternaria alternata [28]. Fungus-derived molecules are important allergens in the induction of asthma [29]. Thus, the role of enolase from different sources in the pathogenesis of asthma is worth investigating further.
The role of HDM-related allergic response in the pathogenesis of asthma has been studied extensively. It is accepted that IgE, mast cells and eosinophils are central effectors in the pathogenesis of asthma attacks [30]. The present data reveal that another important factor, the mite-specific IgG, is involved in the pathogenesis of mite antigen-related inflammation of the airway tissues. Others also found that asthma patients had mite antigen-specific IgG in the serum [31, 32]. Although some subtypes of IgG, such as IgG1, are beneficial to asthma patients by binding the IgG-inhibitory Fc γ-receptor (FcγRIIB) to suppress C5a receptor-mediated inflammation [33], the present data show that one of the mite antigen-specific IgGs, EnoAb, has another functional aspect that attacks the enolase of the airway tissues. EnoAb forms a complex with enolase in airway epithelial cells. The complex activates complement and initiates neutrophilic inflammation in the airway. The data also show that the serum HDM-specific IgG3 levels are higher in neutrophil-type asthma patients than that in eosinophil-type asthma patients. Among the IgG subtypes, IgG3 has stronger effects on activating complement, while IgG1 mainly works in IgE production and IgG4 cannot activate complement [34]. Complement activation is one of the causative factors in neutrophilic inflammation [13]; our data are in line with this, showing that sputum neutrophil counts are positively correlated with serum sIgG3, whereas no correlation was detected between serum sIgG3 and sputum eosinophil counts since eosinophils are more relevant to the IgE-mediated allergic inflammation [35].
The present data show that EnoAb induced airway epithelial barrier dysfunction. Epithelial barrier dysfunction in the airway mucosa of asthma has been recognised. It is a consensus that the disintegration of the epithelial barrier plays an important role in the pathogenesis of airway inflammation [36]. Our early work showed that the dysfunctional epithelial barrier allowed macromolecular substances, such as foreign protein antigens, to penetrate into the deep regions of the mucosa [6], where the foreign antigens gained the opportunity to contact various immune cells to elicit an immune response [37]. A variety of factors have been found to affect epithelial barrier functions, such as psychological stress [38], air pollution [39], allergic response [40], etc. The present data reveal that a novel factor, a cross-antibody against mite-derived enolase, can induce airway epithelial barrier dysfunction.
By directly dropping EnoAb into the airway, we induced airway inflammation in mice, which was characterised by profound infiltration of neutrophils in the lung tissues. This inflammatory type is the same as the neutrophilic phenotype of asthma [17]. Neutrophil infiltration in the lung is often associated with the severity of asthma, such as severe asthma and corticosteroid-resistant asthma [1]. This type of asthma is common in the clinic [17], while its pathogenesis remains elusive. The present data provide mechanistic evidence for the explanation of this phenomenon. The mite-derived molecule, the enolase, induces cross-antibodies that may be the important factor in the initiation of neutrophilic airway inflammation.
The data can answer Witebsky's postulates for the definition of autoimmune diseases: direct evidence of transfer of disease-causing antibody [41]. By treating naïve mice with nasal drops containing EnoAb, neutrophilic inflammation was induced in the lung. In other words, the mite-derived homologous antigen, enolase, can induce autoimmune disorders in the lung. As mentioned earlier, we identified 16 376 Der f genes, in which ∼50% of Der f genes had identical gene sequences to human genes, varying from 2% to 100%. Therefore, besides enolase, whether other Der f genes with identical sequences to human are involved with the pathogenesis of human autoimmune diseases following the cross-antibody concept needs further investigation. Whether all foreign antigens have identical gene sequences of human genes that are associated with autoimmune diseases in human is an interesting research topic which needs to be investigated.
In addition, we detected high serum IgE levels in patients with the eosinophilic asthma subtype. IgE is an important mediator of allergy. By binding the high-affinity receptors, IgE can sensitise mast cells. Re-exposure to specific antigens triggers asthma attacks by activating sensitised mast cells. Administration of anti-IgE antibodies can alleviate asthma attacks or attenuate chronic rhinosinusitis with nasal polyps and nasal allergy [42, 43]. However, not all asthma patients well respond to anti-IgE therapy [42]. Our data provide mechanistic evidence. The IgE levels were significantly lower in neutrophilic asthma patients. These asthma patients may not be suitable to be treated with anti-IgE antibodies. The data suggest that treating asthma with anti-IgE antibodies may only be effective in those with high IgE titres [42].
In summary, the present data show that Der f-derived enolase has identical gene sequences as 72% of the sequences of the human enolase gene, which can induce a cross-antibody in the human body. Adoptive transfer with the cross-antibody (generated from mouse) can induce neutrophilic inflammation in the mouse airway mucosa. We conclude that Der f-derived enolase can induce neutrophilic inflammation in the mouse airway.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material ERJ-02375-2019.SUPPLEMENT
Shareable PDF
Supplementary Material
This one-page PDF can be shared freely online.
Shareable PDF ERJ-02375-2019.Shareable
Footnotes
This article has supplementary material available from erj.ersjournals.com
Author contributions: J. Lin, N. Huang, J. Li, X. Liu, Q. Xiong, C. Hu, D. Chen, L. Guan, K. Chang and D. Li performed experiments and analysed data. S.K-W. Tsui provided bioinformatic analysis. J. Li provided clinical materials. P-C. Yang, Z. Liu and N. Zhong organised the study and supervised experiments. P-C. Yang, Z. Liu and J. Lin designed the project. P-C. Yang wrote the manuscript. All authors reviewed the manuscript.
Conflict of interest: J. Lin has nothing to disclose.
Conflict of interest: N. Huang has nothing to disclose.
Conflict of interest: J. Li has nothing to disclose.
Conflict of interest: X. Liu has nothing to disclose.
Conflict of interest: Q. Xiong has nothing to disclose.
Conflict of interest: C. Hu has nothing to disclose.
Conflict of interest: D. Chen has nothing to disclose.
Conflict of interest: L. Guan has nothing to disclose.
Conflict of interest: K. Chang has nothing to disclose.
Conflict of interest: D. Li has nothing to disclose.
Conflict of interest: S.K-W. Tsui has nothing to disclose.
Conflict of interest: N. Zhong has nothing to disclose.
Conflict of interest: Z. Liu has nothing to disclose.
Conflict of interest: P-C. Yang has nothing to disclose.
Support statement: This study was supported by grants from the National Key Research and Development Program on Precision Medicine (No.2016YFC0905802, 2016YFC0903700), Natural Science Foundation of China (31729002, 91542104, 31570932), Science and technology project of Guangdong Province (No.2014B090901041, 2016A020216029), Opening fund of State Key Laboratory of Respiratory Disease (No.SKLRD2016ZJ001), Shenzhen Scientific Technology Basic Research Projects (No.KQTD20170331145453160, JCYJ20160328144536436, KQJSCX20180328095619081), Shenzhen Nanshan District Pioneer Group Research Funds (No.LHTD20180007) and Guangdong Provincial Key Laboratory of Regional Immunity and Diseases (2019B030301009).
- Received December 11, 2019.
- Accepted July 30, 2020.
- Copyright ©ERS 2021
References










