





Figures
- FIGURE 1
Stereotypical clinical course of coronavirus disease 2019 complicated by acute respiratory distress syndrome (ARDS). Mean incubation period and viral load threshold cycle (Ct) values at days 0, 7, 14 and 21 according to He et al. [12]. Median days from symptom onset to dyspnoea (interquartile range 4–9 days), ARDS (interquartile range 8–15 days), and discharge or death (interquartile range 17–25 days) according to Zhou et al. [14].
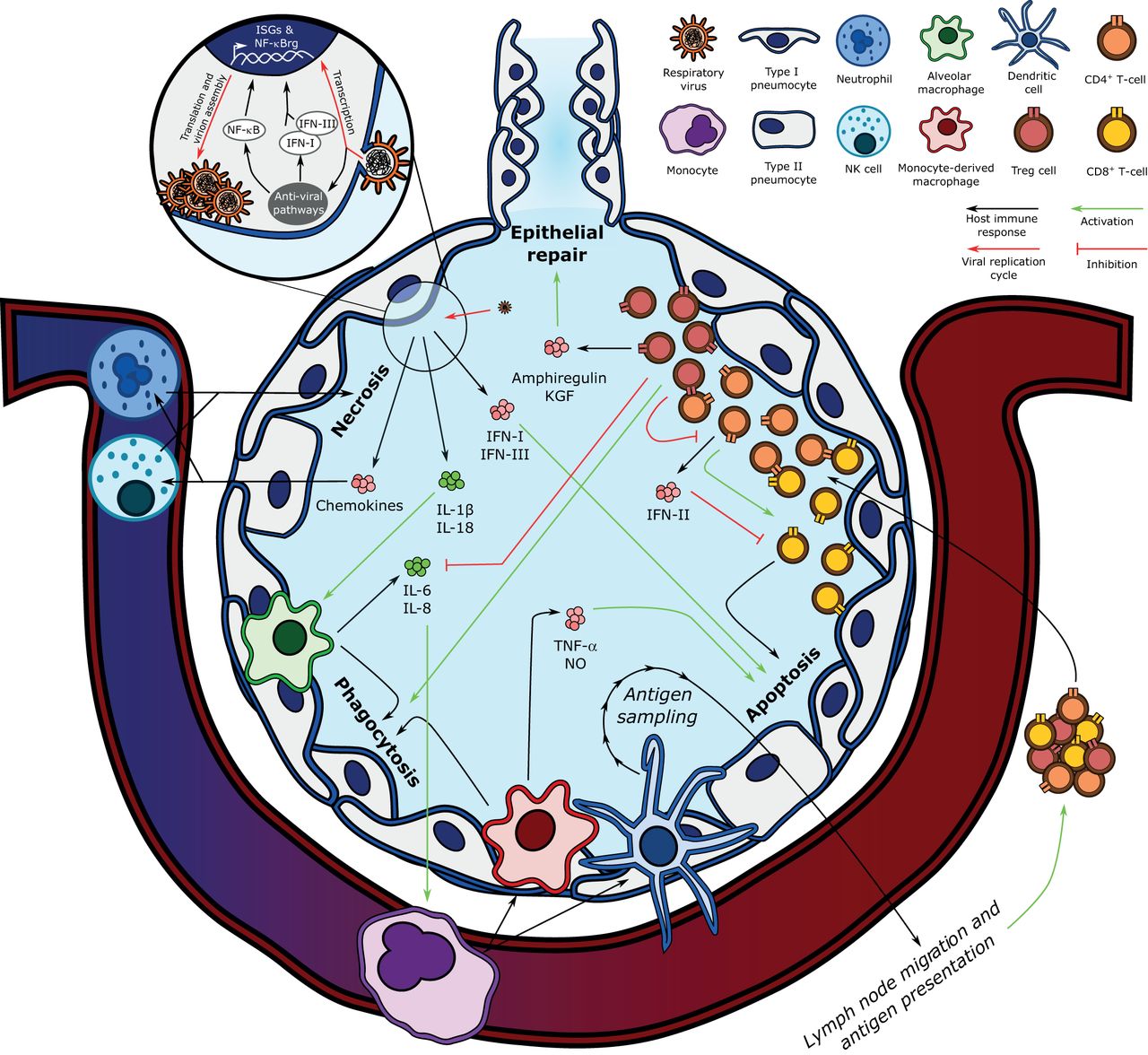
- FIGURE 2
Key components of the pathogenesis of influenza-induced acute respiratory distress syndrome. Influenza's replication cycle requires the presence of viral RNA in the host epithelial cell's cytoplasm, which is recognised by anti-viral pathways such as Toll-like receptors, mitochondria-associated anti-viral signalling, and the NLRP3 inflammasome. The host cell responds by upregulating nuclear factor (NF)-κB-regulated genes (NF-κBrg) and secreting type-I and -III interferons (IFN-I and IFN-III), which stimulate the transcription of interferon-stimulated genes (ISGs). Infected epithelial cell-derived IFN-I and IFN-III induce epithelial cell apoptosis, and interleukin (IL)-1β and IL-18 secretion activates alveolar macrophages, which phagocytose apoptotic cells. Alveolar macrophages also secrete IL-6 and IL-8, which recruit the central immune system, including monocytes, to the lung parenchyma. Monocytes differentiate into monocyte-derived macrophages and dendritic cells. Monocyte-derived macrophages also contribute to infected epithelial cell phagocytosis and secrete tumour necrosis factor (TNF)-α and nitric oxide (NO) to activate epithelial cell apoptosis. Dendritic cells link the innate and adaptive immune responses by sampling alveolar antigens and migrating to lymph nodes, where they present viral antigens to naïve T-cells. After undergoing antigen-specific clonal expansion, T-lymphocytes are mobilised to the lung. CD8+ T-cells exert their cytotoxicity on viral antigen-presenting cells when co-stimulated by CD4+ T-cells, which also secrete type-II interferon (IFN-II), a negative regulator of CD8+ T-cell activity. In turn, CD4+ T-cells are negatively regulated by regulatory T-cells (Treg), which also promote epithelial repair via the secretion of amphiregulin and keratinocyte growth factor (KGF), and attenuate inflammation by inhibiting alveolar macrophage-derived IL-6 and IL-8 secretion while potentiating their phagocytic activity (efferocytosis). NK: natural killer.
- FIGURE 3
Examples of the contribution of hallmarks of ageing to the age-related predisposition to severe coronavirus disease 2019 (COVID-19). Age-related mitochondrial dysfunction can induce epigenetic changes in regulatory T-cells (Treg), which impair their pro-recovery functions to hinder proper resolution of inflammation and repair of lung injury. Monocytes and naïve T-lymphocytes undergo cellular senescence following telomere attrition from sustained replication, impairing the host's ability to mount an efficient immune response to a viral challenge or create a memory T-cell response to vaccines. Finally, altered intercellular communication underlies the low-grade inflammation associated with ageing, which contributes to the development of age-related comorbidities.



{kind=link}
{kind=link}
{kind=link}
Supplementary Materials
Supplementary Material
This one-page PDF can be shared freely online.
Shareable PDF ERJ-02049-2020.Shareable











Jump To
- Article
- Abstract
- Abstract
- Main novel ideas and hypotheses
- Introduction
- Viral replication cycle and clinical presentation
- Current understanding of viral ARDS and host immune responses
- Insights into COVID-19-induced ARDS
- Potential mechanisms through which ageing drives severe COVID-19
- Conclusion
- Shareable PDF
- Footnotes
- References
- Figures & Data
- Info & Metrics