





Abstract
rhCCSP prevents airway neutrophilia. This might occur via a direct neutralisation of IL8 in humans. http://bit.ly/2UGJlGW
Abstract
Club cell secretory protein (CCSP) knockout mice exhibit increased airway neutrophilia, as found in chronic obstructive pulmonary disease (COPD). We therefore investigated whether treating COPD airway epithelia with recombinant human CCSP (rhCCSP) could dampen exaggerated airway neutrophilia.
Control, smoker and COPD air–liquid interface (ALI) cultures exposed to cigarette smoke extract (CSE) were treated with and without rhCCSP. The chemotactic properties of the supernatants were assessed using Dunn chambers. Neutrophil chemotaxis along recombinant human interleukin 8 (rhIL8) gradients (with and without rhCCSP) was also determined. rhCCSP–rhIL8 interactions were tested through co-immunoprecipitation, Biacore surface plasmon resonance (SPR) and in silico modelling. The relationship between CCSP/IL8 concentration ratios in the supernatant of induced sputum from COPD patients versus neutrophilic airway infiltration assessed in lung biopsies was assessed.
Increased neutrophilic chemotactic activity of CSE-treated ALI cultures followed IL8 concentrations and returned to normal when supplemented with rhCCSP. rhIL8-induced chemotaxis of neutrophils was reduced by rhCCSP. rhCCSP and rhIL8 co-immunoprecipitated. SPR confirmed this in vitro interaction (equilibrium dissociation constant=8 µM). In silico modelling indicated that this interaction was highly likely. CCSP/IL8 ratios in induced sputum correlated well with the level of small airway neutrophilic infiltration (r2=0.746, p<0.001).
CCSP is a biologically relevant counter-balancer of neutrophil chemotactic activity. These different approaches used in this study suggest that, among the possible mechanisms involved, CCSP may directly neutralise IL8.
Introduction
Imbalance between injury and repair universally trends with inflammatory processes. Exaggerated neutrophilic influx is a hallmark of most chronic airway diseases, including neutrophilic asthma, cystic fibrosis, idiopathic pulmonary fibrosis and chronic obstructive pulmonary disease (COPD) [1]. COPD is a growing cause of morbidity and mortality worldwide. In addition, increased neutrophilic inflammation is characteristic in patients with severe disease [2] or with acute exacerbations [3]. Airway neutrophilia is associated with an accelerated decline in lung function [4, 5] and with the severity of peripheral airway dysfunction in COPD [6, 7].
Chemotaxis, a process whereby cells migrate according to a chemical concentration gradient, critically mediates neutrophil recruitment in tissue [8]. Neutrophils express multiple chemoattractant receptors and can be sensitive to individual gradients or combinations thereof. Levels of several chemoattractants for neutrophils are increased in the bronchial secretions of COPD patients. The most abundant and studied are interleukin 8 (IL8) and leukotriene B4 (LTB4) [9, 10]. These chemokines bind to neutrophil CXC chemokine receptor types 1 and 2 (CXCR1/CXCR2) and LTB4 receptor (BLT1) G protein-coupled receptor (GPCR), respectively, to amplify cell recruitment [11]. Past in vitro studies often employed N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP) because of its highly potent chemoattractant ability; fMLP ligates with neutrophil formyl peptide receptors (FPRs) [12]. A previous study has shown that COPD neutrophils, relative to those from healthy patients, are disoriented, migrating faster but with less accuracy [13].
The club cell secretory protein (CCSP, CC10 or SCGB1A1) produced by non-ciliated club cells is known to have immunoregulatory and anti-inflammatory functions [14–17]. However, the mechanisms of action remain largely unknown in the absence of an identified specific receptor. CCSP is defective in COPD airways [17–19]. Appropriately, Laucho-Contreras et al. [18] demonstrated that increased lung neutrophilia was observed in CCSP−/− mice exposed to cigarette smoke, as seen in COPD airways. Moreover, this could be rescued by exogenous CCSP supplementation.
The exact mechanisms by which CCSP mediates its anti-inflammatory activities, including whether or not it binds to a receptor, have not yet been determined. To date, its paracrine signalisation is unknown and remains hypothetical. Of note, CCSP levels are known to be reduced in sera, sputum and bronchoalveolar lavage fluid from COPD patients. Furthermore, two studies demonstrated that serum CCSP levels predict accelerated lung function decline [20, 21].
We hypothesised that injury/repair imbalance may explain exaggerated airway neutrophilia and that CCSP defects could be involved in this process in COPD patients. Our aim was to test whether recombinant human CCSP (rhCCSP) interacts with airway neutrophilic influx in a human model of COPD, and whether this interaction is due to an effect of CCSP on the airway epithelial cells or to an endogenous IL8 binding counter-balancer. For this purpose, air–liquid interface (ALI) cultures of epithelial cells were treated with rhCCSP (exposed to cigarette smoke extract (CSE) or not). The collected supernatants were used to assess neutrophilic chemotaxis. Furthermore, neutrophils were directly exposed to chemotactic agents and treated with rhCCSP. Their chemotactic parameters were also assessed. Finally, we developed an in vitro three-step demonstration of CCSP–IL8 direct binding, and sought possible converging evidence of the relationship between CCSP and IL8 in lung airways.
Materials and methods
Epithelial cell cultures and stimulation
Bronchial biopsies from control subjects (n=4), smokers (n=10) and COPD patients (n=13) were collected during surgery. Subjects were recruited at the Arnaud de Villeneuve Hospital, Montpellier, France. The local ethics committee, Comité de Protection des Personnes Sud-Mediterranée III, approved the study design (approval number: 2013 11 05; NCT02354677) and all patients agreed to participate by reading and signing written informed consent forms.
Human primary bronchial epithelial cells (HBECs) were cultured under ALI conditions, adapted from Gamez et al. [17] and Gras et al. [22]. Details on this method and on subject baseline characteristics are available in the supplementary material. Cells from bronchial biopsies of four control subjects, 10 smokers and 13 COPD patients were maintained in culture for 28 days to obtain a differentiated cell population with a pseudostratified mucociliary epithelium. Cells were treated for 24 h with or without CSE and with or without rhCCSP supplementation (3 µg·mL−1), applied at the apical surface. The supernatants were collected by washing the apical surface and were stored until used for neutrophil experiments and CCSP/IL8 ELISA test (Biovendor, Brno, Czech Republic; Diaclone, Besançon, France).
Cigarette smoke extract
CSE was prepared as previously described [17]. Briefly, the smoke of three commercial cigarettes (Marlboro) was drawn into a Pasteur pipette and bubbled into 30 mL of PBS. CSE was adjusted to pH 7.4 and sterilised through a 0.22 µm filter. We obtained a 100% stock solution of CSE diluted to 50% for the experiments.
Isolation of peripheral blood neutrophils
Neutrophils were isolated from the whole blood of five healthy control subjects and five COPD patients (approval number: 0811738) using discontinuous Percoll gradients followed by hypotonic lysis of residual red blood cells [23]. The neutrophils (purity at 95% and viability assessed by trypan blue exclusion of dead cells, at 95%) were re-suspended in RPMI 1640 medium containing 0.15% bovine serum albumin.
Neutrophil chemotaxis assays and analyses
The chemotaxis assays were performed using Dunn chambers [24]. Details on this method are available in the supplementary material. Chemotaxis was quantified using the x-forward migration index (XFMI) value, which is the ratio of the distance travelled during the acquisition and the final position of the neutrophils on the x-axis in the direction of the chemoattractant gradient.
ImageJ software (https://imagej.nih.gov/ij/) was used to track cells and analyse chemotaxis. All analyses were performed by a single analyst, blinded to the subject group.
Three sets of assays were performed. First, supernatants gathered by a brief wash at the apical side of ALI cultures of airway epithelial cells at baseline were applied in the outer ring and neutrophil chemotaxis from both controls and COPD patients was assessed. Second, supernatants gathered in the same way but treated with each of the above-mentioned conditions were similarly applied and assessed. Third, only pharmacological agents separately or combined were applied in the outer ring in order to delineate whether the observed results in the first set of assays could be modulated and reproduced without any epithelial interaction.
Co-immunoprecipitation
To detect whether a potential interaction exists between CCSP and IL8, co-immunoprecipitation and immunoblotting were performed using anti-Protein A Sepharose 4 Fast Flow magnetic beads (GE Healthcare, Uppsala, Sweden). Details on this method are available in the supplementary material. All immunoblots were developed and quantified using the Odyssey Infrared Imaging System (LI-COR Biosystems, Lincoln, NE, USA) and infrared-labelled secondary antibodies.
Biacore measurements
Real-time surface plasmon resonance (SPR) analyses were performed on a Biacore T200 apparatus (GE Healthcare) at 25°C, in HBS-EP+ buffer (10 mM HEPES, 150 mM NaCl, 3 mM EDTA, 0.05% polysorbate 20 surfactant). rhCCSP, diluted at 10 µg·mL−1 in 10 mM acetate pH 4, was covalently immobilised at 2400 resonance units on a CM5S sensor chip (GE Healthcare) using an amine coupling protocol according to the manufacturer's instructions.
Kinetic titration experiments (single-cycle kinetics) of IL8 were performed at a flow rate of 100 µL·min−1 with two-fold serial dilutions (62–1000 nM). All curves were evaluated (T200 Evaluation Software 3.0; GE Healthcare) after double-referencing subtraction using a bivalent fitting model. This experiment was repeated with a new immobilisation of rhCCSP with a similar result.
Three-dimensional protein modelling and protein–protein docking
A homology model of the homo sapiens CCSP (hsCCSP) homodimer has been generated according to the crystal structure of the rabbit ortholog (Protein Data Bank (PDB): 2UTG), using the software Modeller [25]. The protein–protein docking server ZDOCK [26] was used with blind docking parameters to explore the putative interaction between the three-dimensional (3D) homology model of hsCCSP and the experimental human IL8 structure (PDB: 3IL8).
Surgical lung samples and induced sputum collection
A cross-sectional cohort of 10 COPD patients undergoing surgery for peripheral nodules was recruited after patients signed an informed consent form. This study was approved by the local ethics committee (Comité de Protection des Personnes de Nimes; AOI 2000, CPP 000901, DGS 2001/0075). Both CCSP and IL8 concentrations were assessed in the induced sputum of patients 3–5 days before they underwent lung surgery. Surgical lung samples were optimally processed to allow for quantification of neutrophil elastase-positive cells in the submucosal area of small airways at a maximum depth of 100 µm beneath the basement membrane. Methodological details are available in the supplementary material.
Statistical analysis
Details on statistical analysis are available in the supplementary material.
Results
COPD airway epithelia attracted neutrophils
Dunn chemotaxis chambers were filled with ALI epithelial supernatants. Figure 1a displays representative examples of healthy and COPD neutrophil migratory tracks induced by supernatants from ALI-reconstituted airway epithelia derived from healthy controls, smokers and patients with COPD. Consistent with our previous observations, fMLP was confirmed as a positive control, and PBS as a negative one (p<0.0001) (figure 1b). Although epithelial supernatants from all groups increased neutrophil chemotaxis, only neutrophil chemotaxis induced by COPD supernatants was significantly higher than that induced by control supernatants (p=0.0012) (figure 1b). Neither CCSP nor IL8 concentration was measured in this set of assays.

At steady state, air–liquid interface (ALI)-reconstituted chronic obstructive pulmonary disease (COPD) airway epithelia maintained in a clean environment promoted exaggerated neutrophilic chemotaxis. a–c) Representative examples of healthy neutrophil migratory tracks induced by supernatants of ALI-reconstituted airway epithelium. The large red arrows represent the chemoattractant signal (30 min of phase time-lapse recordings). a) Healthy control: number of tracks 76; counts left: 32, right: 44; centre of mass x=4.18, y=0.32. b) Smoker: number of tracks 74; counts left: 11, right: 63; centre of mass x=43.62, y=−7.03. c) Patient with COPD: number of tracks 95; counts left: 20, right: 75; centre of mass x=33.86, y=0.74. The COPD neutrophil migratory tracks are induced by COPD ALI supernatant epithelial cell cultures. d) The x-forward migration index (XFMI) of healthy neutrophils on N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP) and epithelial supernatant gradients (control subjects n=4, smoker subjects n=3, COPD patients n=5). Epithelial supernatants from all groups increased neutrophil chemotaxis. COPD supernatants attracted neutrophils. Approximately 100 tracked neutrophils by condition. Data are expressed as mean±sem. Paired comparisons were made using Mann–Whitney tests. #: significant differences between compared groups (p=0.0012); *: significant differences compared to control (p<0.05).
Treating the airway epithelium with rhCCSP prevented neutrophil chemotaxis when exposed to CSE to an extent that fits with CSE-induced IL8 release
We further focused on whether treating CSE-exposed COPD airway epithelia with rhCCSP could reduce exaggerated neutrophil influx. As expected, smoker and COPD epithelia exposed to CSE had increased neutrophil chemotaxis (figure 2a, b). When smoker and COPD airway epithelia were exposed to CSE, we observed for the first time on human cells that rhCCSP prevented neutrophil chemotaxis, which returned to nearly baseline values (p<0.0001 and p=0.0035, respectively) (figure 2a, b). IL8 and CCSP concentrations in the supernatants were assessed (figure 2c, d; supplementary figure S1). In this set of experiments, CSE tended to induce IL8 release, but there were no differences between smokers and COPD patients. Similar chemotactic measures were obtained with neutrophils from COPD patients (supplementary figure S2). It should be noted that XFMI was used to represent neutrophil chemotaxis in our study, except for COPD neutrophils. Because the latter are disoriented (according to the literature [13]), the distance travelled by the neutrophils was a more appropriate measure for demonstrating that rhCCSP had the same effect on COPD neutrophils as on healthy neutrophils.
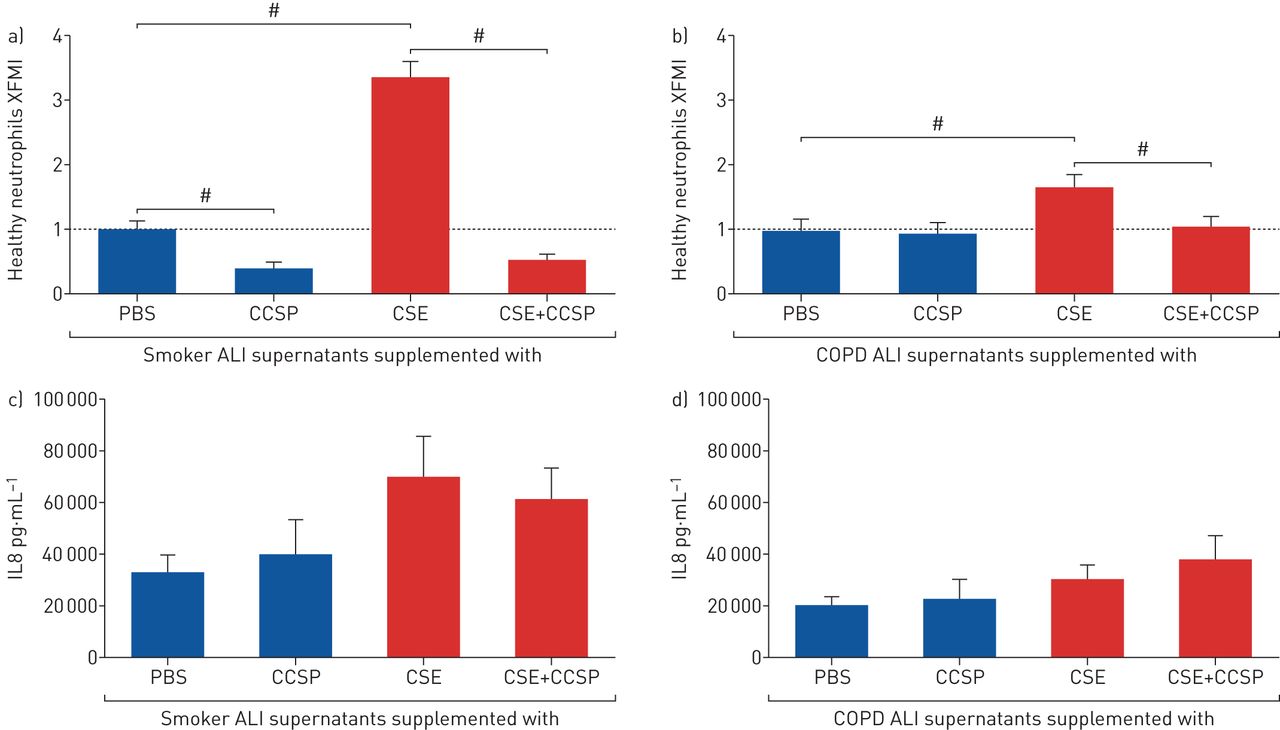
Treating the airway epithelium with exogenous club cell secretory protein (CCSP) prevented cigarette smoke extract (CSE)-induced neutrophil chemotaxis. a) The x-forward migration index (XFMI) of healthy neutrophils on a gradient of smoker air–liquid interface (ALI) supernatants (n=7) stimulated by CSE with or without CCSP supplementation, relative to PBS. CSE increased neutrophil chemotaxis (p<0.0001). CCSP inhibited neutrophil chemotaxis when epithelia were exposed to CSE (p<0.0001). b) The XFMI of healthy neutrophils on a gradient of chronic obstructive pulmonary disease (COPD) ALI supernatants (n=8) stimulated by CSE with or without CCSP supplementation, relative to PBS. CSE increased neutrophil chemotaxis (p=0.0056). CCSP inhibited neutrophil chemotaxis when epithelia were exposed to CSE (p=0.0035). Approximately 100 tracked neutrophils by condition. Data are expressed as mean±sem. #: paired comparisons were made using Mann–Whitney tests. c, d) CSE-induced interleukin 8 (IL8) secretion by epithelial cells from smokers (n=7) and COPD patients (n=8), with or without CCSP supplementation. CSE tended to increase IL8 secretion. CCSP supplementation had no effect on IL8 secretion.
rhCCSP directly inhibited neutrophil chemotaxis induced by rhIL8 and by fMLP
Neutrophil migration along the fMLP gradient was reduced by rhCCSP (p<0.0001 for the XFMI) but not along the rhIL8 gradient (figure 3a). Likewise, no CXCR1/2 inhibitor effects were observed (figure 3a). Because XFMI is the ratio of the distance travelled during acquisition and the final neutrophil positions on the x-axis in the direction of the chemoattractant gradient, we analysed these parameters (i.e. the so-called X endpoints and the distance travelled). Interestingly, the final position of healthy neutrophils in the presence of rhCCSP was significantly delayed in the direction of the rhIL8 gradient (p=0.028) to a similar extent as that for the CXCR1/2 inhibitors (figure 3b), but these neutrophils travelled shorter distances (p<0.0001) (figure 3c). When performed with neutrophils from COPD patients, rhIL8-induced neutrophil chemotaxis, as assessed by XFMI (p<0.0001), X endpoints (p<0.0001) and the distance travelled (p<0.0001), was inhibited by rhCCSP (supplementary figure 3A–C).

Stimulated-neutrophil migration was slowed down by club cell secretory protein (CCSP). a) The x-forward migration index (XFMI) of healthy neutrophils on N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP) or interleukin 8 (IL8) gradients with or without CCSP and with or without CXCR1/2 inhibitors. CCSP inhibited the chemotaxis of neutrophils stimulated by fMLP (p<0.0001), but no difference was observed for IL8-induced neutrophil chemotaxis. No effects associated with CXCR1/2 inhibitors were observed. b) The X endpoints of healthy neutrophils on fMLP or IL8 gradients with or without CCSP and with or without CXCR1/2 inhibitors. fMLP was confirmed as a positive control and PBS (Control) as a negative one (p<0.0001). CCSP inhibited the movement of neutrophils in the direction of the fMLP gradient (p<0.0001), and in the direction of the IL8 gradient (p<0.0001) to a similar extent as found for CXCR1/2 inhibitors. c) The distance travelled by healthy neutrophils on fMLP or IL8 gradients with or without CCSP and with or without CXCR1/2 inhibitors. CCSP inhibited the movement of neutrophils induced by the fMLP gradient (p=0.0005) and the IL8 gradient (p<0.0001). Approximately 100 tracked neutrophils by condition. All conditions were reproduced three times. Data are expressed as mean±sem. Paired comparisons were made using Mann–Whitney tests. #: significant differences between compared groups (p<0.05); *: significant differences compared to control (p<0.05). ns: nonsignificant.
rhCCSP and rhIL8 had a direct interaction
The hypothetical direct rhCCSP–rhIL8 interaction was evaluated by co-immunoprecipitation with anti-IL8 antibodies followed by anti-CCSP antibody blotting. We found that rhCCSP co-immunoprecipitated with rhIL8 (figure 4a, b). The reverse demonstration was performed (figure 4a, c) and confirmed this interaction. This finding was validated by SPR analyses using single-cycle kinetics without regeneration (figure 4d). Accordingly, we determined an equilibrium dissociation constant (KD) of 8 µM. 3D protein modelling and protein–protein docking were performed in order to illustrate the molecules' in silico compatibility (figure 4e). The docking results of ZDOCK [26] demonstrated a set of hsCCSP–hsIL8 complex structures, with the five top-ranked predictions (selected based on the internal ZDOCK scoring function) adopting similar conformations. Given the known binding of IL8 to CXCR1/2 on neutrophils, we next compared both binding sites. The superimposition of the hsCCSP–hsIL8 complex model with the experimental structure of hsCXCR1(C-terminal part)–hsIL8 complex (1ILP) showed that CXCR1 and CCSP would be located in the same area.

In vitro, club cell secretory protein (CCSP) interacted with interleukin 8 (IL8). a) Western blot of IL8 immunoprecipitation detected with CCSP antibodies (Ab). Lane 1: negative control. Lane 2: positive controls (recombinant human (rh)IL8+rhCCSP). Lane 3: rhIL8 immunoprecipitated with CCSP detected with CCSP Ab (red band). rhCCSP co-immunoprecipitated (Co-IP) with IL8 detected with IL8 Ab (green band). b, c) CCSP immunostaining intensity of the IL8 immunoprecipitation (b) and IL8 immunostaining intensity of the CCSP immunoprecipitation (c), relative to negative control. No error bars are shown because the immunoprecipitation was performed once and then reproduced with the inverse relationship. d) Single-cycle kinetics of IL8 on immobilised CCSP: two-fold serial dilutions (62 nM–1000 nM) were injected on immobilised CCSP at a flow rate of 100 µL·min−1 with a contact time of 60 s and 400 s of dissociation after the last injection. The dotted line represents the fitting curve using a bivalent model. Association rate constant for the first site (Ka1) (1/ms)=(2.36±0.06)×10+3; dissociation rate constant for the first site (Kd1) (1/s)=(1.89±0.04)×10−2; equilibrium dissociation constant (KD)1 (M)=8.02×10−6; Chi-squared (RU2)=0.675. e) Superimposition of the homo sapiens (hs)CCSP–hsIL8 complex (red) with the experimental structure of hsCXCR1 (C-terminal part)/hsIL8 complex (Protein Data Bank: 1ILP; CXCR1 in green and IL8 in blue).
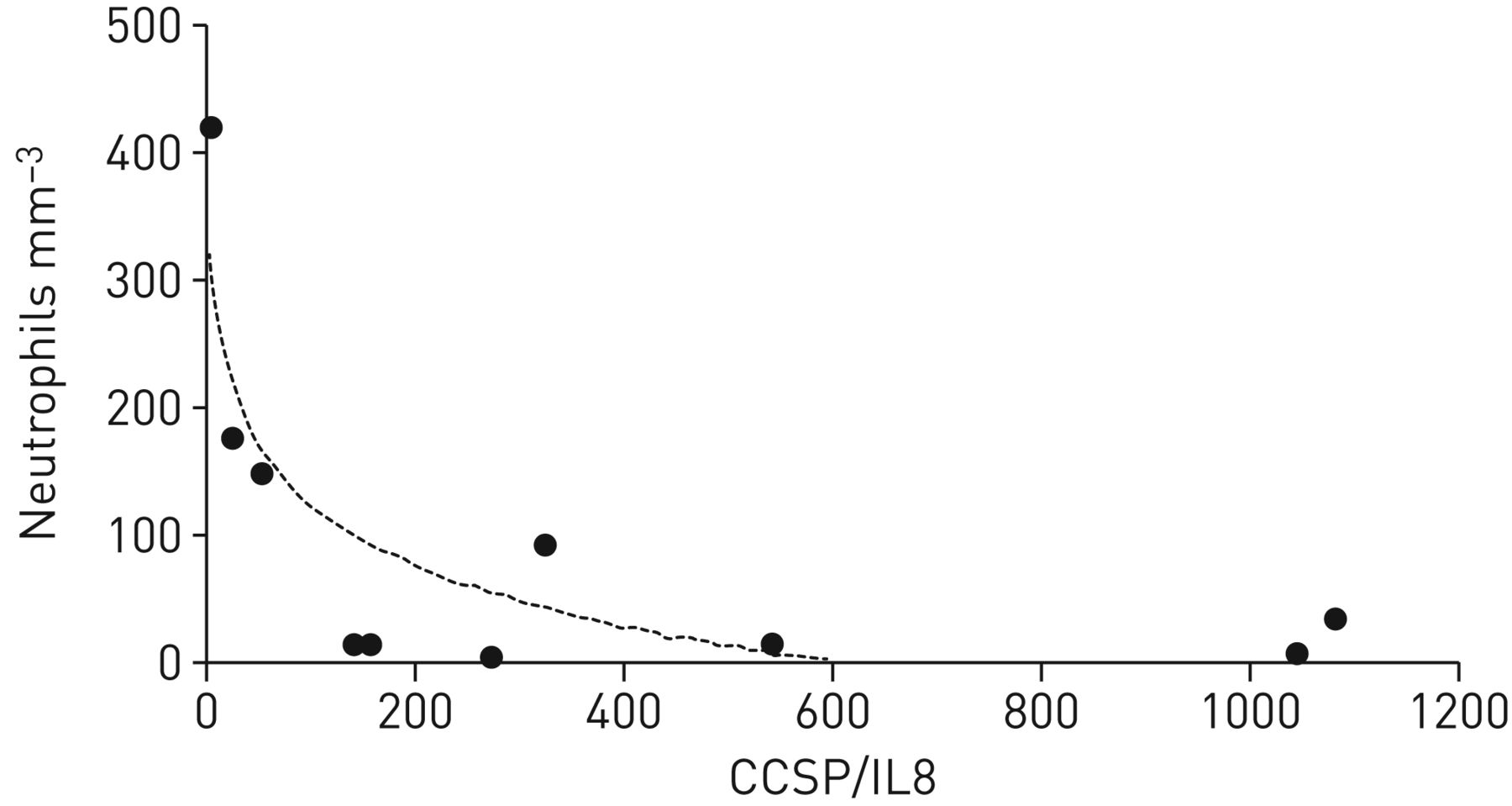
Small airway wall infiltration by neutrophils negatively correlated with the CCSP/IL8 ratio measured in the induced sputum of COPD patients undergoing surgery
In order to verify whether these findings are observable in vivo, neutrophilic infiltration of the small airway wall of COPD patients was quantified in human lung samples obtained during surgery. These results correlated with the CCSP/IL8 ratio assessed in the induced sputum collected before the surgery. The level of this negative correlation was as high as 0.746 (figure 5), indicating a greater intensity of neutrophilic infiltration when the CCSP/IL8 ratio was low and vice versa.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The small airway wall infiltration by neutrophils negatively correlated with the club cell secretory protein (CCSP) to interleukin 8 (IL8) ratio measured in the induced sputum of chronic obstructive pulmonary disease (COPD) patients (n=10) collected before surgery (r2=0.746). The level of this negative correlation indicated a greater intensity of neutrophilic infiltration when the CCSP/IL8 ratio was low and vice versa.
Discussion
In this study, we found that the airway epithelium of COPD patients reconstituted using ALI kept one important feature of the disease [17], in that it exerted an increased neutrophilic chemotactic activity at steady state: CSE-induced IL8 release by these epithelia obtained from either smokers and COPD was reduced by exogenous rhCCSP supplementation. The latter was true for the neutrophils of both controls and COPD patients, which is important considering the disorientation shown to affect COPD neutrophils [13]. Because rhCCSP reduced both fMLP- and rhIL8-induced chemotaxis irrespective of the presence of the airway epithelium at a magnitude similar to a CXCR1/2 antagonist, we strove to find evidence that rhCCSP could bind to rhIL8. We developed in vitro (through two-way co-immunoprecipitation and the Biacore SPR method), in silico and in vivo assays indicating convergent support for the observed correlation between CCSP/IL8 ratios in induced sputum versus levels of neutrophilic airway infiltration.
The development of COPD treatments is probably the greatest challenge for respiratory medicine for the next 20 years. The role played by airway inflammation is unclear at steady state, but neutrophilic bursts (associated with exacerbation episodes) are the established culprits of irreversible “stair-shaped” decline [27]. The role played by the only “bronchiolar-specific” epithelial cell type, the club cell, means that it is emerging as a potential regulator of these pathogenic COPD features [16, 17, 28]. We and others have been able to establish the role of club cells at the interface between structural alterations and inflammation, with possible modifications according to smoking status. In particular, serum CCSP levels are decreased in COPD and restoring their normal values decreases CSE-induced IL8 and LPS-induced mucin 5AC (MUC5AC) [17, 29]. Indeed, though rhCCSP appears ready to be tested in humans, major knowledge gaps concerning its mechanisms of action should be filled [30].
Different approaches have previously been used to assess CCSP effects on neutrophils in animals. In an equine model, incubating neutrophils with rhCCSP significantly reduced their respiratory oxidative bursts and increased their phagocytic activity [31]. Furthermore, neutrophil counts were increased in the bronchoalveolar lavage fluid of CCSP−/− mice exposed to cigarette smoke for 6 months [18]. With our human model, we assessed neutrophil chemotaxis in Dunn chambers using rhIL8 and fMLP as controls, and ALI epithelium supernatants. Using epithelial supernatants made sense because, physiologically, chemokines are also secreted by bronchial epithelial cells in order to attract neutrophils within the bronchial airway lumen [32]. These chemotactic chambers have the advantage of not being affected by gravity (e.g. compared to Boyden chambers), which is important for this model. Sapey et al. [13] first used this model to demonstrate that neutrophil chemotactic responses are different in COPD compared to in healthy subjects. Our initial results suggest that COPD airway epithelia are exerting increased neutrophilic chemotaxis at steady state. Interestingly, CSE exposure increased this chemotactic activity in both smokers and COPD and could be prevented by treating the epithelium with rhCCSP. We failed to pinpoint the intracellular signalling pathway involved. The small quantity of proteins extracted through ALI cultures on filters did not enable Western blot analysis of changes in NF-κB p65 subunits. NF-κB inhibition was suggested as a possible CCSP target by Laucho-Contreras et al. [18], whereas Tokita et al. [29] alternatively propose extracellular signal-regulated kinase 1/2. CCSP is also known to be a specific phospholipase A2 inhibitor, but this is highly controversial in the literature [18, 33]. Surprisingly, COPD airway epithelia in the present assays did not produce very high IL8 levels when exposed to CSE. The heterogeneity of COPD may at least partially explain this finding. In any case, the chemotactic activity measurements fit with the IL8 concentrations and support our overall interpretation of the findings.
Pharmacologically, neutrophil chemotaxis was increased by fMLP and rhIL8, and rhCCSP was efficient in reducing this activity. Candidate mediators of neutrophil chemotactic activity, such as LTB4 or growth-regulated oncogene-α, can be found in the literature [34, 35]. rhCCSP may act through those different mechanisms. It also suggests that rhCCSP interferes with neutrophilic chemotaxis not only via an epithelial effect, such as decreased IL8 signalling mimicked by CXCR1/2 antagonism, but also through FPR2 as suggested by Vasanthakumar et al. [36] and probably, given the unspecific effects of CSE, through broader interactions. Because this pharmacological modulation could be obtained irrespective of the presence of the airway epithelium, we strove to determine whether rhCCSP could directly neutralise IL8.
Over 20 years ago, a striking property was identified for CCSP: the presence of a hydrophobic pocket that binds airway pollutants [37, 38]. Our in silico experiments not only showed that hsCCSP directly binds hsIL8, but also that hsCCSP interacts with hsIL8 through the same binding site at CXCR1. As a consequence, CCSP and CXCR1 most likely compete with each other for IL8 binding (suggesting that IL8 can either bind CCSP or CXCR1, but not both at the same time). Treatment with an antagonist of CXCR2 (another IL8 receptor) led to significant improvement in forced expiratory volume in 1 s in patients with COPD, suggesting clinically important anti-inflammatory effects associated with CXCR2 antagonism [39]. Such results further support CCSP as a credible therapeutic strategy. We hypothesised that rhCCSP could directly trap rhIL8. Co-immunoprecipitation assays were reproduced twice and bi-directionally. SPR, an up-to-date real-time and label-free technology [40], was used to measure the direct interaction of rhIL8 on immobilised rhCCSP from dose–response curves (KD=8 µM). Whether these rhCCSP–rhIL8 complexes are biologically active or not is difficult to address. The reality of this interaction with non-recombinant protein remains to be demonstrated.
In vivo, we observed a statistically significant relationship between the intensity of the neutrophilic airway inflammation in the small airway of COPD patients and the CCSP/IL8 ratio measured in the induced sputum collected just before the lung surgery. Although inconclusive and not evidencing any kind of causality, this observation is convergent with the hypothesis. Once again, we acknowledge that this convergence should not be over-interpreted.
In conclusion, this study confirms that ALI-reconstituted COPD airway epithelia exert increased neutrophilic chemotactic activity and that rhCCSP is likely able to prevent airway neutrophilia in humans. We suggest that multiple mechanisms of action exist, including the possibility of direct IL8 neutralisation. A therapeutic strategy that limits airway neutrophilia must simultaneously address potential infection risks. Compared to CXCR1/2 blockades that induced systemic neutropenia, rhCCSP has the potential, due to its bronchial specificity, to have a narrower compartment target and subsequently tighter regulation of neutrophilia.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary methods, tables and figures ERJ-02408-2018.Supplement
Acknowledgements
The authors are grateful to Valérie Scheuermann (PhyMedExp, INSERM U1046, University of Montpellier, Montpellier, France) for her technical assistance.
Footnotes
This article has supplementary material available from erj.ersjournals.com
This study is registered at www.ClinicalTrials.gov with identifier numbers NCT02354677 and NCT01947218. Individual de-identified participant data will be shared with participants. This data will include all the data collected in these trials Additional, related documents have been made available (study protocol, statistical analysis plan).
Author contributions: L. Knabe collected the clinical data, performed the experiments and statistical analyses, interpreted the data and wrote the manuscript. A. Petit, C. Vernisse and J. Charriot helped perform the experiments. M. Pugnière and C. Henriquet performed SPR experiments and provided their expertise. S. Sasorith performed 3D protein modelling and protein–protein docking analyses. N. Molinari provided statistical expertise and helped analyse the data. P. Chanez, J-P. Berthet, I. Vachier and E. Ahmed participated in the study design, interpreted the data and provided critical edits of the manuscript. C. Suehs helped in correcting the language and style. J. Charriot and J-P. Berthet recruited patients. A. Bourdin designed the study, recruited patients, collected the clinical data, performed statistical analyses, interpreted the data and provided critical edits of the manuscript. All authors provided critical review of the manuscript and approved its submission.
Conflict of interest: L. Knabe has nothing to disclose.
Conflict of interest: A. Petit has nothing to disclose.
Conflict of interest: C. Vernisse has nothing to disclose.
Conflict of interest: J. Charriot has nothing to disclose.
Conflict of interest: M. Pugnière has nothing to disclose.
Conflict of interest: C. Henriquet has nothing to disclose.
Conflict of interest: S. Sasorith has nothing to disclose.
Conflict of interest: N. Molinari has nothing to disclose.
Conflict of interest: P. Chanez has nothing to disclose.
Conflict of interest: J-P. Berthet has nothing to disclose.
Conflict of interest: C. Suehs has nothing to disclose.
Conflict of interest: I. Vachier has nothing to disclose.
Conflict of interest: E. Ahmed has nothing to disclose.
Conflict of interest: A. Bourdin has nothing to disclose.
Support statement: This study was supported by grants from the Agence Nationale de Recherche (ANR) Mucocil (ANR-13BSV5-0015-02), the Programme Hospitalier de Recherche Clinique (PHRC IR) AP-HM (2013-14) and the CHRU Montpellier (AOI 9244).
- Received May 29, 2018.
- Accepted April 8, 2019.
- Copyright ©ERS 2019
References










