





Abstract
Cathepsin S (CatS) is upregulated in the lungs of patients with cystic fibrosis (CF). However, its role in CF lung disease pathogenesis remains unclear.
In this study, β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice, a model of CF-like lung disease, were crossed with CatS null (CatS−/−) mice or treated with the CatS inhibitor VBY-999.
Levels of active CatS were elevated in the lungs of βENaC-Tg mice compared with wild-type (WT) littermates. CatS−/−βENaC-Tg mice exhibited decreased pulmonary inflammation, mucus obstruction and structural lung damage compared with βENaC-Tg mice. Pharmacological inhibition of CatS resulted in a significant decrease in pulmonary inflammation, lung damage and mucus plugging in the lungs of βENaC-Tg mice. In addition, instillation of CatS into the lungs of WT mice resulted in inflammation, lung remodelling and upregulation of mucin expression. Inhibition of the CatS target, protease-activated receptor 2 (PAR2), in βENaC-Tg mice resulted in a reduction in airway inflammation and mucin expression, indicating a role for this receptor in CatS-induced lung pathology.
Our data indicate an important role for CatS in the pathogenesis of CF-like lung disease mediated in part by PAR2 and highlight CatS as a therapeutic target.
Abstract
Cathepsin S is involved in inflammation, mucus production and lung tissue damage in a model of CF-like lung disease http://ow.ly/tHcm30nhlcX
Introduction
Cathepsin S (CatS) is expressed by professional and nonprofessional antigen-presenting cells, and is a crucial component in major histocompatibility complex class II antigen processing and presentation [1]. CatS is a potent elastinolytic and collagenolytic cysteine protease with the ability to maintain its enzymatic activity in more neutral pH environments [2, 3]. Previous work has demonstrated that levels of CatS are elevated in lungs of patients with cystic fibrosis (CF) lung disease [4–7] and chronic obstructive pulmonary disease (COPD) [8], and that CatS correlates significantly with a decline in lung function and increased pulmonary neutrophilic infiltration into the CF lung [7]. Furthermore, CatS possesses the ability to cleave and inactivate host defence proteins such as lactoferrin, β-defensins, LL-37, secretory leukoprotease inhibitor and surfactant protein A [4, 9–12], which may increase susceptibility to infection in patients with these muco-obstructive lung diseases.
However, the role of CatS in CF lung disease pathobiology has yet to be fully elucidated. In this study we employed a mouse model of CF-like lung disease to investigate the functionality of CatS in vivo. Overexpression of the β subunit of the epithelial Na+ channel (βENaC) results in airway surface dehydration and impaired mucus clearance, producing chronic lung disease characterised by airway mucus obstruction, inflammation and structural lung damage in mice [13]. Therefore, the βENaC-overexpressing transgenic (βENaC-Tg) mouse model shares many characteristics with human CF, and is a useful in vivo model to study the role and therapeutic potential of CatS in CF-like lung disease [14].
Following demonstration that elevated CatS levels and activity were a feature of the βENaC-Tg lung, we crossed βENaC-Tg mice onto a CatS null (CatS−/−) background and assessed airway inflammation, mucus obstruction and lung damage. To validate observations in this model we also investigated the effects of therapeutic targeting of CatS via a small-molecule inhibitor in the diseased lung. In addition, we instilled CatS directly into the lungs of wild-type (WT) mice to investigate effects on inflammatory cell recruitment, mucin production and lung remodelling. To further understand how CatS functions in the lung, we assessed the role of the protease-activated receptor 2 (PAR2), a target for CatS [15–20], in mediating the effects of CatS in βENaC-Tg mice.
Methods
Full details are available in the supplementary methods.
Abbreviated methods
All experimentation was carried out in accordance with the Animal (Scientific Procedures) Act 1986 and current guidelines approved by the Ethical Review Committee of Queen's University Belfast (Belfast, UK).
All mice used in this study were from a C57BL/6 background. This breeding programme generated four genotypes of mice for investigation: WT, CatS−/−, βENaC-Tg and CatS−/−βENaC-Tg mice. Offspring were genotyped by PCR as previously described [13, 21, 22] and mice were used from the newborn stage. Newborn βENaC-Tg mice and WT littermates were dosed once daily for 14 days via a subcutaneous injection of the CatS inhibitor VBY-999 (100 mg·kg−1; Virobay, Menlo Park, CA, USA), the selective PAR2 peptide antagonist FSLLRY-NH2 (4 mg·kg−1; R&D Systems, Abingdon, UK) or their respective vehicle controls, i.e. dextrose solution and sterile water. WT mice were intratracheally instilled with recombinant human CatS (rhCatS, 5 µg; Merck Millipore, Nottingham, UK). Bronchoalveolar lavage (BAL) fluid was collected for analysis of inflammatory markers and cell recruitment as previously described [13, 23]. Histological staining and morphometric assessments of airspace enlargement, alveolar wall destruction and airway mucus content were conducted as previously described [24–26].
Results
Upregulation of CatS in the lungs of βENaC-Tg mice
In order to validate the βENaC-Tg mouse model as a suitable model to test the effects of targeting CatS, we determined the status of CatS levels and activity in βENaC-Tg mice. We observed increased levels of CatS protein (figure 1a) and CatS activity (figure 1b) in BAL fluid from βENaC-Tg mice compared with healthy WT counterparts. In addition, we observed increased CatS staining in βENaC-Tg mouse lungs, which was primarily localised to macrophages, with some epithelial staining also present (figure 1c). These findings demonstrate that, like human CF, increased levels of active CatS are a feature of the βENaC-Tg mouse model.

Levels and activity of cathepsin S (CatS) are upregulated in the lungs of β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. WT: wild-type; RFU: relative fluorescence units; BAL: bronchoalveolar lavage. a) Western blot of CatS in BAL fluid from βENaC-Tg and WT mice. n=4 per group. b) CatS activity in BAL fluid from WT and βENaC-Tg mice was determined using Z-Phe-Arg-7-amido-4-methylcoumarin, hydrochloride substrate (pH 7.5). Results are presented as ΔRFU over time. n=10 per group. ***: p<0.001. c) Immunohistochemical staining for CatS on lung sections from WT and βENaC-Tg mice. Red arrows: macrophage CatS staining; yellow arrows: epithelial CatS staining.
Genetic ablation of CatS results in decreased pulmonary inflammation in βENaC-Tg mice
In order to investigate the role of CatS in the pathogenesis of CF-like lung disease, we first assessed pulmonary inflammation in CatS−/−βENaC-Tg, βENaC-Tg and control mice (CatS−/− and WT littermates). There was no significant change in BAL fluid total cell numbers in CatS−/− compared with WT control mice (figure 2a). As expected, βENaC-Tg mice demonstrated significantly increased cell infiltration compared with WT and CatS−/− mice (figure 2a). CatS−/−βENaC-Tg mice BAL total cell counts were significantly decreased compared with βENaC-Tg mice (figure 2a) as evidenced by a reduction in both macrophage (figure 2b) and neutrophil (figure 2c) cell populations. BAL fluid total protein concentrations were significantly higher in βENaC-Tg mice compared with control mice (figure 2d). However, total protein content was significantly reduced in CatS−/−βENaC-Tg mice compared with βENaC-Tg mice, suggesting a reduction in the severity of lung damage and inflammation (figure 2d). In addition, we observed significantly decreased levels of the chemokine KC (figure 2e) in BAL fluid of CatS−/−βENaC-Tg mice compared with βENaC-Tg mice.

Genetic ablation of cathepsin S (CatS) reduces airway inflammation in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. WT: wild-type; BAL: bronchoalveolar lavage. βENaC-Tg mice were crossed with CatS−/− mice to produce CatS−/−βENaC-Tg mice. BAL fluid was collected from WT, CatS−/−, βENaC-Tg and CatS−/−βENaC-Tg mice. BAL a) total cell, b) macrophage and c) neutrophil counts, and d) total protein concentration were quantified. n=7–10 per group. e) BAL fluid levels of chemokine KC were assessed by ELISA. n=5–7 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Genetic ablation of CatS results in decreased structural lung damage and mucus obstruction in the lungs of βENaC-Tg mice
Mean linear intercept length (Lm) is a measurement of distal airspace enlargement. Airway sections stained with haematoxylin and eosin (representative images shown in figure 3a) demonstrated no difference in Lm between CatS−/− and WT mice (figure 3b). As expected, a significant increase in Lm was evident in the lungs of βENaC-Tg mice compared with control mice (figure 3b). In comparison with βENaC-Tg mice, Lm was significantly reduced in CatS−/−βENaC-Tg mice (figure 3b), suggesting that increased CatS in the lungs of βENaC-Tg mice may play a role in distal airspace enlargement. In addition, the level of alveolar septal destruction, as assessed by measurement of the destructive index (DI), was significantly reduced in CatS−/−βENaC-Tg mice compared with βENaC-Tg mice, with no difference in DI measurements observed between WT and CatS−/− control mice (figure 3c). Given the levels of airspace enlargement and damage observed, these findings indicate that the augmented CatS levels found in βENaC-Tg may play an important role in mediating lung tissue damage and remodelling in CF-like lung disease. Histological staining and quantification of airway mucus content demonstrated a significant reduction in airway mucus obstruction in CatS−/−βENaC-Tg mice compared with βENaC-Tg mice (figure 3d and e). Overall, these findings suggest that CatS is implicated in the pathogenesis of chronic inflammation, lung tissue damage and the development of mucus plugging in CF-like lung disease.
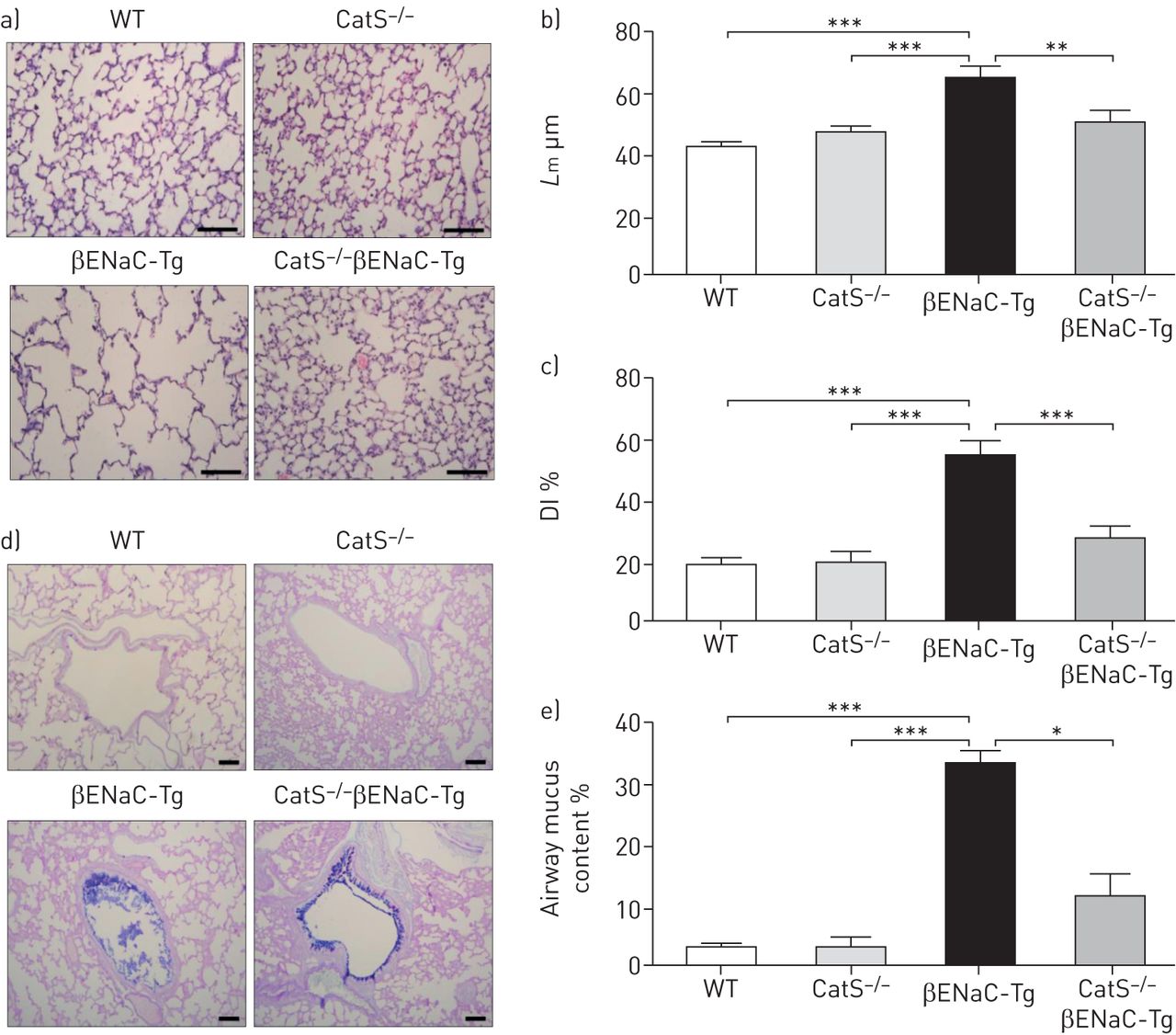
Lack of cathepsin S (CatS) in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice reduces airway mucus obstruction and lung tissue damage. WT: wild-type; HE: haematoxylin and eosin; Lm: mean linear intercept length; DI: destructive index. a) Representative images of lung sections from WT, CatS−/−, βENaC-Tg and CatS−/−βENaC-Tg mice stained with HE used to assess airway damage. Scale bar: 100 µm. b) Lm and c) DI measurements were assessed from the HE-stained lung sections. d) Representative images of lung sections stained with Alcian Blue–Periodic acid–Schiff. Scale bar: 100 µm. e) Airway mucus quantification expressed as percentage of the total airway containing mucus. n=7–10 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Pharmacological inhibition of CatS reduces airway inflammation in βENaC-Tg mice
CatS was targeted using a small-molecule inhibitor, VBY-999 [27], in βENaC-Tg mice. Inhibition of CatS alters invariant chain (Ii) processing and results in the accumulation of the 10 kDa intermediate (p10) [1, 22]. To determine whether VBY-999 was successfully targeting CatS in the lung, we assessed the accumulation of the Ii p10 fragment by Western blotting. The lungs of βENaC-Tg mice treated with VBY-999 exhibited an accumulation of Ii p10 levels compared with vehicle (dextrose)-treated βENaC-Tg mice (figure 4a). Initially, we observed that βENaC-Tg mice treated with VBY-999 from birth for a period of 14 days demonstrated a significant increase in survival compared with dextrose-treated mice (supplementary figure S1). βENaC-Tg mice treated with VBY-999 had significantly lower BAL total inflammatory cellular infiltrate compared with vehicle-treated βENaC-Tg mice (figure 4b). There was no effect on total cell numbers in the control mice regardless of treatments. VBY-999 treatment significantly decreased the number of BAL macrophages (figure 4c) and neutrophils (figure 4d) in βENaC-Tg mice. In addition, we observed significantly decreased levels of KC (figure 4e) in BAL fluid of VBY-999-treated βENaC-Tg mice compared with vehicle-treated βENaC-Tg mice. These data suggest that inhibition of CatS suppresses pulmonary inflammation in this mouse model of CF-like lung disease.

Pharmacological targeting of cathepsin S (CatS) leads to a reduction in pulmonary inflammation in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. Dex: dextrose; VBY: VBY-999; Ii: invariant chain; WT: wild-type; BAL: bronchoalveolar lavage. WT and βENaC-Tg mice were treated daily with the CatS inhibitor VBY (100 mg·kg−1) or vehicle control (Dex) for 14 days. a) Western blot of Ii p10 in lung homogenate from Dex- and VBY-treated βENaC-Tg mice. n=4 per group. BAL b) total cell, c) macrophage and d) neutrophil counts were quantified. n=10–15 per group. e) BAL fluid levels of chemokine KC were assessed by ELISA. n=9–15 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Inhibition of pulmonary CatS reduces the level of structural lung damage and lung protein content in βENaC-Tg mice
Protein levels were assessed by BCA (bicinchoninic acid) assay and VBY-999-treated βENaC-Tg mice had lower levels of total protein in BAL fluid compared with vehicle-treated βENaC-Tg mice (figure 5a). Lm and DI were measured using haematoxylin and eosin-stained lung sections from each group (figure 5b) as markers of airspace enlargement and alveolar destruction, respectively. There was a significant decrease in Lm in βENaC-Tg mice treated with VBY-999 compared with vehicle (figure 5c). Additionally, a significant decline in alveolar wall destruction was observed in VBY-999-treated βENaC-Tg mice compared with vehicle-treated βENaC-Tg mice (figure 5d). These data demonstrate that inhibition of CatS resulted in a reduction in airway damage in CF-like lung disease.

Cathepsin S (CatS) inhibition reduces airway tissue damage in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. WT: wild-type; Dex: dextrose; VBY: VBY-999; BAL: bronchoalveolar lavage; BCA: bicinchoninic acid; HE: haematoxylin and eosin; Lm: mean linear intercept length; DI: destructive index. WT and βENaC-Tg mice were treated daily with the CatS inhibitor VBY (100 mg·kg−1) or vehicle control (Dex) for 14 days. a) BAL fluid protein levels were quantified by BCA assay. n=7–8 per group. b) Representative images of lung sections stained with HE used to assess airway damage. Scale bar: 100 µm. c) Lm and d) DI measurements were assessed from the HE-stained lung sections. n=6–7 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Inhibition of CatS results in reduced airway mucus content in βENaC-Tg mice
Histological analyses demonstrated that VBY-999-treated βENaC-Tg mice presented with a significant decrease in airway mucus content compared with dextrose-treated βENaC-Tg mice (figure 6a and b). Given our previous findings of altered mucus content in CatS−/−βENaC-Tg mice (figure 3d and e), we investigated the expression of the mucins Muc5ac and Muc5b and the goblet cell marker Gob5 (figure 6c–e). Treatment of βENaC-Tg mice with VBY-999 from birth significantly reduced the expression levels of Muc5ac (figure 6c) and Gob5 (figure 6e) compared with dextrose-treated βENaC-Tg mice. In contrast, inhibition of CatS had no effect on the expression level of Muc5b (figure 6d).

Pharmacological targeting of cathepsin S (CatS) reduces airway mucus plugging and mucin gene expression in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. Dex: dextrose; VBY: VBY-999; WT: wild-type. WT and βENaC-Tg mice were treated daily with the CatS inhibitor VBY (100 mg·kg−1) or vehicle control (Dex) for 14 days. a) Representative images of lung sections from WT and βENaC-Tg mice treated with Dex or VBY stained with Alcian Blue–Periodic acid–Schiff to assess airway mucus content. Scale bar: 100 µm. b) Airway mucus quantification in WT and βENaC-Tg mice treated with Dex or VBY. Results are presented as percentage of the total airway area containing mucus. n=9–13 per group. Expression levels of c) Muc5ac, d) Muc5b and e) Gob5 in lung tissue were analysed by quantitative PCR. n=6–14 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Effect of CatS pulmonary instillation on lung inflammation and mucin production
Direct instillation of rhCatS into the lungs of WT mice induced significant recruitment of inflammatory cells (figure 7a), largely attributed to the influx in macrophages (figure 7b) and neutrophils (figure 7c). Although there was a significant increase in the expression levels of Muc5ac (figure 7d) and Gob5 (figure 7e) in the lungs of mice that received CatS, the expression level of Muc5b was unchanged (figure 7f). In addition, rhCatS administration was associated with goblet cell hyperplasia (figure 7g–i). KC levels in BAL fluid (figure 7j) were significantly increased following rhCatS instillation and evidence of lung damage was also observed with significantly increased levels of the elastin breakdown product, desmosine, detectable in the lungs of CatS-treated mice (figure 7k).

Cathepsin S (CatS) instillation induces inflammation and mucus in the lungs. rhCatS: recombinant human CatS; BM: basement membrane; WT: wild-type; BAL: bronchoalveolar lavage; AB-PAS: Alcian Blue–Periodic acid–Schiff. WT mice received a single intratracheal instillation of rhCatS (5 µg) or sodium acetate buffer control. After 24 h, BAL fluid was collected, and a) total cell, b) macrophage and c) neutrophil counts were quantified. Expression levels of d) Muc5ac, e) Gob5 and f) Muc5b in lung tissue were analysed by quantitative PCR. g) Representative images of lung sections stained with AB-PAS. Scale bar: 50 µm. h) Goblet and i) mucus-producing cells were quantified from AB-PAS-stained lung sections. Results are presented as cells·mm–1 BM. BAL fluid j) chemokine KC and k) desmosine levels were assessed by ELISA. n=4–9 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
CatS-induced lung inflammation and mucin expression is mediated in part via PAR2
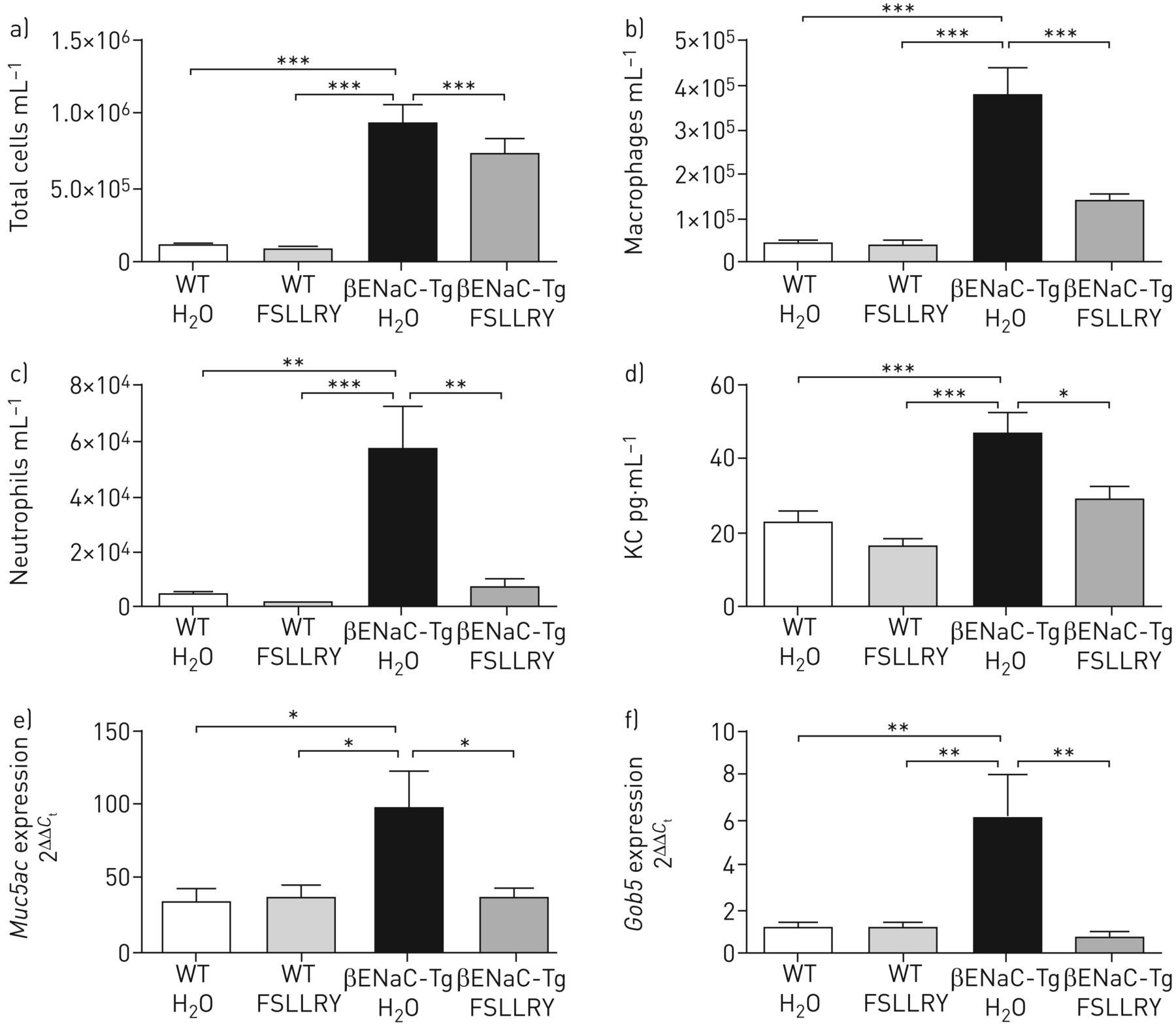
CatS has previously been shown to activate the receptor PAR2, leading to symptoms of inflammation, itch and pain, and has been associated with conditions such as atopic dermatitis and colitis as well as endothelial dysfunction associated with lupus nephritis and diabetes [15–20]. However, a role for PAR2 in CatS-mediated lung inflammation or mucin expression in vivo has not been described to date. Treatment of WT mice with a PAR2 antagonist (FSLLRY-NH2) reduced the infiltration of inflammatory cells into the lungs of rhCatS-treated mice (supplementary figure S2). Therefore, the impact of a PAR2 antagonist in the βENaC-Tg mouse lung was evaluated. Similar to genetic and pharmacological inhibition of CatS, blockade of PAR2 in βENaC-Tg mice led to a significant reduction in lung inflammation, with a decline in total inflammatory cell lung infiltrate (figure 8a), macrophages (figure 8b) and neutrophils (figure 8c) in the lungs of βENaC-Tg mice compared with vehicle (sterile water)-treated βENaC-Tg mice. In addition, BAL fluid KC levels were significantly decreased in PAR2 antagonist-treated βENaC-Tg mice compared with vehicle-treated βENaC-Tg mice (figure 8d). The expression levels of Muc5ac and Gob5 (figure 8e and f) were decreased in the lungs of PAR2 antagonist-treated βENaC-Tg mice compared with vehicle-treated βENaC-Tg mice. However, there was no reduction in the level of airspace enlargement or alveolar destruction as measured by Lm and DI, respectively (data not shown). Overall, these data indicate a significant role for PAR2 in CatS-mediated lung inflammation and mucus production in the lungs of βENaC-Tg mice.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Protease-activated receptor 2 (PAR2) inhibition reduces airway inflammation in β-epithelial Na+ channel-overexpressing transgenic (βENaC-Tg) mice. WT: wild-type; H2O: sterile water; FSLLRY: FSLLRY-NH2. WT and βENaC-Tg mice were treated daily with the PAR2 peptide antagonist FSLLRY (4 mg·kg−1) or vehicle control (H2O) for 14 days. Bronchoalveolar lavage a) total cell, b) macrophage and c) neutrophil counts, and d) chemokine KC levels were quantified. n=6–11 per group. Expression levels of e) Muc5ac and f) Gob5 in lung tissue were analysed by quantitative PCR. n=9 per group. *: p<0.05; **: p<0.01; ***: p<0.001.
Discussion
CatS is elevated in a number of muco-inflammatory conditions like CF lung disease and COPD, and is viewed as a potential therapeutic target for various diseases (recently reviewed in [28, 29]). We have previously shown that increased CatS activity in the CF lung is associated with a decline in lung function [7]. In addition, pulmonary CatS levels and activity significantly correlated with neutrophilic infiltration in the CF lung [7], substantiating the hypothesis that CatS may play a role in pulmonary neutrophil recruitment [30], which is associated with CF lung disease. In the current study, we show that elevated CatS is also a key feature of the pulmonary disease of βENaC-Tg mice, a model of CF-like lung disease. The βENaC-Tg mouse provides a relevant model for investigating CF lung disease pathogenesis, particularly the interactions between ion transport, airway surface liquid and mucociliary clearance [13, 14]. However, because CF transmembrane conductance regulator (CFTR) function is normal in the βENaC-Tg mouse lung [13], this model does not allow the study of pathophysiological changes associated directly with mutant CFTR, such as cellular dysfunction and mucus composition. Knockdown of CatS by pharmacological (VBY-999 inhibitor) or genetic methods (CatS−/−) in the βENaC-Tg model resulted in a significant decline in both lung macrophage and neutrophil cell populations, providing further evidence of a role for CatS in mediating pulmonary inflammation [30–32].
Previous work in βENaC-Tg mice demonstrated that genetic knockdown of neutrophil elastase and matrix metalloproteinase 12 significantly reduced structural lung damage, but had no effect on mucus obstruction and pulmonary mortality [33, 34]. In this study, pharmacological inhibition and genetic ablation of CatS not only led to reduced airway structural damage and pulmonary inflammation, but also ameliorated airway mucus plugging and improved survival rates of neonatal mice. These data indicate the involvement of CatS in inflammatory cell recruitment, lung tissue damage and the development of mucus plugging. In agreement, we have also shown that direct administration of CatS to the lungs of WT mice induced neutrophil recruitment and increased KC levels in the lungs of these mice. Pulmonary CatS instillation resulted in significantly increased expression of the mucin Muc5ac and Gob5, a marker associated with goblet cell hyperplasia [35, 36], as well as the number of goblet cells and mucus-producing cells, although no effect on Muc5b expression was observed. These results indicate a role for CatS-mediated lung inflammation in the CF lung, and suggest that CatS may contribute to airway mucus plugging via goblet cell hyperplasia and increased mucus production.
These effects may be due, in part, to the activation of PAR2 by CatS, which has previously been demonstrated in other organ systems outside of the lung [15–20]. Research to date suggests that PAR2 activation is associated with inflammation, leukocyte recruitment and mucin expression [37–40]. In this study we have shown that antagonism of PAR2 led to a reduction in the inflammatory cell infiltrate in the lungs of mice exposed to CatS as well as in the βENaC-Tg lung. In addition, reductions in BAL fluid KC levels and expression levels of Muc5ac and Gob5 were also observed in PAR2 antagonist-treated βENaC-Tg mice. In agreement with previous reports, our data suggest that CatS may exert its effects on pulmonary inflammation and mucus plugging, at least in part, via PAR2. In contrast, while knockdown or inhibition of CatS reduced lung tissue destruction in βENaC-Tg mice, we did not observe a reduction in lung damage (Lm or DI) in PAR2 antagonist-treated βENaC-Tg mice, suggesting that CatS may be acting via other pathways to elicit lung damage. For example, we have shown that direct instillation of CatS into the WT mouse lung can rapidly induce (within 24 h) desmosine levels in BAL fluid, indicating that the effects of CatS may also be mediated directly on lung tissue as well as via PAR2.
In summary, our investigations show that CatS is elevated in an in vivo model of CF-like lung disease, and may promote pulmonary cell infiltration, mucus obstruction and structural lung damage associated with chronic lung diseases such as CF. These results suggest that CatS may play a vital role in CF-like lung disease pathogenesis, which ultimately leads to the loss of tissue structural integrity and lung function. As previously mentioned, CatS possesses the ability to cleave and inactivate host antimicrobial proteins [4, 9–12], which may increase susceptibility to infection in patients with muco-obstructive lung diseases such as CF and COPD. However, further work is needed to investigate the impact of elevated CatS activity and the effects of pulmonary CatS inhibition on Pseudomonas infection in the context of chronic lung disease. Our results indicate that CatS may be a promising therapeutic target to halt the progression of airway inflammation, mucus obstruction and pulmonary damage associated with CF lung disease.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material ERJ-01523-2018_Supplement
Footnotes
This article has supplementary material available from erj.ersjournals.com
Conflict of interest: D.M. Small has nothing to disclose.
Conflict of interest: R.R. Brown has nothing to disclose.
Conflict of interest: D.F. Doherty has nothing to disclose.
Conflict of interest: A. Abladey has nothing to disclose.
Conflict of interest: Z. Zhou-Suckow has nothing to disclose.
Conflict of interest: R.J. Delaney has nothing to disclose.
Conflict of interest: L. Kerrigan has nothing to disclose.
Conflict of interest: C.M. Dougan has nothing to disclose.
Conflict of interest: K.S. Borensztajn has nothing to disclose.
Conflict of interest: L. Holsinger is an employee of Virobay.
Conflict of interest: R. Booth has a patent US 7,547,701 issued.
Conflict of interest: C.J. Scott is a consultant for Fusion Antibodies PLC, outside the submitted work; and is named inventor on various patents on antibodies to cathepsin S for treatment of cancer, many of which have now lapsed as not a current research direction for the company (Fusion Antibodies PLC).
Conflict of interest: G. López-Campos has nothing to disclose.
Conflict of interest: J.S. Elborn reports personal fees for advisory board work from Bayer, grants and personal fees for advisory board work from Horizion, during the conduct of the study; personal fees for advisory board work from Chiesi and Polyphor, outside the submitted work.
Conflict of interest: M.A. Mall reports grants from German Federal Ministry of Education and Research (contract numbers 82DZL00401 and 82DZL004A1), during the conduct of the study; personal fees for advisory board and consultancy work from Spyryx Biosciences, Boehringer Ingelheim and Polyphor, personal fees for advisory board work from ProQR, PTC Pharmaceuticals, Arrowhead and Pro Axis, personal fees for consultancy and lecturing from Bayer, personal fees for consultancy from Enterprise Therapeutics and Sterna Biologicals, personal fees for advisory board work, consultancy and lecturing from Vertex Pharmaceuticals, outside the submitted work; and has a patent on the Scnn1b-transgenic mouse with royalties paid.
Conflict of interest: S. Weldon reports grants from Randox and Pfizer UK, outside the submitted work.
Conflict of interest: C.C. Taggart reports personal fees for consultancy from Albumedix, and grants from Randox and Pfizer UK, outside the submitted work.
Support statement: Supported by the Cystic Fibrosis Foundation (TAGGAR12I0, WELDON15G0, WELDON18G0), EU FP7 (CFMATTERS; grant 603038), Engineering and Physical Sciences Research Council (studentships to A. Abladey and L. Kerrigan), QUB start-up funds (S. Weldon), Medical Research Council Confidence in Concept Programme (S. Weldon, C.C. Taggart), Dept for the Economy (C.C. Taggart), and German Federal Ministry of Education and Research (82DZL004A1 to M.A. Mall). Funding information for this article has been deposited with the Crossref Funder Registry.
- Received August 10, 2018.
- Accepted December 27, 2018.
- Copyright ©ERS 2019
References










