





Abstract
Intracellular Ca2+-binding S100A8/A9 proteins gain novel functions when released during inflammation. The exact outcome of their extracellular function depends on the local tissue environment in which they are released; both anti-inflammatory and pro-inflammatory responses are described, modulating the immune system by binding Toll-like receptor (TLR)-4 or the receptor for advanced glycation end-products (RAGE). However, the contribution of the proteins in the pathophysiology of chronic rhinosinusitis (CRS) remains unclear.
Homomeric S100A8 and S100A9, and heteromeric S100A8/A9 proteins were evaluated in CRS with/without nasal polyps (CRSw/sNP) and controls. Functional responses were assessed in polyp tissue stimulated with S100 proteins in the presence of TLR-4 and RAGE blocking antibodies.
S100A8, S100A9 and S100A8/A9 protein levels were significantly higher in CRSwNP patients, showing increased deposition on extracellular matrix (ECM) structures of CRSwNP tissue in contrast to CRSsNP and controls. In the presence of Staphylococcus aureus, S100A8/A9 is released from neutrophils and from the ECM. Extracellular S100A8 and S100A9 proteins induced increased levels of diverse inflammatory mediators via TLR-4 engagement.
The inflammatory/remodelling characteristics of CRSwNP specifically allow increased retention of S100A8, S100A9 and S100A8/A9 proteins in the ECM of CRSwNP tissue. Upon release, homodimeric proteins act as a local danger signal inducing inflammatory mediators, predominantly via TLR-4 activation.
Abstract
Deposition of S100A8/A9 proteins on ECM of CRSwNP tissue released from neutrophils induces pro-inflammatory response http://ow.ly/Q2sV7
Introduction
Chronic rhinosinusitis with nasal polyps (CRSwNP) is a prevalent chronic inflammatory condition of the upper airways which is often accompanied by comorbid asthma, associated with a substantially impaired quality of life and reduced workplace productivity [1]. A diverse spectrum of factors including viral and bacterial insults [2, 3], together with epithelial barrier malfunctions and tissue remodelling [4–6] could be implicated in the aetiology of CRSwNP. However, despite massive research efforts to unveil the pathophysiology, the exact reason for a lack of resolution still remains poorly understood and appears to be multifactorial [7].
A novel set of molecules that could be implicated in sustaining the inflammatory reaction may be found within the host itself. Indeed, in addition to mediators of inflammation originating externally, some endogenous intracellular molecules from the host can be released into the extracellular space where they gain functions that are distinct from those during normal physiology, modulating inflammatory responses under pathological conditions. These molecules are collectively classified as damage-associated molecular patterns (DAMPs) [6]. S100A8 and S100A9 proteins possess all of the characteristics required to be included in the group of DAMPs [8, 9]. S100A8 and S100A9 proteins are two of the 24 members of the multifunctional S100 family of cytoplasmic EF-hand helix-loop-helix Ca2+-binding proteins and are abundantly expressed in myeloid cells. Both S100A8 and S100A9 form noncovalently associated homo- and heterodimers [10, 11]. The heterodimeric complex is the predominantly occurring form and considered relevant for normal physiology [12, 13]. However, recent results have also shown that homodimers of S100A8 and S100A9 exhibit strong pro-inflammatory activities in various mouse models [14, 15].
Extracellularly, while both the homodimeric and heterodimeric proteins are described to possess chemotactic properties [13, 16, 17], the individual homodimeric S100A8 and S100A9, and the heterodimeric S100A8/A9 proteins often have distinct functions [18]. It has been shown that the antimicrobial activity of extracellular S100 proteins is only exhibited via divalent metal cation sequestration by the heterodimeric form S100A8/A9 [19, 20]. Conversely, the pro-inflammatory effects of these S100 proteins per se can be exclusively assigned to the homodimeric S100A8 and S100A9 forms [14]. S100A8 and S100A9 proteins are described to modulate the innate immune system by binding the receptor for advanced glycation end-products (RAGE) [21] and Toll-like receptor (TLR)-4 [14].
S100A8 and S100A9 proteins have been associated with human diseases such as rheumatoid arthritis, juvenile idiopathic arthritis, inflammatory bowel disease, cystic fibrosis and several auto-inflammatory diseases [9, 22]. Although many functions have been identified and proposed for these proteins, their exact roles in various biological processes still remain to be defined. In particular, the implication of S100A8/A9 proteins and their potential mechanism of action in the pathophysiology of CRSwNP is not well investigated.
Methods
Patients
Patient clinical data can be found in table 1. More detailed information regarding the sampling of tissues, sera and nasal secretions can be found in the online supplementary material.
Patient clinical data and other characteristics
Tissue homogenates
Frozen tissue samples were homogenised by means of mechanical disruption as described in the online supplementary material.
S100A8, S100A9 and S100A8/A9 ELISA
S100A8, S100A9 and S100A8/A9 protein levels were measured in tissue homogenates, serum, nasal secretions, and tissue pellets/supernatants from tissue-cube experiments (see below) by means of commercially available ELISA kits (BMA Biomedicals, Augst, Switzerland). Tissue homogenates were prepared from nasal polyp tissue of CRSwNP patients, ethmoidal mucosal tissue from CRS without nasal polyps (CRSsNP) patients and from inferior turbinates of control subjects. Human ex-vivo tissue-cube fragment stimulation assays were performed on nasal polyps from CRSwNP patients. More detailed technical information regarding the ELISA kits is provided in the online supplementary material.
S100A8, S100A9 and S100A8/A9 immunohistochemistry
Paraffin and cryo-sections of human sinonasal tissues from controls (inferior turbinates), CRSwNP patients (inferior turbinates and nasal polyps) and CRSsNP patients (ethmoidal mucosa) were stained for S100A8, S100A9, S100A8/A9 and/or myeloperoxidase (MPO), as reported in detail in the online supplementary material.
S100A8 and S100A9 gene expression analysis
RNA extraction, first-strand cDNA synthesis and real-time PCR amplification of S100A8 and S100A9 were performed on inferior turbinates of control subjects and on nasal polyps of CRSwNP patients as described in detail in the online supplementary material.
Human sinonasal ex vivo tissue-cube fragment stimulation assay
Fresh human tissues were processed as described previously [23]. For a detailed description on the specific methodology applied in this study, see the online supplementary material.
Assessment of inflammatory mediators
Tissue-cube supernatants and/or tissue homogenates were assayed for interleukin (IL)-1β, IL-5, IL-6, IL-8, IL-10, IL-17 and tumour necrosis factor (TNF)-α levels by means of Luminex xMAP technology using a Fluorokine MAP Multiplex Human Cytokine Panel A kit (R&D Systems, Minneapolis, MN, USA) on a Bio-PlexTM 200 Array Reader (Bio-Rad, Hercules, CA, USA). Eosinophilic cationic protein (ECP) levels were measured by UniCAP (Phadia, Uppsala, Sweden). A commercially available ELISA kit was used to measure the levels of transforming growth factor (TGF)-β1 (R&D Systems); acid was added during the ELISA procedure, resulting in physicochemical activation of latent TGF-β. Total TGF-β concentrations are reported, including both active and latent forms.
Assessment of cell death
Lactate dehydrogenase (LDH) levels were measured by means of a commercial cytotoxicity detection kit (Roche, Basel, Switzerland) according to the manufacturer's guidelines, using an LDH standard from rabbit muscle (Roche).
Statistical analysis
The data obtained from the two patient groups were analysed by a nonparametric Mann–Whitney U-test, while the data derived from human ex vivo tissue-cube fragment stimulation assays were compared by means of a nonparametric Kruskal–Wallis test followed by Dunn's multiple comparison test. Categorical variables were analysed by means of Fisher's exact test. Correlation analysis was performed by the Spearman rank-order method yielding a Spearman rho (rs) coefficient. A p-value ≤0.05 was considered to be statistically significant (GraphPad, San Diego, CA, USA).
Results
S100A8, S100A9 and S100A8/A9 protein levels in human sinonasal tissue
ELISA data show tissue levels of S100A8 (figure 1a), S100A9 (figure 1b) and S100A8/A9 proteins (figure 1c) to be significantly higher in nasal polyp tissue from CRSwNP patients compared with control inferior turbinate. Tissue S100A8/A9 levels highly correlated with tissue IL-8 and MPO levels in both controls (Spearman rs=0.8074, p<0.0001 and rs=0.941, p<0.0001, respectively; figure 2a and b) and CRSwNP patients (Spearman rs=0.7778, p<0.0001 and rs=0.875, p<0.0001, respectively; figure 2c and d). The correlations were absent for homodimeric forms (data not shown).

Sinonasal tissue levels of a) S100A8, b) S100A9 and c) S100A8/A9 proteins in controls and chronic rhinosinusitis with nasal polyps (CRSwNP) patients. Values presented as scatter plots showing the median as a line. *: p<0.05; ***: p<0.001. The corresponding patient clinical data can be found in table 1 and table E1 (in the online supplementary material).
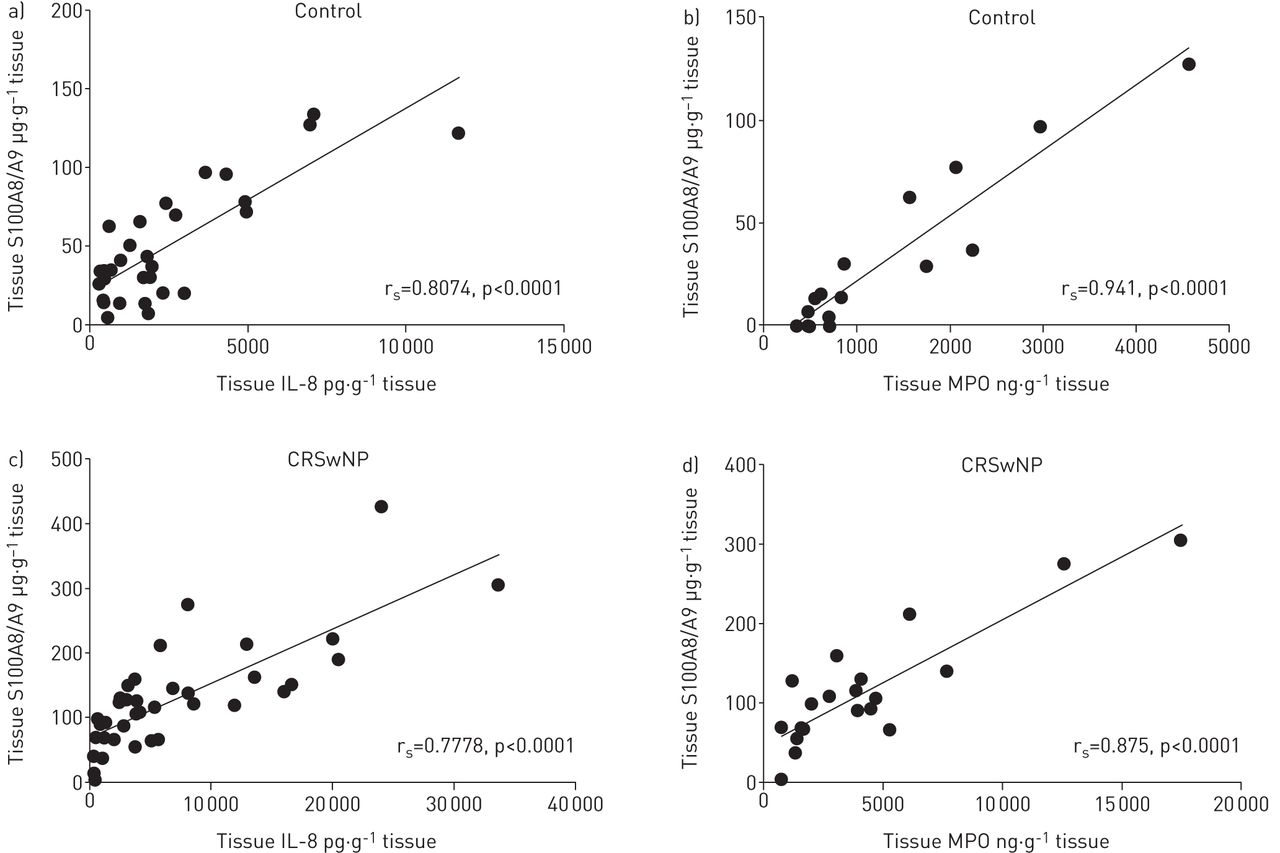
Correlation of tissue S100A8/A9 protein levels with tissue protein levels of interleukin (IL)-8 and myeloperoxidase (MPO) in a, b) controls (Spearman rs=0.8074, p<0.0001, n=30 and rs=0.941, p<0.0001, n=17, respectively) and c, d) chronic rhinosinusitis with nasal polyps (CRSwNP) patients (Spearman rs=0.7778, p<0.0001, n=38 and rs=0.875, p<0.0001, n=22, respectively).
None of the other clinical parameters or inflammatory mediators reported in table 1 showed any relationship to the S100A8, S100A9 and S100A8/A9 protein levels.
S100A8, S100A9 and S100A8/A9 immunohistochemistry
Acetone-fixed cryo-sections showed no or very locally limited S100A9 immunoreactivity (clone IDCP1) in control inferior turbinates of nondiseased subjects (figure 3a), contrasting with a very pronounced S100A9 immunoreactivity around the basement membrane, blood vessels, and in the deeper ECM structures of inferior turbinate (figure 3b) and nasal polyp (figure 3c) tissue from CRSwNP patients (figure 3d).

Immunostaining for S100A9 in inferior turbinate (IT) from a, g) control subjects, and b) inferior turbinate (IT) and c, e, f, h, i) nasal polyps (NP) from chronic rhinosinusitis with nasal polyps (CRSwNP). d) Semi-quantitative levels of immunostaining shown in panels a–c. **: p<0.01 (n=6). Double staining with antibodies against h) myeloperoxidase (MPO) and i) MPO together with S100A9 confirmed that the major source of these S100 proteins is neutrophils. Staining in panels a-c was performed on acetone-fixed cryo-sections; staining in panels e−i was performed on formalin-fixed paraffin sections. Antibody clone IDCP1 was used for all stainings. Scale bars=100 μm.
In formalin-fixed paraffin sections from CRSwNP tissue, the same antibody against S100A9 stained monocytes (figure 3e) and neutrophils (figure 3f), which were more abundantly present in CRSwNP (figure 3e and f) tissues compared with control tissue (figure 3g). Double staining with antibodies against MPO (figure 3h) and MPO together with S100A9 (figure 3i) in human polyp tissue confirmed that the major source of S100A8/A9 proteins is neutrophils.
In both paraffin and cryo-sections, similar results were obtained with the monoclonal antibody against S100A8 (clone S13.67) and with polyclonal antibodies against S100A8 and S100A9 (generated by T. Vogl) (data not shown).
Gene expression for S100A8 and S100A9
The relative levels of gene expression for S100A8 and S100A9 did not differ between inferior turbinates from controls and nasal polyps from CRSwNP patients (figure E3 in the online supplementary material).
Functional effect of S100A8, S100A9 and S100A8/A9 proteins on inflammatory cytokine release ex vivo
Stimulation of nasal polyp tissue fragments with 1 (data not shown) and 5 μg·mL−1 recombinant S100A8 and S100A9 proteins resulted in a concentration-dependent increase in IL-1β (p<0.05 and nonsignificant, respectively), TNF-α (both p<0.001), IL-10 (both p<0.001) and IL-17 (p<0.001 and p<0.05, respectively) levels in the tissue culture supernatants (figure 4a–d), while the levels of IL-5 and IL-8 (figure 4e and f), and IL-6, TGF-β and ECP (figure E4 in the online supplementary material) were not affected. S100A8/A9 did not alter the release of any of the measured mediators (figure 4a–f).

Effect of 5 µg·mL−1 S100A8 (A8), S100A9 (A9) and S100A8/A9 (A8/9) proteins on release of a) interleukin (IL)-1β, b) tumour necrosis factor (TNF)-α, c) IL-10, d) IL-17, e) IL-5 and f) IL-8 proteins from human nasal polyp tissue in an ex vivo tissue assay. Data are presented as box-and-whisker plots showing minimum and maximum values, lower and upper quartiles, and the median (n=6). *: p<0.05; **: p<0.01.
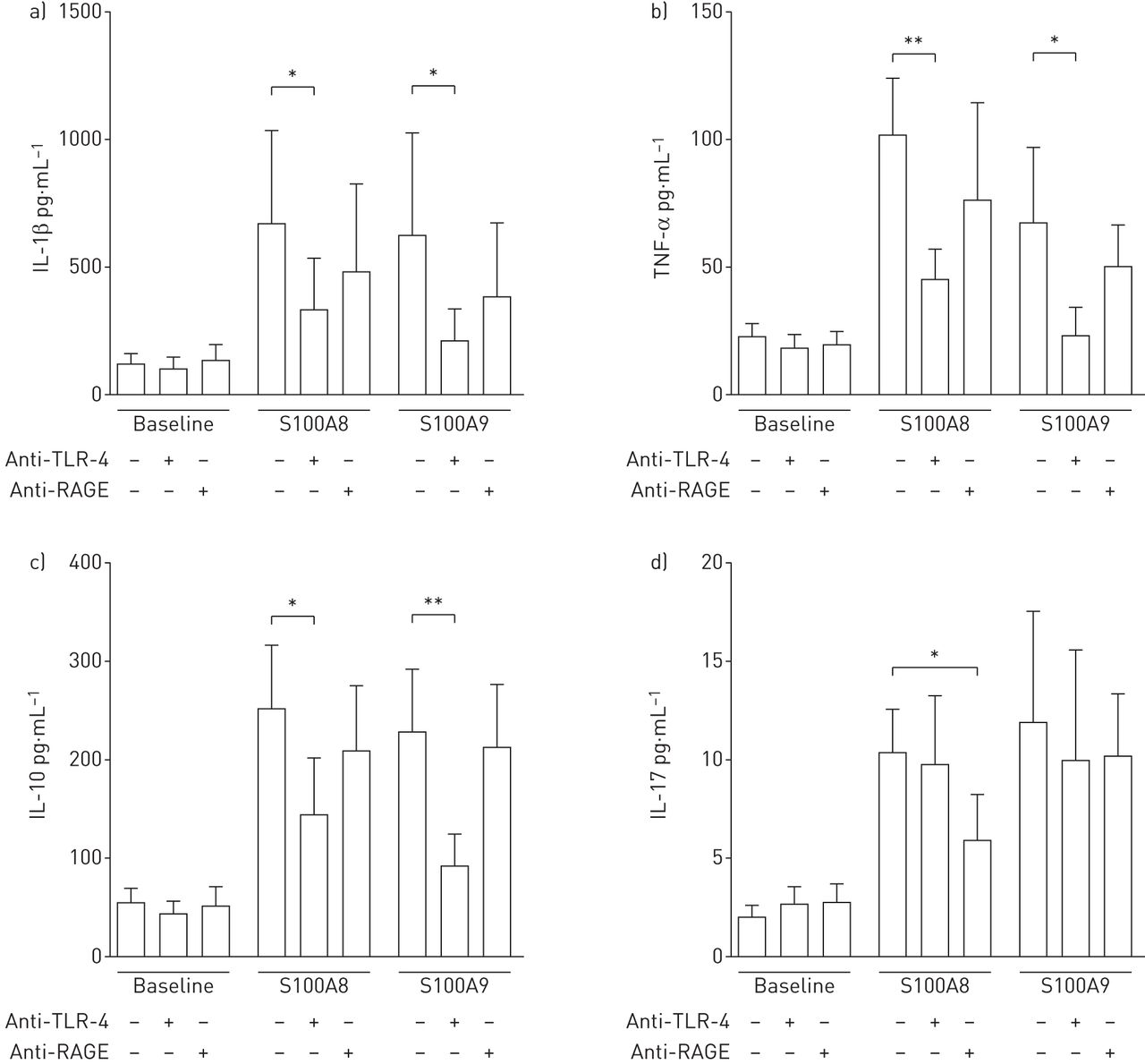
The increases in IL-1β, TNF-α and IL-10 levels induced by S100A8 and S100A9 were significantly reduced in the presence of anti-TLR-4 antibodies, while functional blocking of the RAGE receptor had no effect (figure 5a–c). In contrast, the levels of IL-17 induced by S100A8 were significantly lowered in the presence of RAGE blocking antibodies, while TLR-4 blocking had no effect. Neither RAGE nor TLR-4 blocking could affect the levels of IL-17 induced by S100A9 (figure 5d). Neither the isotype control for TLR-4 nor the isotype control for RAGE had any effect (data not shown).

Effect of Toll-like receptor (TLR)-4 (2 µg·mL−1) and the receptor for advanced glycation end-products (RAGE) (10 µg·mL−1) blocking antibodies on the responses induced by exogenous 5 µg·mL−1 S100A8 and S100A9 proteins on the release of a) interleukin (IL)-1β, b) tumour necrosis factor (TNF)-α, c) IL-10 and d) IL-17 proteins from human nasal polyp tissue in an ex vivo tissue assay. Data are presented as mean±sem (n=6). *: p<0.05; **: p<0.01.
Effect of Staphylococcus aureus on the release of S100 proteins ex vivo
Under baseline conditions, S100A8/A9 time-dependently increased in the tissue-cube supernatants of the human nasal polyp tissue-cube fragment stimulation assay measured between 1 and 6 h, and stabilised after 6 h, as assessed at 24 h (1 h, 1530±610 ng·mL−1; 6 h, 2010±750 ng·mL−1; 24 h, 2000±620 ng·mL−1). Neutralisation of the IL-8 bioactivity did not affect the magnitude of S100A8/A9 release assessed at 24 h compared with control conditions (1910±310 versus 2110±350 ng·mL−1; n=5).
In the presence of S. aureus, but not heat-inactivated S. aureus, a clear-cut increase in heterodimeric S100A8/A9 levels was observed in the tissue-cube supernatants at 24 h, while the levels within the tissue-cube pellets were significantly decreased (figure 6a). A similar release of S100A8/A9 proteins by S. aureus was observed from peripheral blood-derived granulocytes (figure E7 in the online supplementary material) and to a lesser extent from peripheral blood mononuclear cells (0.222±0.037 µg·mL−1 at baseline versus 2.037±0.127 µg·mL−1 in the presence of 105 CFU S. aureus; n=5, p<0.05). Homodimeric S100A9 proteins could not be measured in the supernatants of the conditions described above, but S100A9 was present in the tissue-cube pellets and was significantly reduced by S. aureus but not by heat-inactivated S. aureus (figure 6b). Homodimeric S100A8 could not be measured in all of the samples described above (data not shown). Cell death could be excluded in these experiments as levels of LDH in the assay supernatants were not increased in the presence of S. aureus compared with baseline conditions (figures E7 and E8 in the online supplementary material). S. aureus-derived staphylococcal enterotoxin B and lipoteichoic acid had no effect on the levels of S100A8/A9 (data not shown).

Effect of exogenously added 105 CFU Staphylococcus aureus on the release of a) S100A8/A9 and b) S100A9 proteins from the human nasal polyp tissue into the supernatants in an ex vivo tissue assay. S100A8 was not detectable in the samples (data not shown). Data are presented as mean±sem (n=6–8). HI: heat-inactivated. *: p<0.05; **: p<0.01.
The increased release of S100A8/A9 in the tissue-cube supernatants by S. aureus was significantly reduced by the tubulin polymerisation inhibitor demecolcine and the actin polymerisation inhibitor cytochalasin B, while the de novo protein synthesis inhibitor cycloheximide and the endoplasmic reticulum/Golgi complex transport inhibitor brefeldin A had no effect (figure E9 in the online supplementary material).
Discussion
S100A8/A9 proteins comprise up to 45% of all cytosolic proteins in neutrophils and ∼5% of all monocyte/macrophage cytosolic proteins [24, 25]. Indeed, a clear-cut infiltration of neutrophils that stained intensely positive for S100A8/A9 could be observed in paraffin sections of nasal polyp tissue. Moreover, we found S100A8 and S100A9 protein deposition in the extracellular matrix (ECM) of nasal polyps and inferior turbinates of CRSwNP patients, but not in CRSsNP patients or in control subjects. S100A8/A9 proteins were previously reported to interact with heparan sulfate proteoglycans [26] such as perlecan, agrin and collagen XVIII, which are present on ECM components, explaining our immunohistochemical stainings for S100A8 and S100A9. S100A8- and S100A9-immunoreactive vessels evidently do not synthesise these proteins themselves, but bind the S100A8/A9 proteins that appear to have been released by myeloid cells. It is suggested that when monocytes and neutrophils adhere to vascular endothelium, they release S100A8/A9 proteins onto the associated endothelium [27]. Our data, showing a correlation between tissue levels of IL-8 and S100A8/A9, while both proteins do not have a direct effect on their reciprocal release, can be brought in agreement with this mechanism of action. Indeed, IL-8 produced by activated epithelium or resident macrophages is known to be a strong chemoattractant for neutrophils which constitutively contain large amounts of S100A8/A9 proteins in their cytosol and consequently bring in these proteins in the CRSwNP tissue (figure 7). This influx of S100A8/A9 protein-containing cells and the observed protein deposition in the ECM can equally explain why the relative levels of gene expression for S100A8 and S100A9 do not differ between the tissue of controls and CRSwNP patients, whereas proteins levels do differ.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proposed mechanism of action of S100 proteins contributing to the pathophysiology of chronic rhinosinusitis with nasal polyps as described in detail in the Discussion section. A8: S100A8; A9: S100A9; DAMP: damage-associated molecular pattern; ECM: extracellular matrix; IL; interleukin; mRAGE: membrane-bound receptor for advanced glycation end-products; PAMP: pathogen-associated molecular pattern; S. aureus: Staphylococcus aureus; TLR; Toll-like receptor; TNF: tumour necrosis factor.
Although some S100A8/A9-containing myeloid cells can be found in nondiseased control tissues, even being abundantly present in CRSsNP, no deposition of S100A8/A9 proteins in the ECM structure could be observed in control and CRSsNP tissue, suggesting facilitated binding of the S100 proteins to the ECM under the specific pathology of CRSwNP. Indeed, tissue remodelling in CRSwNP [4] might change the ECM structure composition and thereby affect the ability of the S100 proteins to attach. In addition to increased retention in the ECM, alterations in the magnitude or release mechanisms of S100 proteins under inflammatory conditions can contribute equally to the increased levels of extracellular S100A8/A9 proteins. Indeed, our data show that S. aureus, a frequent coloniser of human upper airway mucosa with an increased prevalence in CRSwNP patients [4], can induce the release of S100A8/A9 from inflamed tissue of CRSwNP patients and from peripheral blood-derived granulocytes and peripheral blood mononuclear cells. S. aureus is known to be implicated in the pathophysiology and severity of CRSwNP disease progression and the rate of developing comorbid asthma [28, 29]. Corroborated by the decreased levels of S100A8/A9 proteins measured in the CRSwNP tissue pellets, S. aureus induces a mechanism that liberates the cytosolic fraction of S100A8/A9 proteins from granulocytes. In view of the effect of brefeldin A, demecolcine and cytochalasin B, the increased release of S100A8/A9 from neutrophils can be attributed to occur via a nonclassical secretory pathway during phagocytic cell migration and phagocytosis in which microtubule and actin-related pathways play important functions (figure 7). Phagocytosis-induced release of antimicrobial [20] heterodimeric S100A8/A9 proteins by neutrophils in CRSwNP can comprise a local environment [19, 30] optimised to combat S. aureus, a major contributor in the pathophysiology of CRSwNP [2].
In contrast to the direct antimicrobial effect of heterodimeric S100A8/A9 proteins [19, 20, 30], homodimeric S100A8 and S100A9 proteins also show the typical parameters of pro-inflammatory DAMPs [6]. In human CRSwNP tissue, we report that homodimeric S100A8 and S100A9 proteins induce a clear-cut increase in several inflammatory cytokines which are predominantly TLR-4-mediated [14], whereas RAGE [31] plays only a minor role (figure 7). In contrast, the heterodimeric S100A8/A9 protein per se yielded no inflammatory responses in our experiments. In addition to the release from the cytosol of myeloid cells into the extracellular environment, another prerequisite for the S100 proteins to induce the observed pro-inflammatory reactions thus relates to the availability of the homodimeric S100A8 and S100A9 proteins at the site of inflammation. Despite the fact that neutrophils exclusively release the heterodimeric S100A8/A9 form [32, 33], as evidenced from our ex vivo tissue and granulocyte model, both homodimeric S100A8 and S100A9 protein forms are present in the upper airway tissue, being significantly higher in CRSwNP patients. The mechanisms leading to the presence of homomeric proteins in the tissue of patients is consequently not due to a direct release from the neutrophils.
It can be speculated that ECM-associated heterodimeric S100A8/A9 proteins can be cleaved into the monomeric forms by a certain trigger that is present in the local tissue environment and subsequently be released from the matrix by the same or another trigger. When attached to the matrix, the monomers, which are normally not stable, might be protected from degradation by the local tissue environment and/or form more stable homodimers via association with an unbound homologous monomer [34]. The binding of S100A8/A9 proteins to the ECM of CRSwNP tissue can be specifically attributed to the binding capacity of the S100A9 part of the dimer as it has been shown that S100A9 but not S100A8 binds to heparan sulfate proteoglycans [26]. When the ECM-bound S100A8/A9 protein dissociates under influence of a specific trigger (see below), the S100A9 proteins could remain intercalated in the ECM while the S100A8 monomers are released from the matrix, subsequently forming homodimers. Although the heterodimer is the preferential form, when one of the two monomers is present in excess over the other, homodimers are formed [32]. These homodimers are much more stable than the monomeric proteins, allowing them to induce their pro-inflammatory responses before being degraded, as they, in contrast to the heterodimeric S100A8/A9 proteins, are still prone to proteolytic degradation over time, which is especially true for S100A8 [35–37]. Indeed, also in the upper airways, the levels of S100A8 proteins from fresh homogenates were substantially lower when compared with the levels of S100A9 and S100A8/A9. When transferred to ex vivo culture conditions for 24 h, endogenous homodimeric S100A8 proteins are not measurable anymore, whereas S100A8/A9 and S100A9 proteins were still clearly detectable in the tissue pellets. A second trigger might induce ectodomain shedding to subsequently induce the release of the ECM-associated homodimeric S100A9 proteins.
The increased prevalence of S. aureus in CRSwNP could bring into play such a set of triggers that might be able to split ECM-associated S100A8/A9 proteins into their monomeric forms because S. aureus is known to produce a diverse set of extracellular proteases, including serine proteases, cysteine proteases and metalloenzymes [38], which are often insensitive to human protease inhibitors, making them particularly aggressive. Moreover, S. aureus is also known to induce ectodomain shedding of host cell surface molecules to enhance their virulence [39], which might subsequently result in the release of the ECM-associated S100A9 proteins, explaining our ex vivo results showing S. aureus to reduce the levels of S100A9 proteins present in CRSwNP tissues (figure 7).
Our results suggest that homodimeric S100A9 proteins are protected from degradation during longer periods when attached to the ECM. Indeed, while S. aureus is able to reduce the levels of S100A9 proteins present in CRSwNP tissues, these proteins cannot be detected in the supernatants after 24 h ex vivo, indicating that they are indeed not 100% unsusceptible to degradation and are broken down over time when released from the ECM. A similar suggestion for protection against breakdown of the homodimeric S100A8 and S100A9 proteins by a local tissue environment could be made based on the phenotyping results of S100A9 knockout mice [37]. Similarly to what we propose for the human upper airway, this indicates that changes in local inflammatory status can affect protein stability and that in more peripherally located cells, this stability is jeopardised, probably by the lack of a protective environment. Hence the importance of studying these proteins under conditions resembling natural human physiology and pathology as close as possible, as we aim to do by means of our human sinonasal tissue model.
In conclusion, we report that S100A8, S100A9 and S100A8/A9 proteins are increased in sinonasal tissue of CRSwNP patients as a consequence of increased deposition on ECM structures in addition to the influx of S100A8/A9-containing inflammatory cells. Although neutrophils are more abundantly present in CRSsNP, this increased deposition in the tissue's ECM is only observed in CRSwNP, indicating that the specific inflammatory/remodelling milieu characteristic of CRSwNP allows the S100 proteins to attach. Heterodimeric S100A8/A9 proteins are released from neutrophils during phagocytosis of S. aureus, while the latter is able to release homodimeric S100A8 and S100A9 proteins from the ECM of CRSwNP tissue. Upon release, the homodimeric proteins show the potential to act as a danger signal by the release of inflammatory mediators, predominantly via TLR-4 activation in human CRSwNP tissue.
Footnotes
This article has supplementary material available from erj.ersjournals.com
Support statement: This work was supported by funding from The Research Foundation Flanders (FWO) (research project no. G.0641.10) to K. Van Crombruggen, a Special Research Fund BOF (no. 01J01113) to K. Van Crombruggen, the Interuniversity Attraction Poles Program-Belgian Science Policy (P7/30) to C. Bachert and the German Research Foundation (DFG) (CRC1009 B8) to T. Vogl. Funding information for this article has been deposited with FundRef.
Conflict of interest: None declared.
- Received January 28, 2015.
- Accepted July 4, 2015.
- Copyright ©ERS 2016
References










