





Abstract
Recently, a locus centred on rs9273349 in the HLA-DQ region emerged from genome-wide association studies of adult-onset asthma. We aimed to further investigate the role of human leukocyte antigen (HLA) class II in adult-onset asthma and a possible interaction with occupational exposures.
We imputed classical HLA-II alleles from 7579 single-nucleotide polymorphisms in 6025 subjects (1202 with adult-onset asthma) from European cohorts: ECRHS, SAPALDIA, EGEA and B58C, and from surveys of bakers and agricultural workers. Based on an asthma-specific job–exposure matrix, 2629 subjects had ever been exposed to high molecular weight (HMW) allergens. We explored associations between 23 common HLA-II alleles and adult-onset asthma, and tested for gene–environment interaction with occupational exposure to HMW allergens. Interaction was also tested for rs9273349.
Marginal associations of classical HLA-II alleles and adult-onset asthma were not statistically significant. Interaction was detected between the DPB1*03:01 allele and exposure to HMW allergens (p = 0.009), in particular to latex (p = 0.01). In the unexposed group, the DPB1*03:01 allele was associated with adult-onset asthma (OR 0.67, 95%CI 0.53–0.86). HMW allergen exposures did not modify the association of rs9273349 with adult-onset asthma.
Common classical HLA-II alleles were not marginally associated with adult-onset asthma. The association of latex exposure and adult-onset asthma may be modified by DPB1*03:01.
Abstract
In an analysis involving >6000 European adults, classical HLA-II alleles were not associated with adult-onset asthma http://ow.ly/xTdFm
Introduction
The highly polymorphic human leukocyte antigen (HLA) class II molecules (DR, DQ and DP) are expressed on the surface of antigen-presenting cells, where they present peptides of exogenous origin to T-cell receptors on CD4+ cells. Because of the pivotal role of HLA-II molecules in the recognition of allergen-derived peptides, HLA alleles have been extensively studied in hypothesis-driven candidate gene association studies for allergic sensitisation and asthma [1, 2]. HLA is also the most evaluated genetic risk factor for occupational asthma. Although results were inconsistent, a role of the HLA-DQB1 locus in occupational asthma aetiology is plausible [3].
Evidence from several independent genome-wide association studies (GWAS) lend further support to the involvement of HLA-II loci in asthma, allergic sensitisation and increased total IgE level [4–11]. The GABRIEL consortium carried out the largest asthma GWAS to date, by genotyping 10 365 asthma patients and 16 110 controls recruited from 23 studies [4]. A locus centred on rs9273349 in the HLA-DQ region showed strong evidence of association in the overall sample (p = 7×10−14; OR for C allele 1.18). This was the only single-nucleotide polymorphism (SNP) with genome-wide significance for asthma with onset at 16 years of age or older (p = 4×10−8; OR 1.26). Interestingly, a more pronounced effect of rs9273349 was observed in later-onset asthma than in childhood-onset asthma, although the difference was not significant. A recent GWAS replicated the association of this SNP with asthma and also reported notably stronger associations in the adult compared with the childhood population [6].
It has been suggested that investigating gene–environment interactions may help to understand associations of HLA with asthma that were identified by GWAS [12]. Indeed, the strength of the association of HLA variants with asthma could depend on concomitant environmental exposures. Considering individuals’ age of asthma onset can be instrumental in revealing gene–environment interactions [13, 14]. The age-dependent effects that were observed in GWAS may provide clues to the role of HLA, because late-onset asthma may have different underlying environmental risk factors than childhood asthma. In adults, occupational exposures are an important cause of incident asthma, with a population attributable risk of around 15% [15–17]. Investigating the effect of gene–environment interactions between HLA alleles and occupational exposures on asthma risk might reveal mechanisms mediated by specific HLA alleles and triggered by occupational sensitisers such as latex, flour or animal proteins.
The aim of the present study is to further investigate the role of HLA-II in adult-onset asthma in a meta-analysis of population-based and workforce-based studies. We imputed classical HLA-II alleles from 7579 SNPs, allowing an evaluation of association between HLA-II alleles and adult-onset asthma. In addition, we explored whether occupational exposure to high molecular weight (HMW) allergens modifies the effect of imputed HLA-II alleles and the rs9273349 SNP in the HLA-DQ region that emerged from the GABRIEL study on adult-onset asthma.
Methods
Population
We studied all European cohorts that were included in GABRIEL with detailed data available on occupational history: EGEA [18, 19], SAPALDIA [20, 21] and ECRHS [15, 22] (referred to as the “ESE cohorts”) [23], the British 1958 birth cohort (B58C) [16, 24], and industrial cohorts of Dutch and Danish agricultural workers (AGRI) [25], and Dutch bakery workers (BAKERY) [26]. We included nested asthma case–control samples (SAPALDIA, ECRHS, AGRI and BAKERY) as previously described [4], and all subjects from EGEA and B58C with data available on genome-wide genotyping, occupational history and adult-onset asthma. Adult-onset asthma was defined as ever having had asthma with an onset ≥16 years [4]. Subjects who had never had asthma were used as the reference group. Ethical approval was obtained for each cohort from the appropriate institutional ethics committee. Detailed cohort descriptions are given in the online supplementary material.
Genotyping
Genome-wide genotyping was conducted using the Human610 quad array panel (Illumina, San Diego, CA, USA) [4]. More details on genotyping of B58C are given in the online supplementary material. We investigated whether the association of the GABRIEL main signal (rs9273349) with adult-onset asthma was modified by occupational exposure. For completeness regarding GABRIEL HLA-II results, we further selected a subset of 18 tag SNPs using Haploview (Broad Institute, Cambridge, MA, USA) [27], and tested for gene–environment interaction with occupational exposure to HMW allergens (online supplementary material).
Imputation of classical HLA-II alleles
For all cohorts in the present analysis, we used 7579 SNPs of the extended major histocompatibility complex (MHC) (20–40 Mbp on chromosome 6) to impute classical HLA types for class II loci at four-digit resolution. Imputations were performed using BEAGLE software (University of Washington, Seattle, WA, USA) on the basis of haplotype patterns in 2767 unrelated individuals of European descent (this reference panel includes B58C subjects), as described previously [28]. Concordance between imputed data and genotyping (Dynal RELI SSO assays; Invitrogen, Carslbad, CA, USA) [29] was assessed for DRB1 and DQB1 alleles in the 2310 B58C subjects included in the present paper who were not part of the reference panel for the BEAGLE imputations (online supplementary table E1). Imputation quality was generally high for common alleles (r2>0.8). Pairwise linkage disequilibrium measures (D′ and r2) between classical HLA-II alleles and rs9273349 and the other 18 SNPs were estimated using Haploview (tables E2 and E3) [27].
Occupational exposure to HMW allergens
Occupational exposure assessment is described in detail in the online supplementary material. Briefly, in all cohorts, occupations were linked to an asthma-specific job–exposure matrix with a local expert re-evaluation step [19]. We only considered seven HMW allergens high-risk substances (namely animal proteins, fish or shellfish, mites, latex, flour, other plant antigens, and enzymes) that may cause asthma through a similar, predominantly IgE-associated mechanism. Subjects who were ever exposed to any of the seven HMW allergens were classified as “exposed”. The “unexposed” reference group consisted of subjects who were never exposed to any of the high-risk or low-risk jobs.
Statistical analysis
The ESE cohorts were pooled to increase statistical power and to avoid convergence problems, as was done before with EGEA and ECRHS data [30]. Raw data from B58C were not available for pooled analysis. We tested association of 23 common classical HLA-II alleles with allele frequency >5% in each population with adult-onset asthma, assuming an additive genetic model. In ESE cohorts, population admixture was taken into account by including principal components for within-Europe diversity as covariates in the association analysis. Genetic effects were estimated in each cohort separately, stratified by exposure status. The overall (exposure-adjusted) gene effect was estimated by standard fixed-effects meta-analysis of ESE exposed, ESE unexposed, B58C exposed and B58C unexposed subjects. To evaluate gene–environment interaction, results in exposed and unexposed subjects from ESE and B58C were combined through fixed-effects meta-analysis to obtain odds ratios in the exposed and unexposed groups. Both the standard 1 degree of freedom (df) test of gene–environment interaction and the 2 df joint test of marginal genetic effects and gene–environment interaction were performed, the latter being generally considered to have greater power than the marginal test when the genetic effect is only expressed in exposed individuals [31, 32]. Additional analyses were performed by including the industrial cohorts in the meta-analysis of HMW allergen exposed workers, and performing the joint test and test for interaction. The industrial cohorts were not included to evaluate the marginal genetic effect because all subjects in these cohorts were exposed, so the interaction effect cannot be assumed to be independent of the exposed-only odds ratio. We did not adjust for multiple testing because of the prior evidence of HLA-II involvement in several GWAS. Instead, we used a p-value threshold of <0.01 for the test of marginal association, joint test and test for interaction. More details are given in the online supplementary material.
Results
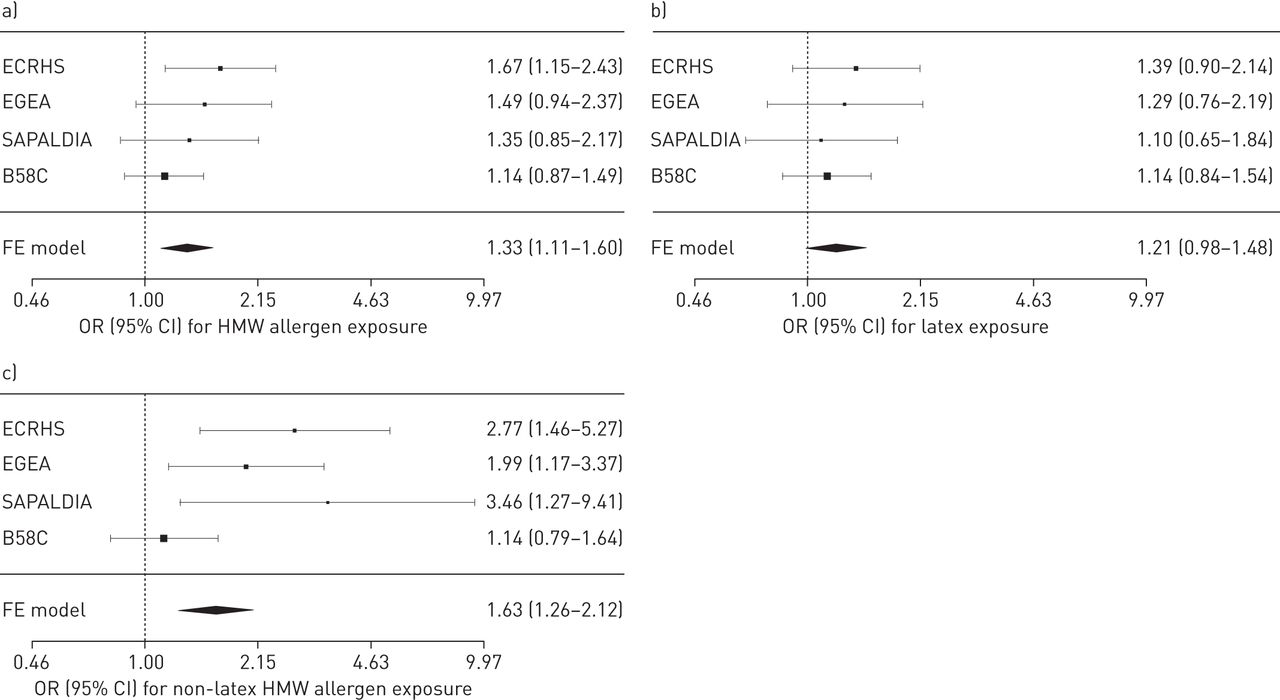
Descriptive characteristics of the HMW allergen exposed and unexposed study subjects are summarised in table 1. Subjects with childhood-onset asthma (n = 685) or missing age of onset (n = 59) were excluded from data analysis, leaving data from 6025 subjects for analysis. Latex was the predominant occupational exposure among subjects from the general population cohorts who were classified as HMW allergen exposed (from 64% exposed in B58C to 84% in SAPALDIA). Occupational exposure to HMW allergens was positively associated with adult-onset asthma in each of the four general population cohorts, with a fixed-effects summary odds ratio of 1.33 (95% CI 1.11–1.60) (fig. 1).
{kind=link}
Forest plot of odds ratios and 95% confidence intervals for the association of occupational exposure to a) high molecular weight (HMW) allergens, b) latex and c) HMW allergens other than latex with adult-onset asthma in ECRHS, EGEA, SAPALDIA and B58C. The summary odds ratio was calculated using a fixed-effects meta-analysis (FE model).
table 2 shows results from association tests of 23 imputed classical HLA-II alleles, rs9273349 C/T, and adult-onset asthma in the ESE and B58C cohorts. Marginal associations of HLA-II alleles were not statistically significant (p>0.01) but rs9273349 was, as expected, significantly associated with adult-onset asthma (OR for T allele 0.82, p = 0.002).
Association between rs9273349 and adult-onset asthma was similar in exposed and unexposed individuals (test for gene–environment interaction p = 0.95). For DPB1*03:01, the joint test was statistically significant (p = 0.007, interaction p = 0.03). In unexposed individuals, DPB1*03:01 was associated with a decreased risk of adult-onset asthma (OR 0.67, p = 0.002), but not in exposed subjects (OR 1.08, p = 0.64). Study-specific estimates showed that the interaction was driven by the ESE cohorts (OR for exposed 1.34, p = 0.20; OR for unexposed 0.62, p = 0.0009). In ESE subjects carrying at least one DPB1*03:01 allele, the proportion of subjects with adult-onset asthma was higher in those exposed to occupational HMW allergens (35.6%) and lower in the unexposed (15.0%), compared with the same groups in ESE noncarriers (28.4% and 22.2% in exposed and unexposed, respectively; fig. E1). Thus, in these cohorts, a stronger association between adult-onset asthma and occupational HMW allergens was detected in carriers of at least one DPB1*03:01 allele (OR 3.13, 95% CI 1.87–5.23) than in noncarriers (OR 1.39, 95% CI 1.08–1.81). In B58C, DPB1*03:01 was not associated with asthma, whichever the exposure status (OR exposed 0.84, p = 0.48; OR unexposed 0.89, p = 0.62). Associations of DPB1*03:01 and rs9273349 with adult-onset asthma were independent: the effect of each genetic polymorphism did not change when both were jointly included in a multiple regression model.
In the HMW allergen exposed industrial cohorts (meta-analysis of AGRI and BAKERY), DPB1*03:01 was not associated with asthma (table 3). However, the test for gene–environment interaction between DPB1*03:01 and HMW exposure was more significant after including the industrial cohorts together with the population-based cohorts in the analysis (p = 0.009). The association of allele DQB1*06:02 with asthma (p = 0.007) was restricted to the industrial cohorts, with a similar effect in agricultural workers (OR 0.72, p = 0.05) and bakery workers (OR 0.65, p = 0.07).
As the majority of exposed subjects from the general population cohorts were exposed to latex, an additional sensitivity analysis was performed in ESE and B58C, by testing for gene–environment interaction in two mutually exclusive exposed groups: “ever being exposed to latex” and “ever being exposed to HMW allergens, but never to latex”, with unexposed as a reference group (tables E4 and E5). The gene–environment interaction between DPB1*03:01 and overall HMW exposure was only observed in latex exposed versus unexposed subjects (interaction p = 0.01). As a result, in ESE subjects carrying at least one DPB1*03:01 allele, a strong association between latex exposure and adult-onset asthma was observed (OR 3.12, 95% CI 1.73–5.65).
Association between the 18 tag SNPs in the HLA-II region and adult-onset asthma was similar in occupationally exposed and unexposed individuals (interaction p>0.05; table E6).
Discussion
In a meta-analysis of European cohorts, we did not find evidence to suggest that common HLA-II alleles are associated with the risk of adult-onset asthma. However, when analyses were stratified by occupational HMW allergen exposure, the DPB1*03:01 allele appeared to be negatively associated with adult-onset asthma in unexposed subjects, and an interaction was found between DPB1*03:01 and exposure to latex. DQB1*06:02 showed a consistent inverse association with asthma in cohorts of bakery workers and agricultural workers, but not in the HMW allergen exposed subjects from the general population studies.
We showed that rs9273349, the SNP in the HLA-DQ region that emerged from the GABRIEL GWAS [4], influences adult-onset asthma to an equal extent in both HMW allergen exposed and occupationally unexposed individuals. Thus, no evidence was found to suggest that occupational allergen exposures might be involved in the association of rs9273349, or any of the other 18 selected tag SNPs in the HLA-II region, and adult-onset asthma. rs9273349 was significantly associated with adult-onset asthma in combined ESE and B58C studies (OR 0.82, p = 0.002). We also computed this association in the other GABRIEL adult cohorts that were not included in the present study because detailed occupational history information was not available, showing a similar result (OR 0.80, p = 0.001). We have shown that rs9273349 is in a region of extended linkage disequilibrium (as indicated by D′) that includes DRB1, DQA1 and DQB1, but not DPA1 or DPB1. However, none of the classical HLA-II alleles were strongly correlated with rs9273349 (r2 = 0.00–0.32; tables E2 and E3), and the rs9273349 association signal was not explained by the classical HLA-II alleles in multiple regression models. Despite the well described role of HLA-II molecules in allergen recognition, it is as yet unclear what the association of rs9273349 with adult-onset asthma might mean. It could be speculated that this SNP might influence the level of expression of HLA molecules. Alternatively, non-HLA genes could play a role. The MHC is one of the most complicated regions of the human genome, with extensive linkage disequilibrium that extends across hundreds of HLA and non-HLA genes. Linkage and association between HLA variants and asthma may in part be due to association between nearby non-HLA genes (in linkage disequilibrium with HLA) and asthma, e.g. TNFA [33]. The latter explanation would imply a role for different mechanisms, which may also explain why we did not find an interaction between rs9273349 and occupational allergen exposure.
Our study population was large enough to reveal moderate associations between classical HLA-II alleles and adult-onset asthma, and the complete absence of marginal associations is probably not due to a lack of statistical power. When taking occupational exposure into account, carrying DPB1*03:01 appeared to protect against adult-onset asthma among unexposed individuals. However, in ESE subjects carrying the DPB1*03:01 allele, a strong positive association between exposure to latex and adult-onset asthma was observed (OR >3). This finding suggests that DPB1*03:01 may confer an increased risk of developing asthma in adulthood, but only for those with specific workplace exposures. We can only speculate on the mechanism behind an opposite effect among the unexposed. A better protection against respiratory infections or the development of allergy to other antigens may be involved. A German case–control study investigated the role of HLA-DQ and HLA-DR (but not HLA-DP) among 269 health care workers with latex allergy and 90 nonatopic control subjects [34]. DRB1*04/DQB1*03:02 haplotype was associated with an increased risk of IgE to a major latex allergen (Hev b 6.02) [34]. We also found positive, but not significant, associations of DRB1*04:01 and DQB1*03:02 with adult-onset asthma among our 1123 latex-exposed workers. Only a few other studies investigated HLA alleles in relation to asthma or allergic outcomes among workers exposed to HMW allergens, namely laboratory animal allergens [35], cow dander [36] and fish food [37]. DQB1*06:02, which was here associated with asthma both in bakery workers and agricultural workers, did not emerge from these other studies.
Studying gene–environment interaction in large-scale studies has some inherent limitations. In populations sampled from the general population (or asthma patients and their families as in EGEA), there is a large heterogeneity in occupations and related exposures. The majority of workers who were classified as “exposed to HMW agents” are latex-exposed healthcare professionals who are typically exposed to other agents, such as cleaning products, that may cause asthma through a non-IgE mediated mechanism [38]. Despite the heterogeneity in exposures, associations between occupational exposure to HMW allergens and adult-onset asthma were comparable across cohorts and showed a significantly increased risk, which was similar in magnitude to earlier published associations [15, 16, 19]. In the present sample of B58C, the association of HMW exposure and asthma appeared to be weaker than the earlier published association in a larger sample of B58C [16]. This difference appeared to be due to chance variation, as the odds ratios did not differ significantly between the B58C samples with and without genotyping available.
The proportion of males among exposed subjects was lower in SAPALDIA (12%) and ECRHS (14%) than in EGEA (29%) and B58C (33%). In the ESE exposed group, association between DPB1*03:01 and adult-onset asthma was more pronounced in males (OR 2.71, p = 0.055) than in females (OR 1.18, p = 0.52), but interaction was not significant (p = 0.18) due to small sample size. This association did not differ between unexposed males and females in ESE. The workforce-based studies also differed from the population-based cohorts in terms of exposures, phenotype and proportion of male subjects. Therefore, we performed separate analyses in population-based cohorts only and in all exposed subjects. Also within the workforce-based cohorts, heterogeneity in source, nature and levels of allergen exposures exists. Bakery workers can be exposed to allergenic enzymes and flour allergens, but also to cleaning products, whereas agricultural workers (e.g. grain farmers and dairy farmers) may be exposed to allergens from animal danders, storage mites, grain and flour. Various other exposures in the agricultural environment, such as endotoxins and disinfectants, are also associated with (nonallergic) asthma. The association of DQB1*06:02 with asthma among bakers and farmers may provide further explanations why some of these workers develop respiratory health problems, while others with similar exposure levels remain healthy [39].
In this large-scale meta-analysis, we were not able to study specific IgE to occupational allergens (e.g. latex, wheat and α-amylase) or clinically assessed occupational asthma as a phenotype. Instead, we used adult-onset asthma, which was consistently associated with HMW allergen exposure (fig. 1) and with rs9273349 in GWAS. We aimed to further investigate this HLA signal in GWAS and, therefore, we decided not to include other asthma phenotypes or atopic sensitisation in the present study. Although a substantial part of adult-onset asthma is attributable to occupational exposures, other risk factors, triggers and comorbid conditions such as smoking, female sex hormones, respiratory infections and obesity are also associated with the development of asthma in adulthood [40]. The relevance of HLA-II to most of these determinants is, however, less obvious.
In conclusion, imputed common HLA-II alleles were not associated per se with adult-onset asthma in a meta-analysis of European cohorts. However, when taking occupational allergen exposures into account, results showed a negative association between DPB1*03:01 and adult-asthma in the unexposed, and suggested a gene–environment interaction between the DPB1*03:01 allele and exposure to latex. Furthermore, DQB1*06:02 was associated with asthma among bakery workers and agricultural workers, suggesting a role for specific occupational allergens. Occupational HMW allergen exposures did not modify the association between rs9273349, the SNP in the HLA-DQ region that emerged from the GABRIEL GWAS, and adult-onset asthma.
Acknowledgments
The author’s affiliations are as follows. Lidwien A.M. Smit: Division of Environmental Epidemiology, Institute for Risk Assessment Sciences (IRAS), Utrecht, The Netherlands; David P. Strachan: Division of Population Health Sciences and Education, St George’s University of London, London, UK; Roel Vermeulen: Division of Environmental Epidemiology, Institute for Risk Assessment Sciences (IRAS), and Julius Center for Health Sciences and Primary Care, University Medical Center Utrecht, Utrecht, The Netherlands; Paul I.W. de Bakker: Julius Center for Health Sciences and Primary Care, and Institute for Molecular Medicine, University Medical Center Utrecht, Utrecht, The Netherlands; Florence Demenais: INSERM, U946, and Université Paris Diderot, Sorbonne Paris Cité, Institut Universitaire d’Hématologie, Paris, France; Orianne Dumas: INSERM, Centre for Research in Epidemiology and Population Health (CESP), U1018, Respiratory and Environmental Epidemiology Team, and Université Paris Sud 11, UMRS 1018, Villejuif, France; Anne-Elie Carsin: Centre for Research in Environmental Epidemiology (CREAL), Municipal Institute of Medical Research (IMIM-Hospital del Mar), and CIBER Epidemiologia y Salud Pública (CIBERESP), Barcelona, Spain; Paul Cullinan: Dept of Occupational and Environmental Medicine, National Heart and Lung Institute, Imperial College, London, UK; Ivan Curjuric: Swiss Tropical and Public Health Institute, and University of Basel, Basel, Switzerland; Rebecca E. Ghosh: Respiratory Epidemiology and Public Health Group, National Heart and Lung Institute, and Small Area Health Statistics Unit, MRC-PHE Centre for Environment and Health, Imperial College, London, UK; Dick Heederik: Division of Environmental Epidemiology, Institute for Risk Assessment Sciences (IRAS), Utrecht, The Netherlands; Medea Imboden: Swiss Tropical and Public Health Institute, and University of Basel, Basel, Switzerland; Deborah Jarvis: Respiratory Epidemiology and Public Health Group, National Heart and Lung Institute, Imperial College, London, UK; Mark Lathrop: Commissariat à l’Energie Atomique, Institut de Génomique, Centre National de Génotypage (CNG), Evry, France, and McGill University, Montreal, QC, Canada; Nicole Le Moual: INSERM, Centre for Research in Epidemiology and Population Health (CESP), U1018, Respiratory and Environmental Epidemiology Team, and Université Paris Sud 11, UMRS 1018, Villejuif, France; Amar Mehta: Swiss Tropical and Public Health Institute, and University of Basel, Basel, Switzerland, and Harvard School of Public Health, Dept of Environmental Health, Boston, MA, USA; David Miedinger: University of Basel, Basel, and Swiss Accident Insurance Fund (Suva), Lucerne, Switzerland; Torben Sigsgaard: Dept of Environmental and Occupational Medicine, Aarhus University, Aarhus, Denmark; Valérie Siroux: INSERM, U823, and Université Joseph Fourier, Grenoble, France; David Vernez: Institute for Work and Health, Lausanne, Switzerland; Jan Paul Zock: Centre for Research in Environmental Epidemiology (CREAL), CIBER Epidemiologia y Salud Pública (CIBERESP), and Universitat Pompeu Fabra (UPF), Barcelona, Spain; Francine Kauffmann: INSERM, Centre for Research in Epidemiology and Population Health (CESP), U1018, Respiratory and Environmental Epidemiology Team, and Université Paris Sud 11, UMRS 1018, Villejuif, France; Nicole Probst-Hensch: Swiss Tropical and Public Health Institute, and University of Basel, Basel, Switzerland; Manolis Kogevinas: Centre for Research in Environmental Epidemiology (CREAL), Municipal Institute of Medical Research (IMIM-Hospital del Mar), and CIBER Epidemiologia y Salud Pública (CIBERESP), Barcelona, Spain, and National School of Public Health, Athens, Greece; Emmanuelle Bouzigon: INSERM, U946, and Université Paris Diderot, Sorbonne Paris Cité, Institut Universitaire d’Hématologie, Paris, France.
We are grateful to all study members and staff involved in data collections in each cohort. For the 1958 Birth Cohort, the asthma-specific job–exposure matrix was applied with an expert re-evaluation step by UK experts in occupational lung disease (Paul Cullinan, David Fishwick (Centre for Workplace Health, Sheffield, UK), Jennifer Hoyle (North Manchester General Hospital, Manchester, UK) and Chris Warburton (North Manchester General Hospital, Manchester, UK)). Further acknowledgements are given in the online supplementary material.
Footnotes
This article has supplementary material available from www.erj.ersjournals.com
Support statement: This gene–environment interaction analysis was supported by IAGO ANR-2010-PRSP-003, and by the Netherlands Organisation for Scientific Research (NWO)/French Ministry of Foreign and European affairs Van Gogh programme for French–Dutch cooperation. Genotyping of the ESE and industrial cohorts, and part of B58C was funded by the GABRIEL asthma genetic consortium supported by a contract from the European Commission (018996) and grants from the French Ministry of Research, the Wellcome Trust (WT084703MA), Asthma UK and The COLT Foundation. For B58C, Asthma UK provided funding for the asthma-specific job–exposure matrix coding of occupational exposures and investigation of their associations with asthma, and the COLT Foundation provided funding for extension of this work to look at other allergic outcomes. Further sources of funding for the participating cohorts are given in the online supplementary material. None of the funders had any role in study design, data collection and analysis, or preparation of the manuscript.
Conflict of interest: Disclosures can be found alongside the online version of this article at www.erj.ersjournals.com
- Received April 9, 2014.
- Accepted June 9, 2014.
- ©ERS 2014
References










