





Abstract
In preterm human infants, briefly elevated concentrations of oxygen are associated with a prolonged increase in blood chemokine concentrations and the development of bronchopulmonary dysplasia (BPD). Caffeine given to preterm infants for the prevention or treatment of apnoea has been shown to reduce the rate of BPD.
We tested the hypotheses that infant rats exposed to a combination of caffeine and hyperoxia would be less susceptible to lung injury than those exposed to hyperoxia alone and that caffeine decreases the pulmonary tissue expression of chemokines and leukocyte influx following hyperoxia.
Using 6-day-old rat pups, we demonstrated that 24 h of 80% oxygen exposure caused pulmonary recruitment of neutrophils and macrophages. High levels of oxygen upregulated the expression of: the CXC chemokines, cytokine-induced neutrophil chemoattractant-1 and macrophage inflammatory protein-2; the CC-chemokine monocyte chemoattractant protein-1; the pro-inflammatory cytokines tumour necrosis factor-α and interleukin-6, as measured by realtime PCR after the administration of caffeine (10 mg·kg−1 body weight); and attenuated chemokine and cytokine upregulation, as well as the influx of CD11b+, ED-1+ and myeloperoxidase+ leukocytes.
These experiments suggest that protective effects of caffeine in the neonatal lung are mediated, at least in part, by reduction of pulmonary inflammation.
Bronchopulmonary dysplasia (BPD), also known as chronic lung disease of prematurity, affects ∼50% of very low birth-weight infants with a gestational age <28 weeks [1] and is the most common chronic lung disease of infancy in industrialised countries [2]. BPD is associated with serial rehospitalisations during the first year of life, long-term pulmonary problems and persistent neurodevelopmental delays [2]. Oxidative stress plays an important role in the development of BPD [3]. Aeration of the lungs after birth already represents a sudden four- to five-fold increase of local oxygen tension in lung tissue, a switch for which preterm infants are ill prepared. In addition, some of these infants transiently require increased fractions of inspired oxygen (up to 100%) for pulmonary diseases, such as respiratory distress syndrome, pneumonia or primary pulmonary hypertension. Histologically, BPD features the hallmarks of various stages of inflammation, but attempts to treat early stages of BPD by systemic or inhaled corticosteroids yield only temporary relief and are associated with severe side-effects, such as cerebral palsy and gastrointestinal bleeding [4].
At present, few prophylactic interventions have been demonstrated by randomised controlled trials to significantly reduce rates of BPD: exogenous surfactant administration [5], initiating resuscitation in the delivery room with low rather than high concentrations of inspired oxygen [6]; vitamin A [7]; and caffeine [8]. Caffeine is a drug that has been introduced into the care of preterm newborn infants to treat apnoea of prematurity [9]. Caffeine given to preterm infants for the prevention or treatment of apnoea has been shown to reduce the rate of BPD, but it is unclear whether this protection is due to either reduced duration of mechanical ventilation or a direct pulmonary effect of caffeine. In a large randomised placebo-controlled multicentre trial involving 2006 preterm infants, the use of caffeine was found to reduce the duration of positive-pressure ventilation and to decrease the rate of BPD, as defined by the requirement of supplemental oxygen at 36 weeks gestation [8]. The protective effect was greatest when the administration of caffeine was started early, i.e. during the first 3 days of life [10]. The protective role of caffeine has been attributed to improved respiratory muscle strength and reduced rates of apnoea [11], which lead to reduced ventilator-induced lung injury and decreased requirement for mechanical ventilation. However, the direct pharmacological actions of caffeine on immature lungs have not yet been explored.
In animal models, early exposure to high oxygen concentrations results in progressive lung disease, which closely resembles BPD in premature infants [12, 13]. We investigated the effect of administering caffeine in an experimental, acute, lung-injury model by administering high concentrations of inspired oxygen to P6 rat pups at the transition from the saccular to alveolar phase of lung development [14], and analysing the expression of pulmonary chemoattractants, proinflammatory cytokines, extravasation of white blood cells into the lung tissue and histological signs of tissue destruction.
MATERIALS AND METHODS
Animals
6-day-old Wistar rats from timed-pregnant dams (Charité – Universitätsmedizin Berlin, Berlin, Germany) were divided into four biological groups: 1) normoxia (inspiratory oxygen fraction (FIO2) 21%, room air) and 0.9% NaCl solution i.p.; 2) normoxia and caffeine (Sigma, Steinheim, Germany) 10 mg·kg−1 body weight i.p.; 3) hyperoxia (FI,O2 80%, OxyCycler; BioSpherix, Lacona, NY, USA) and 0.9% NaCl solution i.p.; and 4) hyperoxia and caffeine 10 mg·kg−1 body weight i.p. Pups were treated once with saline or caffeine at the beginning of normoxia or hyperoxia lasting for 6, 24 or 48 h. For both conditions, pups were kept with their dam, and for the 48-h pups, nursing dams were switched every 24 h between the normoxic and hyperoxic chambers to provide equal nutrition to each litter. All procedures were approved by the state authorities (LAGeSo, Berlin, Germany) and followed institutional guidelines.
Tissue preparation
After 6, 24 and 48 h of oxygen exposure, pups were anaesthetised with an i.p. injection of ketamine (50 mg·kg−1) and xylazine (10 mg·kg−1). For molecular analysis, pups were transcardially perfused with normal saline and lungs were removed, snap-frozen in liquid nitrogen and stored at -80°C. For immunohistochemical analysis, pups were transcardially perfused with normal saline at pH 7.4, followed by 4% paraformaldehyde at pH 7.4. Lungs were removed, post-fixed for 3 days at 4°C, embedded in paraffin and processed for histological staining.
RNA extraction and realtime PCR
Total RNA was isolated by acidic phenol/chloroform extraction (peqGOLD RNApure; PEQLAB Biotechnologie, Erlangen, Germany) and 2 μg of RNA was reverse transcribed. The PCR products of cytokine-induced neutrophil chemoattractant (CINC)-1, macrophage inflammatory protein (MIP)-2, monocyte chemoattractant protein (MCP)-1, tumour necrosis factor (TNF)-α and interleukin (IL)-6 were quantified in real time, using dye-labelled fluorogenic reporter oligonucleotide probes/sequences (table 1). PCR and detection were performed in triplicate in 11-μL reaction mix which contained 5 μL of 2× KAPA PROBE FAST qPCR Mastermix (PEQLAB Biotechnologie), 2.5 μL of 1.25 μM oligonucleotide mix, 0.5 μL of 0.5 μM probe (BioTeZ, Berlin, Germany) and 3–15 ng cDNA template on a StepOnePlus™ Realtime PCR System (Applied Biosystems, Life Technologies, Carlsbad, CA, USA) with hypoxanthine-guanine phosphoribosyl-transferase (HPRT) used as an internal reference. PCR product levels were normalised to HPRT levels. The expression of target genes were analysed according to the 2-ΔΔCT method [15].
Immunohistochemistry
Tissue fixation
Paraffin-embedded lung tissues were cut in 10-μm sections and mounted onto SuperFrost Plus coated slides (Menzel, Braunschweig, Germany). Each section was deparaffinised in Roti-Histol (Carl Roth, Karlsruhe, Germany) twice for 10 min each, then rehydrated in ethanol (100, 90, 80 and 70%), distilled water and PBS for 3 min, each at room temperature.
Morphological examination
Hydrated sections were stained with hematoxylin and eosin. The pulmonary histological and morphometric changes were viewed by light microscopy using a Leica DM2000 microscope (Leica Microsystems, Wetzlar, Germany) equipped with a ×200 magnification. Digitally stored images were then analysed by Leica Application Suite (LAS) software.
Immunostaining
For increasing the permeability of the cell membrane, and thus demasking intracellular epitopes, sections were fixed in citrate buffer (pH 6.0) at 600 W for 12 min in a microwave oven. Sections were cooled and washed three times in PBS. The slices were incubated for 60 min in PBS containing 2% goat serum, 1% bovine serum albumin, 0.1% Triton X-100, 0.05% Tween 20, and 0.1% bovine gelatin, in order to block nonspecific proteins. Sections were washed once with PBS and subsequently incubated overnight at 4°C with monoclonal mouse anti-rat CD11b (OX42), predominantly expressed in monocytes and granulocytes, 1:200 (Abcam, Cambridge, UK) and monoclonal mouse anti-rat ED-1, a glycoprotein expressed by granulocytes, monocytes and tissue macrophages including alveolar macrophages, 1:100 (Abcam); or 30 min at room temperature with polyclonal rabbit anti-rat myeloperoxidase (MPO) which is expressed by neutrophils, 1:200 (Thermo Fisher Scientific, Schwerte, Germany) diluted in antibody diluent (Zymed Laboratories, San Francisco, CA, USA). For detection, a secondary fluorescein-conjugated goat anti-mouse IgG (Dianova, Hamburg, Germany) or fluorescein-conjugated goat anti-rabbit IgG (Dianova) were applied at a dilution of 1:200 in antibody diluent (Zymed Laboratories) and incubated at room temperature for 1 h. After being washed three times with PBS, slides were counterstained and mounted with aqueous 4,6-diamidino-2-phenylindole (DAPI) containing fluorescence-protecting mounting media (Vectashield HardSet Mounting Medium with DAPI; Vector Laboratories, Burlingame, CA, USA).
Sections were viewed under fluorescent light using a Leica DM2000 microscope equipped with a ×200 magnification and analysed using LAS software. Positive staining for CD11b-, ED-1-, and MPO-positive cells within the compartment of interest was counted in 10 separate fields per animal. We analysed cell numbers using ImageJ (National Institutes of Health, Bethesda, MD, USA) by creating a set threshold for particle size.
Statistical analyses
All data are expressed as mean±sem. Groups were compared using a one-way ANOVA, and significance was determined using Bonferroni's correction for multiple comparisons with an independent sample t-test. A two-sided p-value <0.05 was considered to be significant.
RESULTS
Hyperoxia leads to perturbation of alveolar development which is attenuated by caffeine
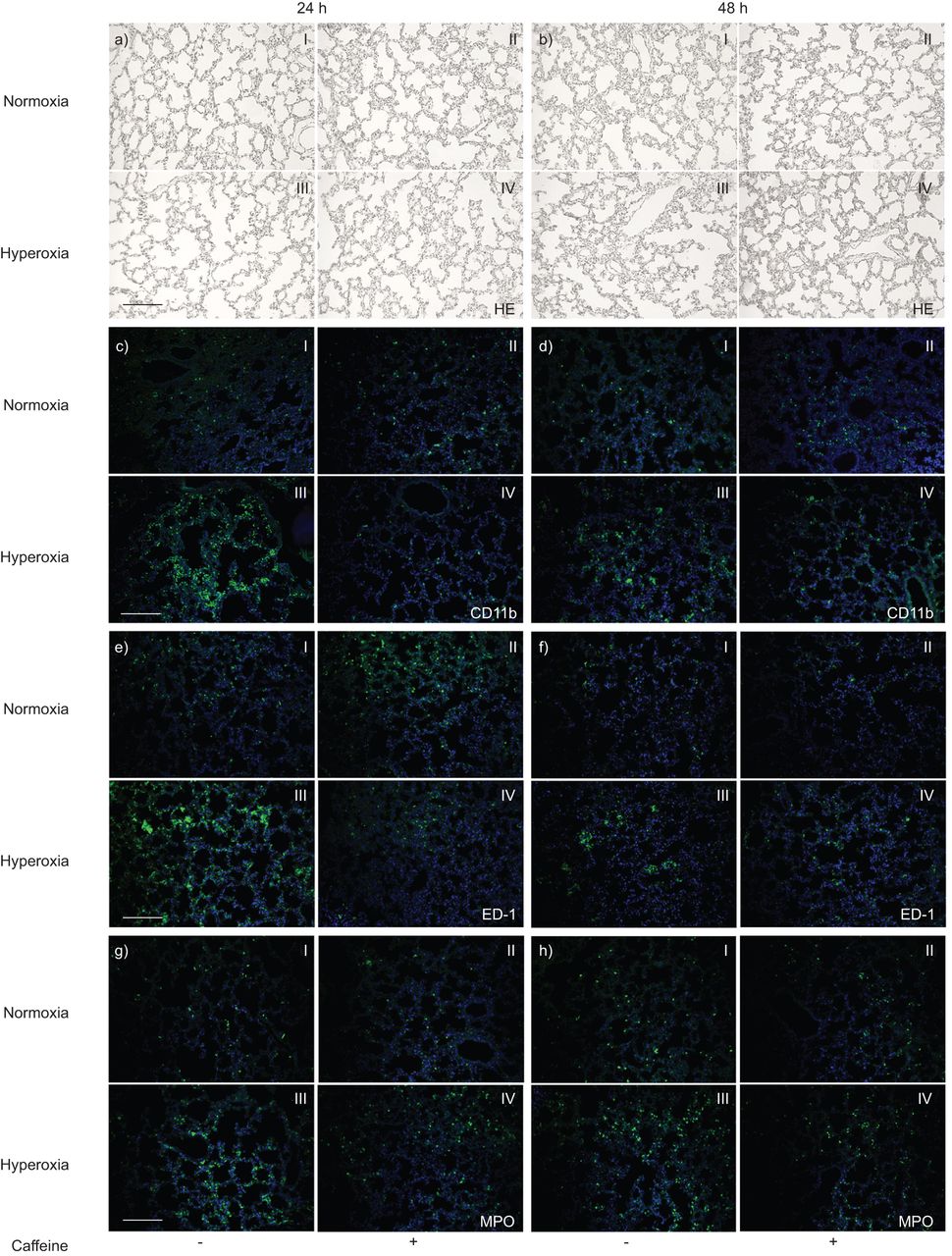
Exposure to high oxygen in 6-day-old rat pups for 24 h (to postnatal day (P)7, fig. 1aIII) or 48 h (to P8, fig. 1bIII) resulted in a heterogeneous distribution of enlarged air spaces, thinning of septa and oedema formation, which was reduced by caffeine (fig. 1a and bIV).

Paraffin lung sections stained with: haematoxylin and eosin (HE) a) at postnatal day (P)7 and b) P8; double immunofluorescence staining with anti-CD11b (green) and 4,6-diamidino-2-phenylindole (DAPI) nuclear staining (blue) c) at P7 and d) P8; anti-ED-1 (green) and DAPI nuclear staining (blue) [continued next page] e) at P7 and f) P8; and anti-myeloperoxidase (MPO) (green) and DAPI nuclear staining (blue) g) at P7 and h) P8. The pictures represent the lung sections of normoxia (I, II) or hyperoxia (III, IV), without (I, III) or with caffeine administration (II, IV). The lungs from the normoxic group without (aI and bI) and with (aII and bII) caffeine administration demonstrated normal alveolar development at P7 and P8. Hyperoxic exposure disrupted alveolar development at P7 (aIII) and P8 (bIII), which was attenuated by caffeine administration (aIV and bIV). Hyperoxia evokes pulmonary leukocyte infiltration, which is diminished by caffeine. Note the presence of large numbers of: CD11b+ cells at c) P7 and d) P8; ED-1+ cells at e) P7 and f) P8; and MPO+ cells at g) P7 and h) P8 in hyperoxia-exposed animals (III). Caffeine administration completely abolished (at 24 h: cIV, eIV and gIV) or markedly attenuated (at 48 h: dIV, fIV, hIV) this increase. All micrographs were taken at identical magnification. Scale bars=200 μm.
Hyperoxia evokes pulmonary leukocyte infiltration, which is diminished by caffeine
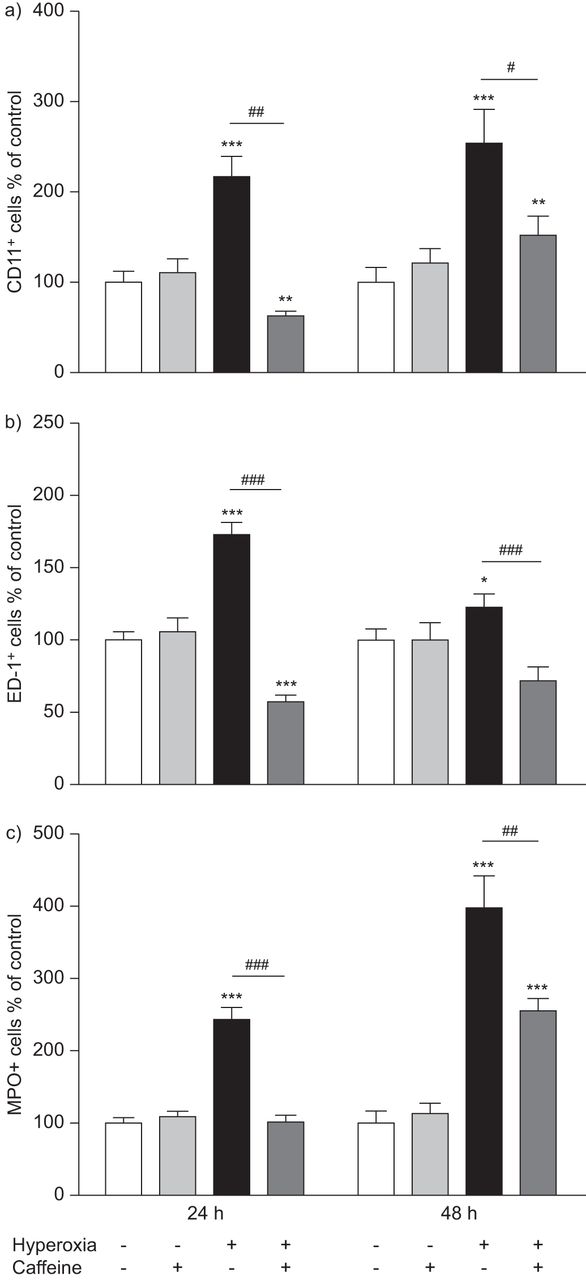
Hyperoxia led to a marked inflammatory reaction involving leukocytes (CD11b+), especially monocytes and macrophages (ED-1+) and neutrophil granulocytes (MPO+). In control animals in room air, a small number of resident CD11b+ (fig. 1cI and dI), ED-1+ (fig. 1eI and fI) and MPO+ (fig. 1gI and hI) cells were found in the septa and alveoli of the lungs. In pups exposed to hyperoxia, a markedly increased number of CD11b+ (fig. 1cIII and dIII), ED-1+ (fig. 1eIII and fIII) and MPO+ (fig. 1gIII and hIII) cells were identified. There was an increase in CD11b of 115±22.9% at 24 h (p<0.001) and 152±38.6% at 48 h (p<0.001), in ED-1 of 72±9% at 24 h (p<0.001) and 22±10.1% at 48 h (not significant), and in MPO of 142±17.2% at 24 h (p<0.001) and 296±45.2% at 48 h (p<0.001) of exposure to hyperoxia (fig. 2, black bars) compared to control pups kept in room air (fig. 2, white bars). Caffeine treatment significantly reduced the influx of CD11b+ (fig. 1cIV and dIV), ED-1+ (fig. 1eIV and fIV) and MPO+ (fig. 1gIV and hIV) immune cells in lungs of pups after hyperoxia compared to animals exposed to hyperoxia with saline injections. Treatment of pups with a single dose of caffeine (10 mg·kg−1 body weight) just before hyperoxic exposure significantly decreased the number of: CD11b+ cells, 3.4-fold (p<0.01) at 24 h and 1.7-fold (p<0.05) at 48 h (fig. 2a, dark grey bars); ED-1+ cells, three-fold (p<0.001) at 24 h and 1.7-fold (p<0.001) at 48 h (fig. 2b, dark grey bars); and of MPO+ cells, 2.4-fold (p<0.001) at 24 h and 1.6-fold (p<0.01) at 48 h (fig. 2c, dark grey bars). In rat pups kept in room air there was no significant effect of caffeine treatment on immune cell numbers in lung tissue (fig. 2, light grey bars).

Quantitative analysis of a) CD11b+, b) ED-1+, and c) MPO+ cell counts on paraffin sections showed a marked accumulation in lung tissue samples of rat pups that were kept for 24 and 48 h under hyperoxia, whereas caffeine treatment reduced the influx of different subtypes of leukocytes. Caffeine treatment under room air showed no significant regulation. Data are expressed relative to the normoxia-exposed control group as mean+sem of at least four rat pups per normoxia group and six rat pups per each experimental group. The 100% value is a) 1.985 and 0.537 CD11b+ cells·mm−2, b) 1.963 and 2.302 ED-1+ cells·mm−2 and c) 1.030 and 0.603 MPO+ cells·mm−2, for 24 h and 48 h groups, respectively. *: p<0.05, **: p<0.01 and ***: p<0.001 versus control; #: p<0.05, ##: p<0.01 and ###: p<0.001 versus hyperoxia (t-test).
Hyperoxia increases and caffeine decreases pulmonary mRNA expression of chemokines and proinflammatory cytokines
Chemokines are a prerequisite for leukocyte extravasation into inflamed tissue. Hence, we used realtime PCR to determine the differential expression of two pivotal CXC chemokines, CINC-1 and MIP-2, as well as the CC chemokine MCP-1, and two early proinflammatory cytokines, TNF-α and IL-6. Hyperoxia resulted in a time-dependent increase in mRNA expression of all three chemokines (fig. 3) and the two proinflammatory cytokines studied (fig. 4).

Caffeine prevents hyperoxia-induced increase in the lung mRNA expression of CXC- (cytokine-induced neutrophil chemoattractant (CINC)-1 and macrophage inflammatory protein (MIP)-2) and CC- (monocyte chemoattractant protein (MCP)-1) chemokines. Increased mRNA levels of a) CINC-1, b) MIP-2 and c) MCP-1, as detected by realtime PCR, were evident in total rat lung extracts at 6, 24 and 48 h of hyperoxia when compared to animals kept in room air. These levels were decreased through systemic caffeine pretreatment. Caffeine treatment under room air showed no relevant regulation. Results of realtime-PCR quantification are shown as mean+sem. Data are normalised to level of rat pups exposed to normoxia with saline (control 100%). CT: cycle threshold. *: p<0.05, **: p<0.01 and ***: p<0.001 versus control; #: p<0.05 and ##: p<0.01 versus hyperoxia; +: p<0.05, ++: p<0.01 and +++: p<0.001 versus intra-group (t-test, n=6 per group).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Caffeine prevents hyperoxia-induced increase in mRNA expression of the early cytokines tumour necrosis factor (TNF)-α and interleukin (IL)-6 in the lung. Increased mRNA levels of a) TNF-α and b) IL-6, as detected by realtime PCR, were evident in total rat lung extracts at 6, 24 and 48 h of hyperoxia when compared to animals kept in room air. These levels were decreased through systemic caffeine pretreatment at all time points from TNF-α (a), and at 24 h and 48 h for IL-6 (b). Caffeine treatment under room air showed no relevant regulation for TNF-α and a significant decrease for IL-6 at 6 and 48 h. Results of realtime PCR quantification are shown as mean+sem. Data are normalised to level of rat pups exposed to normoxia with saline (control 100%). CT: cycle threshold. *: p<0.05, **: p<0.01 and ***: p<0.001 versus control; #: p<0.05, ##: p<0.01 and ###: p<0.001 versus hyperoxia; +: p<0.05 and +++: p<0.001 versus intra-group (t-test, n=6 per group).
Hyperoxia caused a significant increase in CINC-1 expression at 6 h (247±66.5%; p<0.05), 24 h (222±44.1%; p<0.01) and 48 h (405±141.7%; p<0.05) (fig. 3a, black bars), a significant decrease in MIP-2 expression at 6 h (65±10.6%; p<0.01) and a significant increase at 48 h (167±63.7%; p<0.05) (fig. 3b, black bars), and a significant decrease at 6 h (64±6.7%; p<0.001) and significant increases at 24 h (91±32.7%; p<0.05) and 48 h (450±140.2%; p<0.05) of MCP-1 (fig. 3c, black bars). The mRNA expression of the proinflammatory cytokines under hyperoxic conditions showed a significant increase in TNF-α at 6 h (199±88.6%; p<0.05) and at 24 h (117±37.0%; p<0.05) (fig. 4a, black bars), a significant decrease at 6 h (88±1.7%; p<0.001) and significant increases in IL-6 at 24 h (122±38.3%; p<0.01) and 48 h (323±34.8%; p<0.01) (fig. 4b, black bars).
Caffeine treatment effectively abolished any hyperoxia-mediated increase in chemokine and proinflammatory cytokine mRNA expression. Treatment of pups with a single dose of caffeine (10 mg·kg−1 body weight) before hyperoxic exposure significantly decreased mRNA expression, when compared with normoxic controls of: CINC-1, 2.2-fold (p<0.05) at 6 h, 2.0-fold (p<0.05) at 24 h and 8.9-fold (p<0.01) at 48 h (fig. 3a, dark grey bars); MIP-2, 2.6-fold (p<0.05) at 48 h (fig. 3b, dark grey bars); MCP-1, 2.7-fold (p<0.01) at 24 h and 6.2-fold (p<0.05) at 48 h (fig. 3c, dark grey bars); TNF-α, 3.5-fold (p<0.01) at 6 h, 2.2-fold (p<0.05) at 24 h and 1.8-fold (p<0.05) at 48 h (fig. 4a, dark grey bars); and IL-6, 2.0-fold (p<0.05) at 24 h and 9.8-fold (p<0.001) at 48 h (fig. 4b, dark grey bars).
In normoxic animals, caffeine treatment caused a transient increase in CINC-1 expression at 6 h (253±18.8%; p<0.05), followed by a significant decrease at 48 h (53±5.3%; p<0.05) (fig. 3a, light grey bars). Caffeine also decreased MCP-1 at 6 h (52±9.9%; p<0.01) and 24 h (76±3.5%; p<0.05) (fig. 3c, light grey bars), as well as IL-6 at 6 h (23±9.0%; p<0.01) and 48 h (59±8.4%; p<0.05) (fig. 4b, light grey bars), compared with normoxic controls.
DISCUSSION
The results of these investigations demonstrate that 6–48 h of hyperoxia are sufficient to cause marked pulmonary inflammation in infant rats. The upregulation of chemokines and proinflammatory cytokines, the influx of neutrophils and macrophages, and the tissue destruction caused by short-term hyperoxia were found to be largely prevented by the administration of caffeine.
Exposing neonatal rodents to prolonged periods of hyperoxia (60–100% oxygen for 7–14 days) has been found to result in chronic lung disease closely resembling that observed in preterm human infants, and has, therefore, become an established animal model for BPD [12, 13, 16]. Our investigations, in contrast to most experiments reported previously, focus on the first 48 h of hyperoxia, and support the notion that avoiding even brief exposure to increased fractions of inspired oxygen is beneficial for the infant’s lungs. Our animal data correlate with clinical data from a prospective randomised trial assigning preterm infants ≤28 weeks gestational age to resuscitation with 90% or 30% oxygen during the first 5 min of life. In that study, rates of BPD later on were significantly lower in infants who were resuscitated with 30% oxygen. Moreover, plasma concentrations of the CXC chemokine IL-8 (the human homologue of CINC-1) were consistently elevated during the first 3 weeks of life in infants randomised to 90% oxygen, compared to those randomised to 30% [6].
In the animal experiments reported here, short-term hyperoxia upregulated the CXC chemokines CINC-1 and MIP-2, as well as the CC chemokine MCP-1. These chemokines have previously been demonstrated to be pivotal for the recruitment of leukocytes into pulmonary tissue [17, 18]. Furthermore, hyperoxia leads to a sequential induction of the proinflammatory nuclear factor-κB-regulated cytokines TNF-α and IL-6 which serve to amplify the activity of invading leukocytes [19]. Increased levels of chemokines (IL-8, MIP-1 and MCP-1) and cytokines (TNF-α and IL-6) have also been detected in airway secretions of preterm infants with BPD [20, 21].
Treatment with caffeine at the beginning of hyperoxia blocked the upregulation of chemokines and proinflammatory cytokines and the influx of myeloid leukocytes seen with high oxygen. Caffeine not only reduced mRNA expression and decreased inflammatory cell infiltration into pulmonary tissue, but also ameliorated histological damage of animals exposed to hyperoxia. We propose that the anti-inflammatory action of caffeine observed in these animal experiments underlies at least in part the reduced rate of BPD seen in preterm infants receiving caffeine [8]. However, while we found caffeine to effectively attenuate oxygen-induced chemokine expression, leukocyte influx and subsequent tissue damage, caffeine administered to animals kept in room air also promoted septal thinning. This seemingly paradoxical observation is mirrored by recent data from a murine model of oleic acid-induced acute lung injury, with attenuated neutrophil recruitment obtained by high-dose caffeine but enhanced lung tissue damage with low-dose caffeine [22].
The pharmacological effects of caffeine are well understood [23, 24], but its effects on other processes, such as inflammation, are still unclear [25–27]. Caffeine is a nonspecific adenosine receptor (AR) antagonist; blockade of one or more AR subtypes can produce the opposite effect [27, 28]. A study by Chavez Valdez et al. [29] showed that caffeine plasma levels outside this therapeutic range are associated with a proinflammatory cytokine profile of unknown clinical significance. Therefore, the variability of our data on cytokine expression could be explained by the nonlinear pharmacokinetics of caffeine and its metabolites [30, 31].
There are several limitations of this study pointing to areas of future investigations. First, we did not assess whether even shorter oxygen exposure times were sufficient to cause sustained inflammation, as suggested by the randomised controlled trial of Vento et al. [6]. Secondly, caffeine was given only once to the animals, while preterm infants receive caffeine for many weeks [10]. Thirdly, caffeine is thought to exert most of its actions via competitive nonspecific antagonism of ARs [32, 33], and investigating the involvement of these receptors has been beyond the focus of our experiments.
Data from adenosine A2a receptor knockout mice suggest that possible damaging effects of caffeine can be mediated via the adenosine A2a receptor, while the protective effects follow distinct pathways. Adenosine A2b receptors are important for modulating pulmonary inflammation in various animal models and in the pathogenesis of chronic obstructive pulmonary disease [34, 35].
Further investigations are needed to determine the contribution of A2a- and A2b-receptor and AR-independent mechanisms, e.g. by using specific pharmacological agonists and antagonists. However, based on the results of randomised clinical trials and our experimental data, caffeine and possibly other methylxanthines are promising candidates for the prevention of oxygen-induced lung injury in preterm infants.
Acknowledgments
We thank M. Blanco (Vanderbilt University, Nashville, TN, USA) for proofreading the manuscript.
Footnotes
Statement of Interest
None declared.
- Received January 20, 2012.
- Accepted July 19, 2012.
- ©ERS 2013
REFERENCES










