





Abstract
The role of mannose-binding lectin (MBL) deficiency (MBL2; XA/O and O/O genotypes) in host defences remains controversial. The surfactant proteins (SP)-A1, -A2 and -D, other collectins whose genes are located near MBL2, are part of the first-line lung defence against infection. We analysed the role of MBL on susceptibility to pneumococcal infection and the existence of linkage disequilibrium (LD) among the four genes.
We studied 348 patients with pneumococcal community-acquired pneumonia (P-CAP) and 2,110 controls. A meta-analysis of MBL2 genotypes in susceptibility to P-CAP and to invasive pneumococcal disease (IPD) was also performed. The extent of LD of MBL2 with SFTPA1, SFTPA2 and SFTPD was analysed.
MBL2 genotypes did not associate with either P-CAP or bacteraemic P-CAP in the case–control study. The MBL-deficient O/O genotype was significantly associated with higher risk of IPD in a meta-analysis, whereas the other MBL-deficient genotype (XA/O) showed a trend towards a protective role. We showed the existence of LD between MBL2 and SP genes.
The data do not support a role of MBL deficiency on susceptibility to P-CAP or to IPD. LD among MBL2 and SP genes must be considered in studies on the role of MBL in infectious diseases.
Community-acquired pneumonia (CAP) remains the leading cause of death from infection in developed countries [1]. Several microorganisms may be causative agents of CAP, but Streptococcus pneumoniae is the most common cause [1].
Mannose-binding lectin (MBL) is a serum collectin that promotes phagocytosis of microorganisms and initiates the lectin pathway of complement activation [2]. Deficient and low MBL serum levels are mainly due to the presence of three common point mutations in the exon 1 of the MBL2 gene (10q11.2-q21): alleles B, C and D are termed O alleles, and A is the wild-type allele. Heterozygous individuals for O alleles have reduced serum MBL levels and MBL-dependent lectin pathway activity, whereas these values are very low or absent in homozygous individuals for O alleles. The presence of the promoter allele X has an important downregulating effect, and O/O together with XA/O genotypes are considered MBL deficient genotypes, which are common in most populations [2, 3].
MBL deficiency has been considered a common primary immunodeficiency [4]. However, its role in host defence remains a matter of debate [5, 6]. An initial study suggested that O/O genotypes predispose to invasive pneumococcal disease (IPD) [7], but these results were not replicated in two other populations [8, 9]. The data from several studies argue against a role of MBL in host defences, particularly to pneumococcus [5, 10–13]. We have previously observed that MBL plays a redundant role in human defences against primary infection, at least in adults with CAP, but also that MBL insufficiency predisposes to higher severity and fatal outcome in CAP [14].
Surfactant proteins (SP)-A1, -A2 and -D, and other collectins, also promote phagocytosis of microorganisms and play a pivotal role in the regulation of the inflammatory response, as well as in clearance of apoptotic cells [15, 16]. SPs, but not MBL, take part in the first-line host defence in healthy lung. Genetic variability at genes coding for these SPs was associated with higher susceptibility and poor outcome of CAP [17]. The human SP-A locus consists of two similar genes, SFTPA1 and SFTPA2, localised within a cluster (10q21-24) that includes the SP-D gene (SFTPD) [15]. MBL2 was reported not to be in physical linkage with the genes of these SPs [18], but no studies of linkage disequilibrium (LD) of MBL2 with SFTPA1, SFTPA2 or SFTPD have been performed to date.
In the present study, we assessed the role of MBL2 genotypes in the susceptibility to and the severity of pneumococcal CAP (P-CAP). We also performed a meta-analysis aimed to analyse the role of MBL2 genotypes in susceptibility to P-CAP and to IPD. Lastly, we analysed the extent of LD of the most frequently studied single nucleotide polymorphisms (SNPs) of MBL2 with missense SNPs at SFTPA1, SFTPA2 and SFTPD.
METHODS
Patients and controls
In the present study, 1,398 white Spanish patients hospitalised with CAP (59.50±17.62 yrs, 34.8% females) from five Spanish hospitals (Hospital Universitario de Gran Canaria Doctor Negrín, Las Palmas de Gran Canaria; Hospital Clínico y Universitario de Valencia, Valencia; Hospital San Jorge de Huesca, Huesca; Hospital Universitario de la Princesa, Madrid; and Hospital Vall D`Hebron, Barcelona), were prospectively included. A total of 348 patients had P-CAP. The control group consisted of 2,110 unrelated healthy volunteers (blood and bone marrow donors as well as hospital staff) and patients without signs of relevant infectious diseases (47.27±17.40 yrs, 48.2% females) from the same origin as CAP patients. Foreigners and individuals with ancestors other than Spanish were previously excluded. Exclusion criteria and clinical definitions are shown in the Methods section of the online supplementary material. For susceptibility to P-CAP, a sex- and age-matched case–control study was performed; 340 patients and 1,736 controls were finally compared. Severity and outcome were evaluated in a prospective study of the 348 P-CAP patients.
In addition, we included a group of 84 patients with P-CAP (62.00±16.53 yrs, 32.1% females) from another Spanish population. These patients were included in a published study [19], but several patients were excluded on the basis of our inclusion/exclusion criteria. A group of 91 healthy controls from the same origin were also used (64.95±18.61 yrs, 72.5% females).
Informed consent was obtained from either the patients or their relatives. The protocol was approved by the local ethics committee of all hospitals. All steps were performed in complete accordance to the Helsinki declaration.
Genotyping
Genomic DNA was isolated as previously described [17]. The following MBL2 polymorphisms were analysed as described elsewhere [20]: codon 52 C/T (rs5030737, allele D), codon 54 A/G (rs1800450, allele B), codon 57 A/G (rs1800451, allele C) and codon -221 G/C (rs7096206, alleles X/Y). Haplotypes were simplified as YA, XA and O.
We have previously genotyped polymorphisms in SFTPA1 (aa19 T/C, rs1059047; aa50 G/C, rs1136450; aa219 C/T, 4253527), SFTPA2 (aa9 A/C, rs1059046; aa91 G/C, rs17886395; aa223 C/A, rs4253527) and SFTPD (aa11 T/C, rs721917) genes in part of both patient and control groups [17]. Haplotypes were named as 6An for SFTPA1 and 1An for SFTPA2 based on previous nomenclature [21]. For each individual, haplotypes were inferred using PHASE statistical software (version 2.1; available at www.stat.washington.edu/stephens/phase).
LD was measured by means of Arlequin (version 3.11; available at http://cmpg.unibe.ch/software/arlequin3) and Haploview (version 4.2; available at www.broadinstitute.org/haploview/haploview) softwares. Pairwise LD between MBL2 haplotypes and SP genes was characterised using Arlequin 3.11. The existence of LD was considered if D' >0.3.
Study selection for the meta-analysis
Eligible studies were identified by searching in PubMed using the search terms “mannose-binding lectin” or “mannose binding protein” and “pneumococcal” or “pneumonia”, and abstracts and references were reviewed for relevance. Full text of the relevant articles was reviewed to ensure that they met pre-set inclusion criteria. Data were extracted independently by two investigators, and the duplicate results were compared.
We identified 93 publications related to MBL and pneumonia or pneumococcus. After reviewing, 89 articles were excluded because they were reviews, met our exclusion criteria, or because they provided only serum data, data from patients with or without pneumococcus could not be separated, or they were irrelevant to the focus of this meta-analysis. Four studies remained after the selection process [7–9, 22]. Studies were separated into those focused on P-CAP or IPD. Three genotypes were studied in our meta-analysis: O/O, A/O+O/O and XA/O.
Statistical analysis
Quantitative variables are presented using arithmetic mean±sem. The comparison of MBL2 genotypes distribution based on the susceptibility, severity and outcome were performed using the Chi-squared test or Fisher’s exact test when needed, and odds ratios with 95% of confidence intervals were calculated. In addition, for the study of susceptibility, cases and controls were sex- and age-matched (considering intervals of 5 yrs), and the strata created were used for the conditional logistic regression analysis. The relationship between severity or outcome and genotypes was evaluated by binary logistic regression models, and hospital of origin and pneumonia severity index (PSI) were included as independent variables. The Hardy–Weinberg equilibrium for the genotypic frequencies was tested in the control groups by Chi-squared analysis. Assuming a frequency in our population of 0.15 for XA/O+O/O genotypes, our study (1,591 controls, 348 P-CAP patients and 96 bacteraemic P-CAP patients; incidence for bacteraemia of 0.28) had 80% power to detect an odds ratio of 1.53 and 2.03 for susceptibility to P-CAP and bacteraemic P-CAP, respectively. All tests were two-tailed. Statistical significance was taken as p<0.05. Statistical analysis was performed using SPSS 15.0 (SPSS Inc., Chicago, IL, USA). Meta-analyses were performed using DerSimonian and Laird random-effects models. For individual studies and pooled estimation, odds ratios and 95% confidence intervals are given. Heterogeneity was evaluated and it is showed in the forest plot. Analysis was performed using the Metafor package (available at www.jstatsoft.org/v36/i03/).
RESULTS
Patients admitted at five Spanish Hospitals were evaluated for the diagnosis of CAP. After excluding those patients without informed consent, ethnicity other than white Spanish and those that fulfilled the exclusion criteria, a total of 1,398 CAP patients were finally studied. S. pneumoniae was detected in 348 (24.89%) of these patients (57.43% of the patients with known causative microorganism). The main clinical characteristics of the P-CAP patients are shown in table 1.
Susceptibility to P-CAP related to MBL2 genotypes
No significant deviation from Hardy–Weinberg equilibrium of the studied MBL2 variants was found in our control population.
When MBL2 genotypes encompassing exon 1 wild-type (A) and mutated alleles (O), as well as promoter X/Y alleles, were analysed, no differences in A/O+ O/O or XA/O+O/O genotypes between sex- and age-matched P-CAP patients and controls were observed. However, the high-MBL genotype YA/YA was found to be over-represented in these patients, and genotype XA/YA was under-represented when compared with controls (tables 2 and 3).
Severity and outcome of P-CAP patients related to MBL2 genotypes
The relevance of MBL2 variants in the severity of P-CAP was analysed in our main cohort (tables 2 and 3). The high-MBL genotype YA/YA was under-represented in P-CAP patients with multi-organ dysfunctions (MODS). Likewise, the XA/XA genotype was under-represented in P-CAP patients with the most severe forms of sepsis (septic shock (SSh) and severe sepsis), as well as in those with acute respiratory failure (ARF) or moderate-to-high PSI. Conversely, the frequency of XA/O genotypes was found to be higher in P-CAP patients with SSh. In addition, when MBL-deficient genotypes (XA/O+O/O) were analysed, we found them to be associated with a need of intensive care unit admission, development of SSh and MODS; and with a moderate-to-high PSI at admission. Six of these associations remained significant in multivariate analysis including the variables hospital of origin and PSI (except for the analysis of the PSI, which only included hospital of origin): p=0.029 (OR 0.49, 95% CI 0.26–0.93) for YA/YA genotype in patients with MODS; p=0.005 (OR=0.12, 95% CI 0.03–0.53) and p=0.005 (OR=0.11, 95% CI 0.02–0.52) for XA/XA in patients with ARF and PSI IV–V, respectively; p=0.041 (OR=2.43, 95% CI 1.04–5.70) for XA/O in patients with SSh; and p=0.019 (OR=2.37, 95% CI 1.16–4.88) and p=0.038 (OR=1.63, 95% CI 1.03–2.58) in patients with PSI IV–V for XA/O+O/O and A/O+O/O genotypes, respectively. No significant differences were observed when acute respiratory distress syndrome, bacteraemia and fatal outcome were analysed (data not shown).
Data from another Spanish population
We also analysed data from another Spanish population included in a previously published study [19]. A total of 84 P-CAP patients (51.19% with bacteraemic P-CAP), and a group of 91 healthy controls were compared for MBL2 genotypes. This control population was in Hardy–Weinberg equilibrium. No relevant differences between patients and controls were observed. Data about severity were not available.
Association between MBL2 and pneumococcal infection: meta-analysis
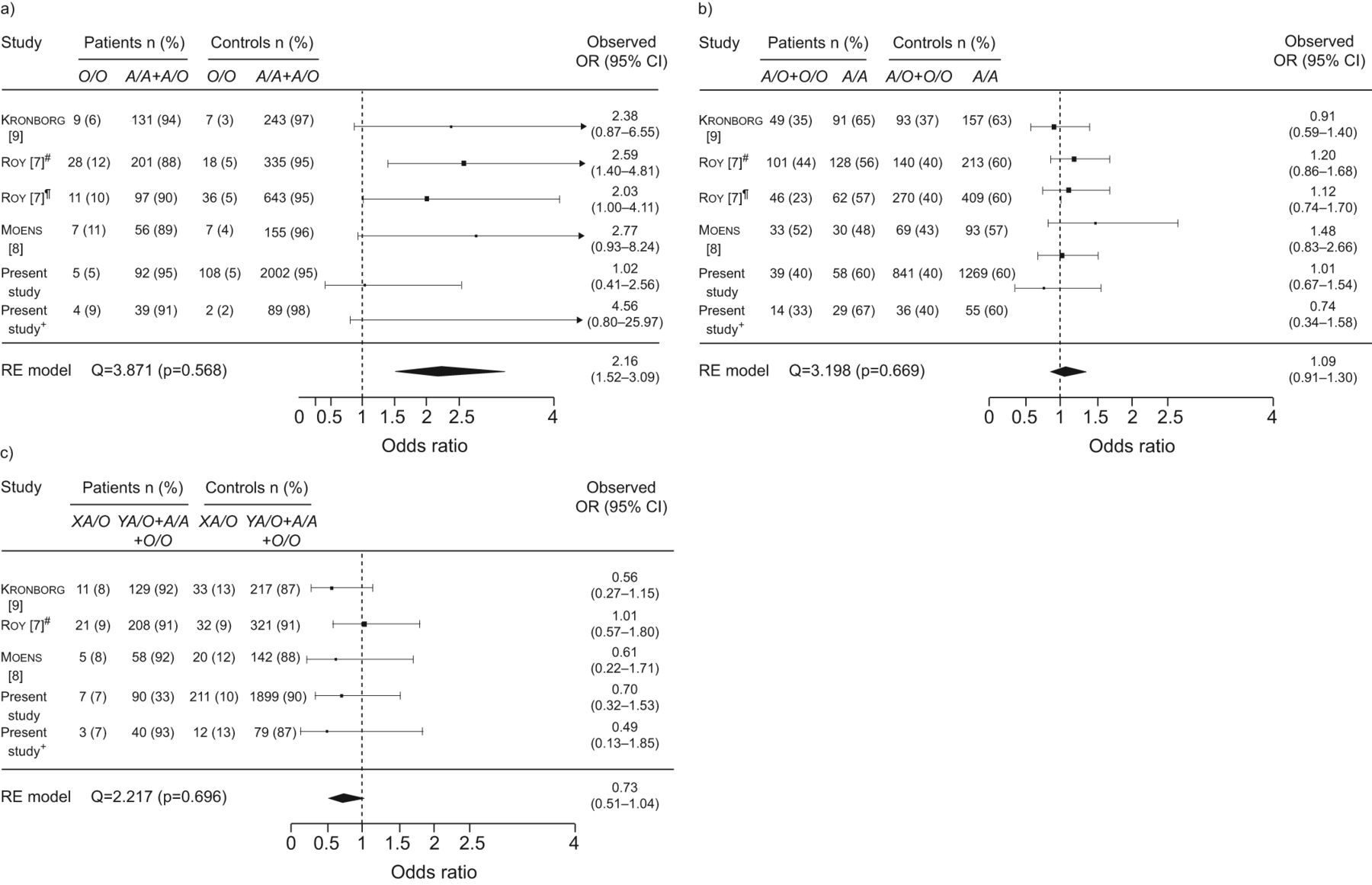
The characteristics of previous studies included in our meta-analyses are shown in table 4. The role of MBL2 genotypes on susceptibility to P-CAP was analysed in a meta-analysis including our two case–control studies and data from a previous study [22]. No differences of MBL2 genotypes between patients and controls were observed (fig. 1). There was no statistically significant heterogeneity among the included studies for any of the three meta-analysis.

Meta-analysis of association between MBL2 and pneumococcal pneumonia. RE: random effects. #: additional group of patients.
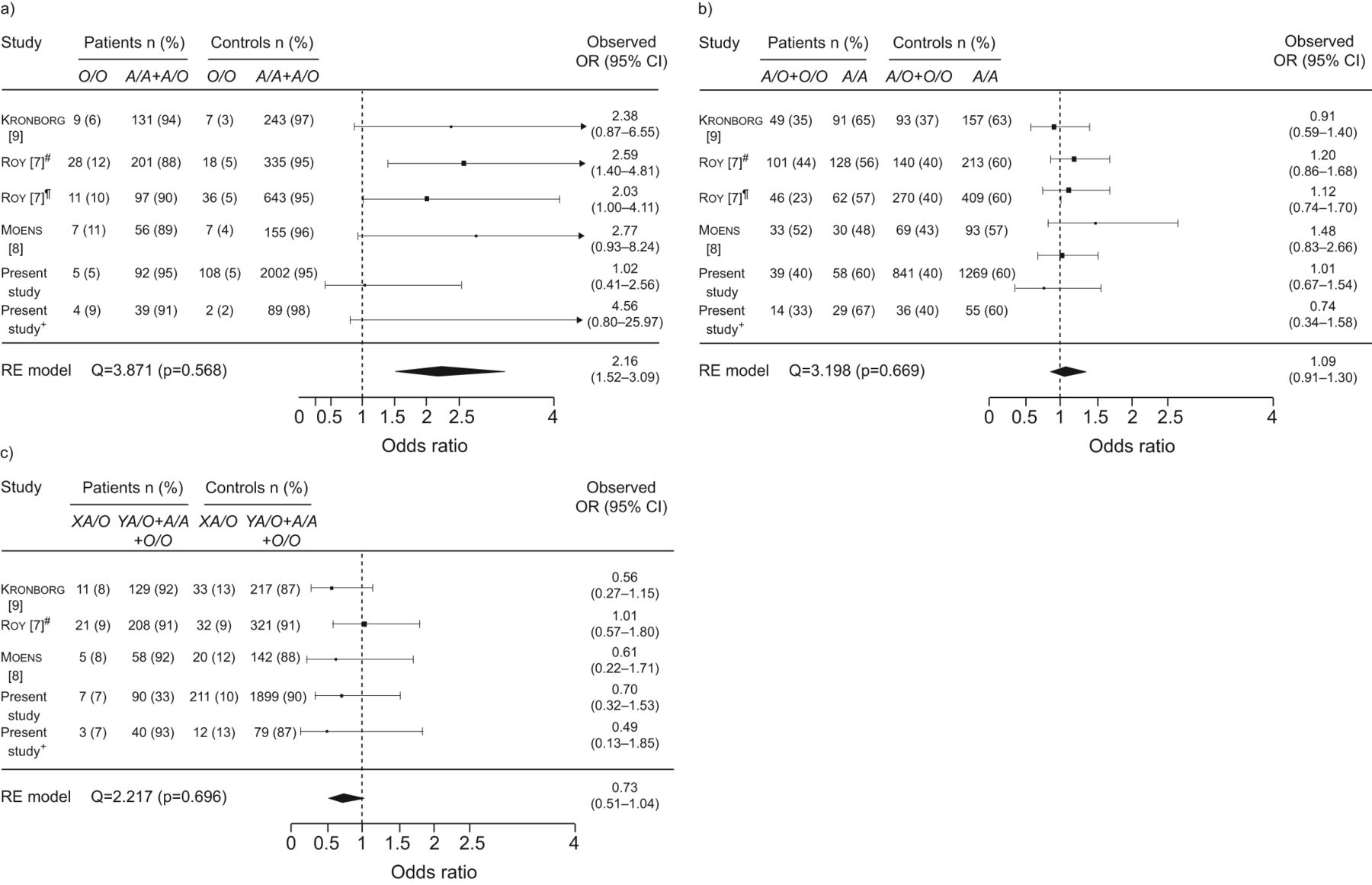
Three previous studies analysed the role of MBL2 genotypes in the susceptibility to IPD. Roy et al. [7] reported in two independent case–control studies that O/O homozygous patients have an increased risk of IPD, but these results were not replicated in other populations [8, 9]. Data from patients with bacteraemic P-CAP and healthy controls from our two case–control studies were independently included with those previous data in a meta-analysis. Figure 2a shows the O/O versus A/A+A/O forest plot. This analysis showed that the MBL-deficient genotype O/O was significantly associated with a risk of acquiring IPD (p<0.0001; pooled OR 2.16, 95% CI 1.52–3.09). No significant associations were found for the AO+O/O genotypes (fig. 2b). Nevertheless, the other MBL-deficient genotype, XA/O, showed a trend towards a protective role (pooled OR 0.73, 95% CI 0.51–1.04) (fig. 2c). There was no statistically significant heterogeneity among the included studies for any of the three meta-analysis. As expected, when XA/O+O/O genotypes were analysed, no significant differences were found (data not shown).
{kind=link}
{kind=link}
Meta-analysis of association between the mannose-binding lectin 2 gene (MBL2) and invasive pneumococcal disease. RE: random-effects. #: initial group of patients; ¶: confirmatory study in the same paper; +: additional group of patients.
LD of MBL2, SFTPA1, SFTPA2 and SFTPD genes
As we have previously shown in our population, there is LD among several SNPs at SFTPA1 and SFTPA2, whereas SFTPD aa11 was only observed in LD with SFTPA1 aa19 (fig. S1) [17]. As expected, pairwise LD (D') confirmed the existence of a very strong LD within MBL2 SNPs. Several SNPs of SFTPA1 and SFTPA2, but not the SFTPD aa11 SNP, were found to be in LD with MBL2 SNPs (fig. S1). The value of LD measured as r2 was very low for every pair of SNPs (data not shown), and none of the studied SNPs could be used as haplotype-tagging SNP to infer the observed haplotypes. In addition, when pairwise LD was measured among haplotypes instead of among SNPs; some haplotypes were found to be in LD with MBL2 variants (table 5).
Susceptibility to P-CAP related to haplotypes encompassing SFTPD, SFTPA1, SFTPA2 and MBL2
We also intended to analyse whether phased variants encompassing the four genes were involved in susceptibility to P-CAP. Due to the existence of LD, only 177 of the 2,048 expected haplotypes encompassing SFTPD, SFTPA1, SFTPA2 and MBL2 were observed, and only 18 had frequencies higher than 1% (data not shown). We previously reported a protective effect of the 6A2, 1A0, 6A2-1A0 and C-6A2-1A0 haplotypes on susceptibility to CAP [17]. When susceptibility to P-CAP was studied in the present study, the protective effect of these haplotypes was even higher when they co-segregate with the MBL2 XA variant (table 6). However, these results did not remain significant after a conservative Bonferroni correction for the number of observed haplotypes.
DISCUSSION
Previous meta-analysis based on genetic association studies concluded that the MBL2 O/O genotype predisposes to infection by S. pneumoniae. We herein provide new data showing that MBL2 genotypes are not involved in susceptibility to either P-CAP or to IPD.
Earlier studies from our group [14] and from Endeman et al. [22] failed to find any significant association between MBL2 genotypes and susceptibility to P-CAP, although both studies were underpowered to test it. When susceptibility to IPD was studied, only one out of three studies found a significant association with the O/O genotype [7–9]. However, a previous meta-analysis of these three studies yielded a significant association [23]. We have now studied the role of MBL2 genotypes on susceptibility to P-CAP and to invasive P-CAP in two different case–control studies, and no association was observed. We also performed a meta-analysis, including the three previous studies [7–9] and our results. Our meta-analysis also showed that O/O genotypes significantly associated with susceptibility to IPD. MBL deficiency is considered to result mainly from the presence of O/O or XA/O genotypes [2, 3]. However, when the genotype XA/O was analysed in our meta-analysis, a surprising trend towards a protective effect against IPD was observed in our meta-analysis. This data is intriguing, as the effect of O/O and XA/O genotypes on MBL levels and MBL-dependent LP activity was repeatedly found to be similar, as it has also been previously reported in our population [2, 14, 24–26]. More recently, a small study in children from Africa suggested that MBL deficiency would associate to IPD by means of low invasive serotypes [27].
MBL deficiency has been associated with infections by Gram-negative bacteria [6]. MBL binding to Pseudomonas aeruginosa has been documented, and susceptibility is conferred by both MBL-deficient genotypes (O/O and XA/O) [28]. However, the proposal of a putative role of MBL deficiency in susceptibility to pneumococcal infection is challenged by several functional and evolutionary studies. The lack of association between MBL deficiency and P-CAP would not be surprising, as no MBL binding and/or MBL-mediated opsonophagocytosis of S. pneumoniae was observed in vitro [10, 12]. The classical pathway of complement is the main complement pathway in host defences against pneumococcus in mice and humans, and loss of the lectin pathway does not seem to affect significantly innate immunity to pneumococcus in mice [11]. Infectious diseases have been the main selective force shaping the human genome during evolution. Pneumonia, particularly by S. pneumoniae, is the biggest killer worldwide of children aged <5 yrs [29]. If MBL plays a role in protective immunity against S. pneumoniae, then we would expect a selective removal of MBL2-deficient alleles. However, MBL2-deficient alleles are frequent in many populations worldwide. MBL deficiency was suggested to be protective against several inflammatory and infectious diseases, particularly against tuberculosis, which would positively select for low MBL genotypes. Nevertheless, the results from genetic association studies in the field of tuberculosis are controversial [6, 30], and recent studies have shown that the patterns of MBL2 variation worldwide are compatible with neutral evolution [5, 13].
The influence of a suspected LD among SFTPA1, SFTPA2 and/or SFTPD with MBL2 on the results of genetic association studies in infectious diseases has been recently proposed [6, 17]. We have previously found that several haplotypes of SFTPA1 and SFTPA2 are involved in susceptibility to CAP [17]. When we measured LD among MBL2, SFTPA1, SFTPA2 and SFTPD genes, some SNPs and haplotypes of both SFTPA genes were found to be in LD with MBL2 alleles. Interestingly, an additive effect of the MBL2 XA variant with variants at SFTPA1, SFTPA2 and SFTPD genes on protection against P-CAP was observed. Whether the observed association is due either to epistatic effects or to the existence of an extended protector haplotype encompassing the four genes, or both, is not known. In any event, our results suggest that the trend towards a protective effect of the MBL2 genotype XA/O on susceptibility to P-CAP might be due to LD with protector haplotypes on the genes of the studied SP. Likewise, the reported associations between the MBL2 O/O genotype with susceptibility to P-CAP might also be spurious, and due to LD with the SP genes. Wide genetic studies of this region are required to characterise the extent of LD among these genes and its relevance for genetic association studies.
In this and previous studies, XA/O+O/O genotypes and serum MBL-deficiency were associated with a poor prognosis of CAP [14] or pneumococcal disease [25]. Some objections to these results may be argued. How MBL can influence severity of P-CAP if it does not bind to pneumococcus? The rationale for the involvement of MBL deficiency in the severity of pneumonia could be its role in apoptosis [31], or its capacity to bind peptidoglycan and to inhibit peptidoglycan-induced pro-inflammatory cytokine production by macrophages [32]. However, only a weak statistical significance of MBL genotypes with severity or outcome was observed in our study, which could be even lower if the results were corrected for multiple comparisons. SFTPA1, SFTPA2 and SFTPD variants were found to be involved in severity of CAP [17] and, hence, LD of MBL2 with SP genes could underlie the association of MBL2 deficiency with higher severity and/or poor outcome in CAP and P-CAP. Vaccination against pneumococcus may obviously affect susceptibility to pneumococcal disease, particularly among high-risk individuals. Unfortunately, no data about the vaccination status of most patients against pneumococcus were available. However, pneumococcal vaccine is not included in the Spanish vaccination schedule and most adults have not been vaccinated against pneumococcus.
Overall, our data and several lines of evidence do not support a role of MBL deficiency on susceptibility to P-CAP or to IPD. By contrast, our results suggest that MBL deficiency may be associated with higher severity of P-CAP. However, studies aimed to analyse the role of genetic variability of the MBL gene in infectious diseases, particularly respiratory, should be aware of the existing LD among MBL2 and the genes of SP-A1, -A2 and -D. Identification of new pathways and molecules involved in susceptibility to and severity of respiratory infectious diseases could lead to new therapeutic approaches, and the therapeutic use of MBL and SPs has been advocated [15, 16, 33]. Large studies designed to analyse the genetic variability in the region of chromosome 10 containing these genes are desirable in order to unravel their role in susceptibility and severity of pneumococcal infection, particularly P-CAP.
Acknowledgments
The authors' affiliations are: M.I. García-Laorden, Dept of Immunology, Hospital Universitario de Gran Canaria Dr Negrín, Las Palmas de Gran Canaria, Spain; F. Rodríguez de Castro, Dept of Respiratory Diseases, Hospital Universitario de Gran Canaria Dr Negrín, and Dept of Medical and Surgical Sciences, School of Medicine, University of Las Palmas de Gran Canaria, Las Palmas de Gran Canaria; J. Solé-Violán, Intensive Care Unit, Hospital Universitario de Gran Canaria Dr Negrín; A. Payeras, Dept of Internal Medicine, Hospital Son Llatzer, Palma de Mallorca, Spain; M.L. Briones, Dept of Respiratory Diseases, Hospital Clínica y Universitario de Valencia, Valencia, Spain; L. Borderíal, Dept of Respiratory Diseases, Hospital San Jorge, Huesca, Spain; J. Aspa, Dept of Respiratory Diseases, Hospital Universitario de la Princesa, Madrid, Spain; J. Blanquer, Intensive Care Unit, Hospital Clínico y Universitario de Valencia; O. Rajas, Dept of Respiratory Diseases, Hospital Universitario de la Princesa; J.A. Marcos-Ramos, Intensive Care Unit, Hospital Dr José Molina Orosa, Lanzarote, Spain; E. Herrera-Ramos, Dept of Immunology, Hospital Universitario de Gran Canaria Dr Negrín; M.A. García-Bello, Research Unit, Hospital Universitario de Gran Canaria Dr Negrín; J. Noda, Dept of Immunology, Hospital Universitario de Gran Canaria Dr Negrín; J.M. Ferrer, Intensive Care Unit, Hospital Universitario de Gran Canaria Dr Negrín; J. Rello, Hospital Vall d'Hebron, Universitat Autonoma de Barcelona, CIBERES, Institut de Recerca Vall d'Hebron (VHIR). Barcelona, Spain; and C. Rodríguez-Gallego, Dept of Immunology, Hospital Universitario de Gran Canaria Dr Negrín and Dept of Medical and Surgical Sciences, School of Medicine, University of Las Palmas de Gran Canaria.
We are grateful to the patients and their families for their trust, as well as to the healthy volunteers. We also thank A.R. Domínguez-Acosta, N. González-Quevedo, Y. Florido (Dept of Immunology, Hospital Universitario de Gran Canaria Doctor Negrín), C. Ivañez (Dept of Respiratory Diseases, Hospital Universitario de Gran Canaria Doctor Negrín) and I. Martin-Loeches (Dept of Intensive Care Unit, Complejo Hospitalario Parc-Taulí) for their invaluable help, and P. Mangiaracina (Peter’s Language Services, Las Palmas de Gran Canaria, Spain) for his assistance with the final editing of the English manuscript.
Footnotes
This article has supplementary material available from www.erj.ersjournals.com
Support Statement
This work was supported by grants from “Fondo de Investigaciones Sanitarias”, Ministerio de Sanidad (FIS 02/1620, 04/1190, 06/1031 and 10/01718) with the funding of the European Regional Development Fund-European Social Fund (FEDER-FSE), RedRespira-ISCIII-RTIC-03/11, Sociedad Española de Neumología y Cirugía Torácica’ (SEPAR) FUNCIS, Gobierno de Canarias (04/09 and INREDCAN 5/06); M.I. García-Laorden was supported by FUNCIS (Proyecto Bioregion 2006) and E. Herrera-Ramos by a grant from Universidad de Las Palmas de Gran Canaria.
Statement of Interest
None declared.
- Received October 7, 2011.
- Accepted March 27, 2012.
- ©ERS 2013
REFERENCES










