





Abstract
Severity of respiratory syncytial virus (RSV) infection ranges widely. To what extent the local immune response is involved in RSV disease pathogenesis and which markers of this response are critical in determining disease severity is still a matter of debate.
The local immune response was studied in nasopharyngeal aspirates (NPAs) during RSV infection. 47 potential markers of disease severity were analysed in a screening cohort of RSV-infected infants with mild disease at home (n=8), hospitalised infants (n=10) and infants requiring mechanical ventilation (n=7). Results were confirmed in a cohort of infants hospitalised for RSV infection (n=200). Finally, genetic validation was studied in a cohort of infants hospitalised for RSV infection (n=465) and healthy controls (n=930).
The concentration of TIMP-1 (tissue inhibitor of metalloproteinase) was higher in the NPAs of hospitalised infants compared with the NPAs of infants at home (1,199 versus 568 ng·mL−1; p<0.0001). Similar results were found for matrix metalloproteinase (MMP)-3 (765 versus 370 pg·mL−1; p=0.004). MMP-3 was confirmed as a marker of disease severity in a larger cohort and MMP3 gene polymorphism rs522616 was associated with severe RSV infection (OR 0.82, p<0.05).
In conclusion, extracellular matrix proteinases play an important role in the pathogenesis of RSV bronchiolitis.
Respiratory syncytial virus (RSV) is the primary cause of hospitalisation for acute respiratory tract illness in young children [1]. The severity of RSV infection can range from mild upper respiratory tract infection (URTI) to severe lower respiratory tract infection (LRTI) and even death [2]. Most infants only develop URTI and recover in an outpatient setting; however, 1–3% require hospitalisation for observation and supportive care, and ∼10% of the hospitalised infants require mechanical ventilation for severe RSV LRTI [1, 3]. Infants requiring mechanical ventilation typically have low postconceptional age [4]. Well-defined high-risk groups for RSV LRTI include infants with pre-term birth, chronic lung disease of prematurity, congenital heart disease, cystic fibrosis, immunodeficiency disorders and Down's syndrome [5, 6]. Other risk factors for severe disease include age <3 months, crowded living conditions, indoor smoke pollution and male sex [5].
Apart from these known risk factors, little is known about the cause of the widely varying disease severity in previously healthy infants. As there is no effective therapy or broadly available vaccine for RSV infection, it is crucial for future management to characterise the precise mechanisms of this complex infection. A possible explanation of the varying disease severity could be a difference in the local immune response of the respiratory tract. Therefore, we hypothesised that variation in the local inflammatory response during primary RSV infection can be associated with severity of this respiratory tract infection. To study this hypothesis, we compared the local immune response of infants with varying severity of RSV infection: infants with mild RSV infection at home, infants admitted to hospital for RSV LRTI and infants mechanically ventilated for severe RSV LRTI. Markers that were associated with disease severity were subsequently measured in a larger cohort of infants hospitalised for severe RSV infection. Finally, genetic polymorphisms in the selected markers were tested for association with disease severity, because identification of genes that play an important role in disease susceptibility could enhance understanding disease development.
MATERIALS AND METHODS
Study populations
Screening cohort
Three groups of patients were distinguished by the need for hospitalisation and mechanical ventilation. The first group (n=8) consisted of infants with mild RSV infection who recovered at home. These infants participated in the Netherlands Amniotic Fluid (NAF) study [7], a cohort of infants born at term after an uncomplicated pregnancy. The second group (n=10) included infants hospitalised because of RSV LRTI. The third group (n=7) consisted of infants with severe RSV LRTI who were admitted to the paediatric intensive care unit (PICU) and required mechanical ventilation [8]. Selection criteria for the second and third groups were designed to enroll previously healthy children aged <13 months hospitalised with their first episode of RSV LRTI. Symptoms of LRTI were severe cough, wheezing, hoarseness, stridor, shortness of breath, cyanosis and apnoea. Infants with Down’s syndrome, a history of wheezing, or cardiac or pulmonary pathology were excluded. Undiluted nasopharyngeal aspirates (NPAs) of infants in all three groups were collected between October 2007 and March 2008 in the same region of the Netherlands, as described previously [7–9]. All NPAs were collected and processed using uniform methodologies and study personnel were all trained in the same way.
Confirmation cohort
Protein markers that were associated with RSV disease severity in the screening cohort were additionally studied in the NPAs of infants included in the RSV-NPA study to confirm that differences were not due to spurious findings in the small screening cohort. This study consisted of 235 infants hospitalised for RSV infection. 23 of these hospitalised infants required mechanical ventilation, while no infants with mild RSV disease recovering at home were present in this cohort. Inclusion criteria for the RSV-NPA study were previously healthy children aged <13 months hospitalised with a first episode of RSV LRTI. RSV infection was confirmed by positive immunofluorescence in epithelial cells from NPAs. Infants with Down’s syndrome, a history of wheezing, or cardiac or pulmonary pathology were excluded. Identical techniques as in the screening cohort were used to obtain undiluted NPAs of the infants collected between October 2007 and March 2009 in 15 large urban hospitals in the Netherlands.
Cohort for genetic validation
For genetic association analysis, the data of two cohorts of infants with severe RSV infection were combined. One cohort consisted of 480 previously healthy infants hospitalised for RSV LRTI (RSV-GENE study) [10–12] and the other cohort of 235 infants in the RSV-NPA study described above. Only the Dutch infants of these two cohorts (n=465) were used for genetic analysis. 81 of these infants required additional mechanical ventilation at the PICU. Genotype distribution was compared with that of 930 controls randomly taken from the Regenboog study, a large Dutch population health examination survey [13], as described previously [10]. All three cohort studies were approved by the ethics review committees of all participating hospitals. Written informed consent was obtained from parents of all infants before enrolment.
Detection
After collection, the NPAs of the infants were stored at -80°C. Before measurement of markers, samples of the screening cohort were weighed, diluted, sonicated and centrifuged as described previously [9]. Frozen samples were transported to Rules-Based Medicine (RBM, Austin, TX, USA), where 150 μL were used to measure marker concentrations in a Luminex 100 instrument (Luminex, Austin, TX, USA). 90 markers were measured in the human Multi-Analyte Profile 1.6 (Myriad RBM, Austin, TX, USA) for selection of protein markers associated with disease severity. However, this assay was developed for measurement of serum markers. As, in this study, the assay was used for detection of markers in NPA, the first three NPAs were measured at different dilutions to select those markers that could be detected in an accurate manner in the NPAs. This analysis also showed that the levels of 10 markers were too low in NPA to be considered for statistical analysis (calcitonin, erythropoietin, granulocyte macrophage-colony stimulating factor, immunoglobulin (Ig)E, and interleukin (IL)-2, IL-5, IL-12p40, IL-12p70, IL-13 and IL-15). In total, 47 different markers could be measured accurately and were considered clinically relevant for analysis in the NPAs of infants with RSV infection.
From 200 infants participating in the RSV-NPA study, an adequate volume of NPA was available to confirm the results of the screening. The NPAs were weighed and diluted in PBS (4°C) (Tritium Microbiology, Eindhoven, the Netherlands), sonicated on ice for 15 s, output control at 2, duty cycle at 50% with a Branson sonifier type 250 with microtip (VWR, Amsterdam, the Netherlands), and were centrifuged at 10,000×g for 5 min at 4°C. Concentrations of total matrix metalloproteinase (MMP)-3 and tissue inhibitor of metalloproteinase-1 (TIMP) were measured using commercial ELISA kits according to the manufacturer's instructions (R&D Systems, Abingdon, UK). ELISAs were performed on the same day for all samples and groups were randomly distributed across the plates.
Genotyping
DNA was isolated as described before [14]. Single-nucleotide polymorphisms (SNPs) rs522616 and rs679620 in MMP3, and rs4898 in TIMP1 were genotyped by SNP Genotyping Services at KBioscience (Hoddesdon, UK) with the KASPar technology for all 465 Dutch infants of the RSV-NPA study and the RSV-GENE study, and 930 population controls from the RSV-GENE study. For each SNP, 1.5 μL genomic DNA was used at 7 ng·μL−1. If less genomic DNA was available, whole-genomic amplification was performed at KBioscience using primer extension pre-amplification and a thermostable DNA polymerase. Genotyping failed for 1.1% of the samples. Genotyping data were viewed graphically as a scatter plot with SNPviewer2 (Kbioscience, Hoddesdon, UK). All SNPs were in Hardy–Weinberg equilibrium and were analysed for association with severe RSV disease.
Statistical analyses
Marker levels among the three groups were compared using logarithmic transformation and one-way ANOVA testing. A false discovery rate of 5% with correction according to Benjamini and Hochberg [15] was used to correct for multiple comparisons. Only if a significant difference was observed among the groups, a Fisher post hoc test with Bonferroni adjustment was used to determine the exact differences between the groups. Marker levels among two groups were analysed with the Mann–Whitney U-test. Genetic analyses were performed with a Chi-squared distribution of a 2×2 table. The Mann–Whitney U-test was used to compare length of hospital stay between two groups or with the Kruskal–Wallis test in case of three groups. All analyses were performed using SPSS Statistics 17.0 (SPSS Inc., Chicago, IL, USA) and R software (R Development Core Team 2011; R Foundation for Statistical Computing, Vienna, Austria). All hypothesis testing was two-sided, with a 5% threshold for statistical significance. The TIMP1 gene is located on the X-chromosome; therefore, males and females were analysed separately.
RESULTS
Selection of TIMP-1 and MMP-3 as markers of RSV disease severity
The screening cohort consisted of a group of infants with primary RSV infection and mild disease (nonhospitalised), a group of hospitalised RSV-infected infants and a group of mechanically ventilated infants with severe RSV disease. In this cohort, most included infants were male (84%) and were born full-term (84%). The infants with mild disease were older at the time of infection than the infants who were admitted to the hospital for severe RSV infection (table 1). Gestational age of the infants did not differ among the three groups.
The levels of 47 markers that could be accurately measured in NPAs (for details see Materials and methods) were compared among the three groups of the screening cohort. Five markers were present at higher levels in the hospitalised infants only and were not elevated in the mechanically ventilated infants. These markers (endothelin-1, chemokine (C-C motif) ligand (CCL)11, S100 calcium-binding protein A12 (S100A12), IL-10 and IgM) did not, therefore, correlate with disease severity (table 2). Three markers were associated with disease severity, i.e. they were present in higher levels in hospitalised and mechanically ventilated infants compared with infants with mild RSV infection at home. The level of TIMP-1 was higher in the NPAs of both hospitalised and mechanically ventilated infants compared with the NPAs of infants at home (table 2 and online supplementary fig. S1). TIMP-1 is a natural inhibitor of the MMPs, which are involved in degradation of the extracellular matrix. The extracellular matrix provides support and anchorage for cells, segregates tissues from one another and regulates intercellular communication. In addition to the different level of TIMP-1 in the NPAs, the concentration of MMP-3 was higher in the NPAs of ventilated infants compared with the NPAs of infants at home (table 2 and online supplementary fig. S1). Furthermore, the level of growth hormone was higher in the NPAs of hospitalised or mechanically ventilated infants for severe RSV LRTI compared to the NPAs of infants who stayed at home (table 2 and online supplementary fig. S1).
Confirmation of MMP-3 as marker of RSV disease severity
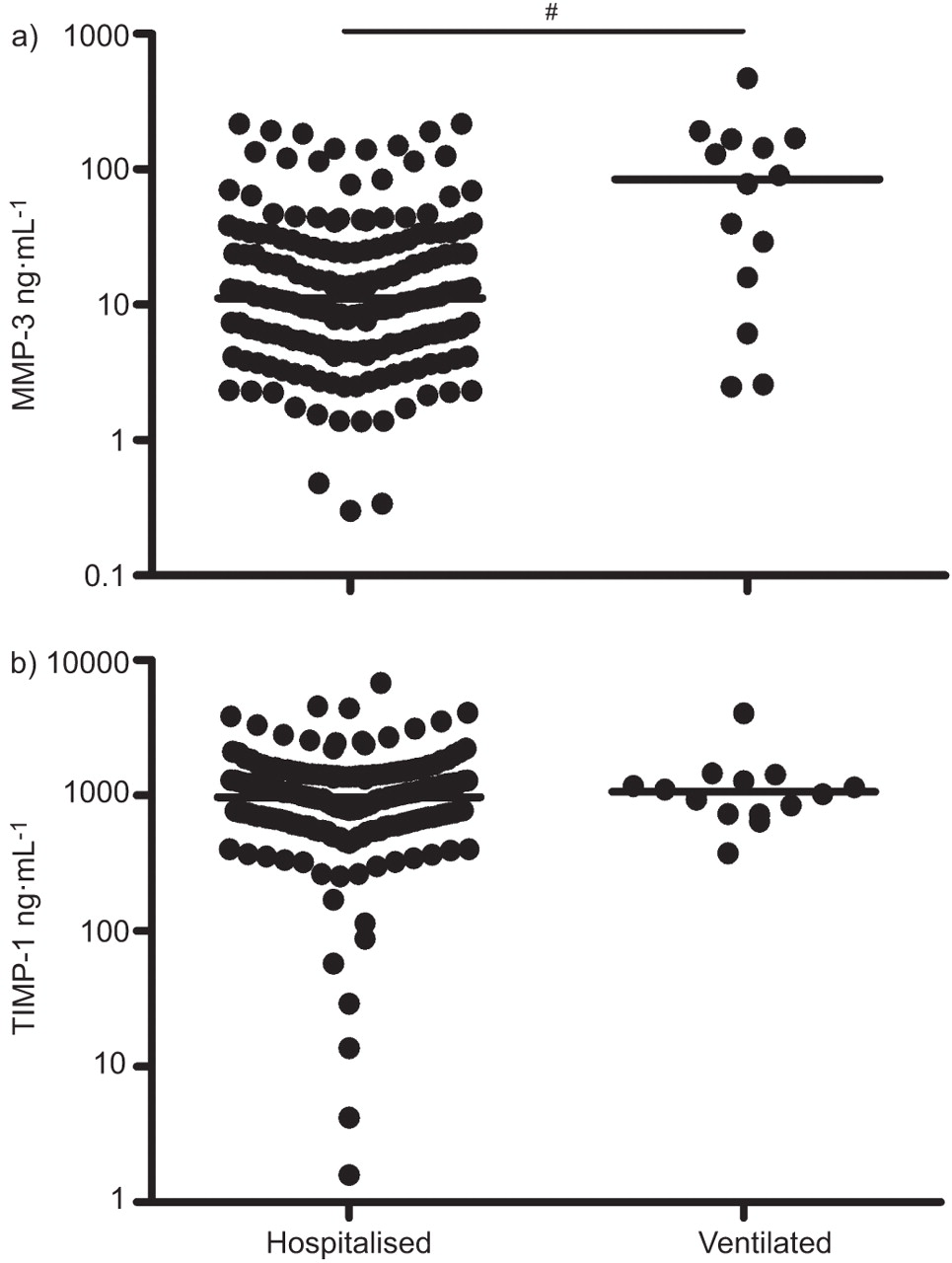
To confirm the association of TIMP-1 and MMP-3 with severity of RSV disease, the levels of TIMP-1 and MMP-3 were analysed in the RSV-NPA study. NPAs of a large cohort composed of infants admitted to hospital for RSV LRTI and infants mechanically ventilated for severe RSV LRTI were analysed. General characteristics of these infants are shown in table 1. More males were included than females. Mechanically ventilated infants were younger at the time of infection than the hospitalised infants without ventilation. figure 1 shows that the level of MMP-3 was significantly higher in the NPAs of mechanically ventilated infants compared with the NPAs of hospitalised infants. The level of TIMP-1 was comparable between the two groups. In order to assess potential confounding by age and sex, these two characteristics were analysed in relation to TIMP-1 and MMP-3 concentrations. There was no statistical association between sex or age and levels of TIMP-1 and MMP-3.

Confirmation cohort: matrix metalloproteinase (MMP)-3 is associated with respiratory syncytial virus (RSV) disease severity in the nasopharyngeal aspirates (NPAs) of RSV-infected infants. Levels of a) MMP-3 and b) tissue inhibitor of metalloproteinase (TIMP) 1 were analysed in NPAs of infants hospitalised for RSV lower respiratory tract infection (LRTI) (n=186) or mechanically ventilated infants at the paediatric intensive care unit with RSV LRTI (n=14). ——: median for analysed groups. #: p=0.002 by Mann–Whitney U-test.
Genetic validation of TIMP1 and MMP3 in the pathogenesis of RSV disease
Markers that are positively associated with disease severity could either be involved in determining the disease severity, or they could merely be a reflection of the extent of pathology. Genetic association of the same genes would favour a role in determining disease severity. Therefore, a SNP in TIMP1 and two SNPs in MMP3 were genotyped in 465 hospitalised infants with RSV LRTI and 930 population controls. SNP rs522616 in the MMP3 gene was associated with RSV disease severity at the allele level (table 3). The major allele of this SNP in the promoter region of MMP3 had a lower frequency in infants with RSV LRTI compared with controls (OR 0.821, 95% CI 0.677–0.996). SNPs rs679620 in MMP3 and rs4898 in TIMP1 were not associated when infants with RSV LRTI were compared with population controls. However, when a subgroup of males who required mechanical ventilation for RSV infection was compared with male controls, an association was found for TIMP1. The T-allele of rs4898 had a higher frequency in mechanically ventilated boys with RSV LRTI (OR 2.168, 95% CI 1.161–4.049) (table 4). In females, no association was found between the TIMP1 SNP and severe RSV LRTI. No association was found between levels of TIMP-1 or MMP-3 and the corresponding SNPs.
Additionally, a secondary outcome of disease severity was analysed. Length of stay in the hospital was determined in 219 male and 166 female infants hospitalised for RSV LRTI. SNP rs4898 in TIMP1 was associated with length of stay in the hospital of RSV-infected males but not females (fig. 2). A subgroup analysis excluding mechanically ventilated infants showed that the association remained significant. SNPs in MMP3 were not associated with duration of hospitalisation (data not shown).

{kind=link}
{kind=link}
Genetic validation cohort: tissue inhibitor of metalloproteinase (TIMP)-1 single-nucleotide polymorphism (SNP) rs4898 in respiratory syncytial virus (RSV)-infected males is associated with length of stay in the hospital. SNP rs4898 in TIMP1 was associated with length of stay in the hospital of RSV-infected males but not in females. Genotypes and length of stay were determined in a) 219 male infants (p=0.005 by the Mann–Whitney U-test) and b) 166 female infants hospitalised for RSV lower respiratory tract infection.
DISCUSSION
This study demonstrates that both a MMP and a metalloproteinase inhibitor affect disease severity in infants with RSV infection. Higher levels of TIMP-1 and MMP-3 were found in the NPAs of hospitalised infants with RSV LRTI compared with the NPAs of infants at home with RSV infection. Interestingly, MMP-3 and TIMP-1 are closely related in function and are both involved in the dynamics of the extracellular matrix, indicating that this process is important in determining disease severity. Confirmation of this association with severe RSV disease found in the screening cohort was seen for MMP-3 in a large cohort of hospitalised and mechanically ventilated infants with RSV LRTI. The level of TIMP-1 was comparable between the two groups. However, this was not surprising as no infants with mild disease at home were included in this confirmation cohort and, in our screening cohort, TIMP-1 was elevated to a similar extent in hospitalised and mechanically ventilated infants. Furthermore, SNPs in the corresponding genes of these markers were associated with RSV disease severity in a large cohort, favouring the hypothesis that these genes are involved in determining disease severity and that higher protein levels are not merely a reflection of the extent of pathology.
The panel of markers tested in this study also included three other proteins involved in extracellular matrix formation, i.e. MMP-2, MMP-9 and S100A12. MMP-2 showed a trend in association; however, no difference was seen after correction for multiple testing (table 2). S100A12 is a multifunctional, pro-inflammatory extracellular factor that inhibits MMP-2, MMP-3 and MMP-9 by chelating Zn2+ ions [16, 17]. A higher level of S100A12 was observed in the NPAs of hospitalised infants but not in mechanically ventilated children. These results highlight the importance of proteins involved in the dynamics of extracellular matrix with RSV disease severity. In contrast, the MMP-9 level in NPAs was comparable among the three groups with different RSV disease severity.
MMPs are Zn2+-dependent proteases classified in terms of substrate specificity. 25 known MMPs are capable of degrading all kinds of extracellular matrix proteins [18, 19]. They are also implicated in the functional regulation of a host of nonextracellular matrix molecules, such as cytokines and chemokines (reviewed in [19–22]). TIMPs regulate MMP activity during tissue remodelling, where each TIMP is the inhibitor of a specific set of MMPs [23, 24]. Therefore, the ratio of for instance TIMP-1 over MMP-9 is often used as a measure for MMP-9 activity. In this study, a ratio of MMP-3 over TIMP-1 was not added as TIMP-1 levels were similar in both groups of the confirmation cohort. Because TIMP-1 also inhibits MMPs other than MMP-3, this ratio also does not aid in drawing conclusions on the activity of MMP-3 in the nasal aspirate. MMP activity has been implicated in the severity of a range of diseases including infectious diseases. For example, TIMP-1 levels are higher in nonsurvivors compared with survivors of severe sepsis [25]. Furthermore, TIMP-1-deficient mice show enhanced resistance to pulmonary infection with Pseudomonas aeruginosa [26], indicating that inhibition of MMP activity is related to a reduced capacity to clear infection. Conversely, in vitro studies have implicated TIMP-1 in inhibiting viral replication and syncytia formation [27]. These data seem to contradict our findings; however, the balance between tissue injury and viral clearance in such a model is not comparable to the in vivo situation. While MMP activity may be beneficial for clearance of infection in vivo, it is also implicated in enhancing tissue injury. Mice lacking MMP-3 demonstrate less tissue injury than their wild-type counterparts following the formation of IgG-containing immune complexes in the alveolar wall [28]. In accordance with this, Davies et al. [29] showed that the level of MMP-9/TIMP-1 complex is higher in infants developing chronic lung disease of prematurity than in infants with resolved respiratory distress syndrome, indicating that this complex is important during ongoing respiratory morbidity. Elliott et al. [18] reported that hypoxic bronchiolitis caused by RSV showed some association with greater amounts of TIMP-1 protein compared with mild RSV disease, although their data did not reach statistical significance.
Taken together, these data indicate that MMP activity is essential to clear infection while promoting tissue injury. Accordingly, higher levels of TIMP-1 in infants with severe RSV infection can be an attempt to limit the injury of severe disease, which may come at the cost of reduced clearance of the infection. Therefore, tight control of MMP activity through modulation of TIMP or MMP levels may be of therapeutic benefit. However, a complete understanding of the biology of these complex enzymes is needed before we can reasonably consider them as therapeutic targets [19].
Our study confirmed the results of Tasker et al. [30] who showed, in serum, that during the acute phase of RSV LRTI, infants requiring mechanical ventilation have higher growth hormone levels in plasma compared with hospitalised, nonventilated infants. This higher level of growth hormone in plasma can explain the higher level found in NPAs in this study.
Cytokine and chemokine levels in the lungs of RSV-infected infants have been studied extensively and chemokine levels have been implicated in disease severity [31]. Our results show that high levels of chemokines (IL-8, CCL2, CCL3, CCL5 and CCL11; table 2) are present in the NPAs of infants with severe RSV disease, consistent with levels previously described in bronchoalveolar lavage samples by McNamara et al. [31]. However, we also observed similar high levels of these chemokines in infants with mild RSV disease. Therefore, the role of chemokines in determining the severity of RSV disease might be limited.
In conclusion, the combination of these immunological and genetic data indicate that proteins involved in extracellular matrix degradation may play an important role in the pathogenesis of RSV bronchiolitis, providing a novel potential target for intervention.
Acknowledgments
We thank C. Tacke, B. Waalewijn, E. Schouten (all at Dept of Pediatrics, Wilhelmina Children’s Hospital, University Medical Centre Utrecht, Utrecht, the Netherlands), and all paediatricians, nurses and laboratory technicians for their help with sample collection in Flevo Hospital, Almere; Meander Medical Center, Amersfoort; Gelre Hospital, Apeldoorn; Alysis Rijnstate Hospital, Arnhem; Gelderse Vallei Hospital, Ede; Rivas Beatrix Hospital, Gorinchem; Jeroen Bosch Hospital, 's-Hertogenbosch; Tergooi Hospital, Blaricum; St Antonius Hospital, Nieuwegein; Twee Steden Hospital, Tilburg; Diakonessen Hospital, Utrecht; Mesos Medical Center, Utrecht; University Medical Center, Utrecht; Maxima Medical Center, Veldhoven; Zuwe Hofpoort Hospital, Woerden, all in the Netherlands. We are grateful to M. Belderbos (Dept of Pediatrics, Wilhelmina Children’s Hospital, University Medical Centre Utrecht) for her laboratory assistance.
Footnotes
This article has supplementary material available from www.erj.ersjournals.com
Statement of Interest
None declared.
- Received January 22, 2011.
- Accepted November 13, 2011.
- ©ERS 2012
REFERENCES










