





Abstract
Aspergillus fumigatus is a fungal pathogen of major clinical importance. However, little is known about the role of human bronchial epithelial cells (HBECs) during A. fumigatus conidia induced inflammation.
Here, we show that differentiated respiratory epithelial cells recognise inactivated resting conidia but not swollen conidia or hyphae, resulting in the induction of the interferon (IFN)-β signalling pathway and the expression of IFN-β-inducible genes, such as IFN-γ-inducible protein (IP)-10. This induction was internalisation dependent.
We identified double-stranded conidial RNA recognised by Toll-like receptor-3 as a factor responsible for the expression of IFN-β and IP-10. Inhibition of receptor-interacting protein-1/TANK-binding kinase-1, known to mediate IFN-β signalling, was sufficient to inhibit the induction of IFN-β and IP-10 expression by conidia. Even though conidia induced the activation of nuclear factor (NF)-κB in HBECs, IP-10 expression was only partially dependent on NF-κB signalling.
These results provide evidence that respiratory cells are activated by the double-stranded RNA of resting conidia and initiate a first immune response to inhaled conidia in an IFN-β-dependent manner.
The fungal pathogen Aspergillus fumigatus is of major clinical importance. Inhalation of fungal spores may cause lung diseases ranging from local inflammation of the airways to severe and life-threatening infections of the lung, such as allergic bronchopulmonary aspergillosis or invasive aspergillosis [1, 2]. Even though every human being inhales ∼100 A. fumigatus conidia every day, severe Aspergillus infections are relatively rare in healthy individuals, indicating that there are efficient immune mechanisms against fungal pathogens [3].
The mucosal surface of the respiratory tract provides a physical barrier where inhaled conidia encounter host defence mechanisms for the first time. At the epithelial surface of the upper and lower respiratory tract, conidia are trapped in mucus and removed with the help of ciliated cells. In the lower respiratory tract, alveolar macrophages represent a first-line innate host defence mechanism for clearing inhaled A. fumigatus from the lung [4–6]. It has been shown that in the alveolar space, conidia are taken up by resident macrophages and killed using reactive oxygen species (ROS) [5]. In spite of the efficiency of alveolar macrophages in removing inhaled particles, conidia sometimes germinate into hyphae, which are also recognised by macrophages. For instance, it has been shown that maturing but not resting (nongerminated) A. fumigatus conidia stimulate nuclear factor (NF)-κB in macrophages in a partially Toll-like receptor (TLR)2-dependent manner, leading to the secretion of pro-inflammatory cytokines and the production of ROS [7]. Furthermore, Steele et al. [6] showed that the β-glucan receptor dectin-1 is required for the pro-inflammatory responses of alveolar macrophages to swollen and germinating A. fumigatus conidia. Whether respiratory epithelial cells play an active role in the defence against invading fungal pathogens is not well understood. However, in recent years, it has become evident that the epithelial surface of the respiratory tract not only provides a physical barrier that is highly effective in blocking penetration by most microbes but also has the ability to recognise microbes and to initiate an immune response [8, 9]. Similar to classical immune cells, such as macrophages or neutrophils, respiratory epithelial cells are known to express a variety of pattern recognition receptors, including TLRs and NOD-like receptors (nuclear oligomerisation domain), allowing for the detection of diverse pathogens, such as bacteria and viruses [10]. Even though innate immune functions of respiratory epithelial cells are well described in the response to bacteria and viruses, little is known about the response of these cells to fungal pathogens such as A. fumigatus and its spores, the conidia. It has been shown that surfactant found in the fluids lining the epithelial surfaces of the lower respiratory tract binds conidia [11, 12] and, therefore, enhances their uptake by neutrophils and alveolar macrophages [13]. In addition, it could be shown that airway epithelial cells can internalise conidia, thereby serving as reservoirs for immune cell evasion and dissemination throughout the host [14]. Furthermore, it has been shown that germinating but not resting (nongerminated) conidia induce the expression of interleukin (IL)-8 in the human bronchial epithelial cell (HBEC) line BEAS-2B [15].
It was the aim of this study to investigate whether differentiated respiratory epithelial cells recognise A. fumigatus conidia and initiate an immune response. We demonstrate that inactivated resting conidia activate epithelial cells in an internalisation-dependent manner, resulting in the activation of the interferon (IFN)-β signalling pathway and that double-stranded RNA (dsRNA) derived from conidia is responsible for TLR3/receptor-interacting protein (RIP)-1/TANK-binding kinase (TBK)-1 signalling in epithelial cells.
METHODS
Cell culture, media and reagents
HBECs were isolated from large airways resected during surgery and cultivated as submersed or air–liquid interface (ALI) cultures as described previously [16]. The protocol was approved by the institutional review board of the University of Marburg (Marburg, Germany), and informed consent was obtained from the patients. The experiments presented in this study were performed using cells obtained from seven individual patients who underwent surgery for lung cancer. Only macroscopically and microscopically cancer free tissue was used for cell isolation. A. fumigatus (clinical isolate 27707) was cultured on YPD agar plates (15 μg·mL−1 penicillin and 40 μg·mL−1 streptomycin to prevent bacterial contamination). Conidia were obtained by flushing the A. fumigatus culture with PBS/0.1% Tween-20. The resting conidia obtained were washed with PBS three times and counted. For experiments with swollen conidia, conidia were incubated for 3 or 6 h in RPMI 1640 at 37°C. For experiments with hyphae, conidia were incubated in RPMI 1640 for 48 h and the grown hyphae were dried. The dry, weighed hyphae were suspended in PBS to obtain a concentration of 50 mg·mL−1. Resting conidia, swollen conidia and hyphae were heat inactivated (90°C for 60 min) or inactivated using ultraviolet (UV) light and a sample was cultured overnight to confirm complete inactivation. HBEC cytotoxicity was determined using an LDH-Cytotoxicity Assay Kit according to the manufacturer’s instructions (Abcam, Cambridge, UK).
Stimulation of HBEC with conidia and hyphae
HBEC cultures were stimulated apically with the indicated concentrations of conidia and hyphae. In some cases, HBECs were incubated for 1 h with the indicated concentrations of cytochalasin D (Zygosporium mansonii; EMD, Darmstadt, Germany), resveratrol (EMD), or helenalin (EMD) before stimulation. Experiments were repeated at least twice and run in triplicates.
RNA isolation and real-time RT-PCR
Total RNA of cells and conidia was isolated using the RNeasy Mini kit (Qiagen, Hilden, Germany). For conidial RNA isolation, 1×109 conidia were frozen in liquid nitrogen and ground in a mortar before RNA was isolated (1.5 μg RNA per 109 conidia). Where indicated, conidial RNA was digested with ribonuclease (RNase) I or III (New England Biolabs, Frankfurt, Germany). RNA was reverse transcribed using a cDNA synthesis kit (MBI Fermentas, St Leon-Rot, Germany). A SYBR Green PCR mix (ABgene, Schwerte, Germany), and β-actin (sense: 5′-AGCCTCGCCTTTGCCGA -3′; antisense: 5′-CTGGTGCCTGGGGCG -3′), IFN-β (sense: 5′- CAGCAATTTTCAGTGTCAGAAGC-3′; antisense: 5′- TCATCCTGTCCTTGAGGCAGT-3′) and IFN-γ-inducible protein (IP)-10 (sense, 5′-TGAAATTATTCCTGCAAGCCAA-3′; antisense: 5′-CAGACATCTCTTCTCACCCTTCTTT-3′) primers were used. Quantitative PCR results were obtained using the ΔΔCt method. Experiments were repeated at least twice and run in triplicate.
Determination of cytokine concentrations
Levels of IP-10, IFN-β and IL-8 in cell-free supernatants were determined using commercially available sandwich-type ELISAs (R&D Systems, Abingdon, UK).
Transfection experiments
Transfections were performed using TransIT LT-1 (Mirus Bio Corporation, Madison, WI, USA). For transfection of HBECs with conidial RNA, 1×105 cells were seeded in a 12-well plate. For NF-κB activation experiments, 5×104 HBECs cultivated in 24-well plates were transfected with 0.5 μg NF-κB–luciferase reporter plasmid per well. The medium was removed after 24 h and medium containing conidia or 10 ng·mL−1 tumour necrosis factor (TNF)-α, serving as positive control, was added for a further 24 h. Luminescence was measured using a Dual-Luciferase Reporter Assay System (Promega, Mannheim, Germany). Human embryonic kidney (HEK)293 cells were seeded in a six-well plate and transfected with 1 μg human (h)TLR3 plasmid or 1 μg pcDNA3.1.
Statistical analysis
Data are presented as mean±sem. Comparisons between groups were analysed by unpaired t-tests (two sided) or ANOVA for experiments with more than two subgroups. Post hoc range tests were performed with the unpaired t-test (two sided) with Bonferroni adjustment. Results were considered statistically significant for p-values <0.05.
RESULTS
A. fumigatus conidia induce internalisation-dependent IFN-β and IP-10 expression in respiratory epithelial cells
Inhaled A. fumigatus conidia can germinate, develop hyphae and cause severe disease, especially in immunocompromised individuals [2, 3]. To characterise the inflammatory response of respiratory epithelial cells to different developmental stages of A. fumigatus, differentiated primary HBECs grown as ALI cultures and conventional liquid cultures were stimulated with heat- or UV light-inactivated resting (nongerminated) conidia, swollen (germinated) conidia and hyphae of A. fumigatus. Interestingly, there was a strong and dose-dependent upregulation of IFN-β and IP-10 expression in HBECs when stimulated with heat- and UV light-inactivated resting conidia, which was not observed when the cells were incubated with swollen conidia or hyphae, as shown by real-time RT-PCR analysis (fig. 1a and b). Furthermore, figure 1c shows that the induction of IFN-β preceded the induction of IP-10, reaching statistical significance after 4 and 24 h, respectively.

Resting conidia, but not swollen conidia or hyphae, induce interferon (IFN)-β and IFN-γ-inducible protein (IP)-10 expression in epithelial cells grown as a) submerged monolayers or b) air–liquid interface (ALI) cultures. a, b) Epithelial cells were inoculated for 24 h with the indicated concentrations of heat-inactivated resting conidia, with 106 heat-inactivated swollen conidia (germinated for 3 and 6 h) per millilitre, 100 μL of heat-inactivated hypha suspension (50 mg·mL−1) or 106 ultraviolet (UV) light-inactivated resting conidia. c) Submerged monolayers were inoculated with 103 heat-inactivated resting conidia per millilitre for the indicated times. Epithelial RNA was isolated and quantitative RT-PCR was performed to measure the induction of IFN-β and IP-10. Data are presented as mean±sem of at least two different experiments each performed in triplicate and using cells from different donors. ns: nonsignificant. *: p<0.05.
It has been shown that conidia are internalised by ciliated and nonciliated epithelial cells, and are subsequently directed to the endosomal pathway [14, 17–19]. To examine whether the observed induction of IFN-β and IP-10 required internalisation of conidia by epithelial cells, HBECs were pre-incubated with the endocytic inhibitor cytochalasin D and stimulated with heat-inactivated resting conidia. The induction of IFN-β and IP-10 expression in HBECs incubated with heat-inactivated resting conidia was inhibited by cytochalasin D in a dose-dependent manner (fig. 2a). In addition, ELISA analysis revealed that the release of IP-10 into the culture supernatant was also inhibited by cytochalasin D (fig. 2b; the levels of IFN-β were below the ELISA detection limit). A lactate dehydrogenase release assay showed that the observed effects were not due to cytotoxicity (data not shown).

Cytochalasin D inhibits the induction of interferon (IFN)-β and IFN-γ-inducible protein (IP-10). Submerged human bronchial epithelial cell monolayers were inoculated with 106 heat-inactivated resting conidia per millilitre after pre-incubation with the indicated concentrations of cytochalasin D for 1 h. a) At 24 h post-inoculation, epithelial RNA was isolated and quantitative RT-PCR was performed to measure the induction of IFN-β and IP-10. b) IP-10 concentrations in supernatants were analysed by ELISA. Data are presented as mean±sem of at least two different experiments each performed in triplicate and using cells from different donors. *: p<0.05.
Together, these results demonstrate that resting (nongerminated) conidia, but not germinated conidia or hyphae, activate primary airway epithelial cells, resulting in the activation of the IFN-β signalling pathway and the release of IFN-β-inducible proteins.
HBECs recognise conidial dsRNA
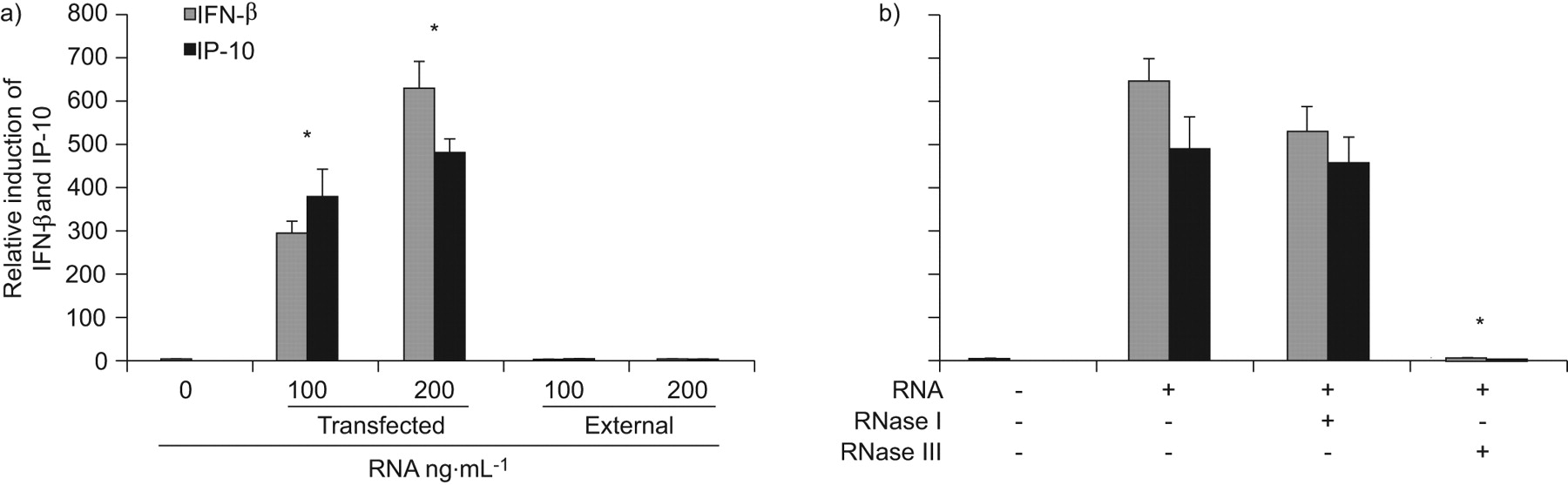
Next, we sought to determine which factors of the conidia mediate the induction of IFN-β and IP-10. As the activation of the IFN-β signalling pathway had previously been associated with foreign RNA [20], we isolated RNA from conidia and transfected it into HBECs. The conidial RNA induced IFN-β and IP-10 expression at similar levels to the heat- and UV light-inactivated resting conidia, whereas external RNA given directly into the culture medium did not induce expression of IFN-β or IP-10 (fig. 3a). To evaluate whether single-stranded RNA (ssRNA) or dsRNA is stimulatory for HBECs, native conidial RNA was digested with RNase I, which is specific for ssRNA, or RNase III, which is specific for dsRNA. Treatment of conidial RNA with RNase III resulted in a complete loss of its stimulatory activity after transfection, whereas RNase I had no effect (fig. 3b). These results show that dsRNA of conidia activates IFN-β signalling in HBECs.

Airway epithelial cells recognise double stranded conidia RNA. Submerged human bronchial epithelial cell monolayers were a) transfected with the indicated concentrations of RNA isolated from conidia or stimulated externally with conidial RNA added directly to the culture medium or b) transfected with 200 ng·mL−1 conidial RNA digested with ribonuclease (RNase) I or RNase III. At 24 h post-inoculation, epithelial RNA was isolated and quantitative RT-PCR was performed to measure the induction of interferon (IFN)-β and IFN-γ inducible protein (IP)-10. Data are presented as mean±sem of at least two different experiments each performed in triplicate and using cells from different donors. *: p<0.05.
The induction of IFN-β and IP-10 is RIP-1/TBK1 dependent
It is known that IFN-β and IP-10 expression is dependent on TIR domain-containing adapter inducing IFN-β (TRIF) that is, for instance, involved in the TLR3 signalling cascade [21]. Furthermore, the kinases RIP-1 and TBK1 are essential factors of the TRIF signalling cascade [22] and can be inhibited with resveratrol [23]. To examine whether the observed induction of IFN-β and IP-10 expression requires RIP-1 and TBK1 activity, HBECs were pre-incubated with resveratrol and stimulated with heat-inactivated resting conidia. The induction of IFN-β and IP-10 expression (fig. 4a), and release of IP-10 into the culture supernatant (fig. 4b) were inhibited by resveratrol in a dose-dependent manner.

Resveratrol inhibits the induction of interferon (IFN)-β and IFN-γ-inducible protein (IP)-10. Submerged human bronchial epithelial cell monolayers were inoculated with 106 heat-inactivated resting conidia per millilitre and incubated with the indicated concentrations of resveratrol. a) At 24 h post-inoculation, epithelial RNA was isolated and quantitative RT-PCR was performed to measure the induction of IFN-β and IP-10. b) IP-10 concentrations in supernatants were analysed by ELISA. Data are presented as mean±sem of at least two different experiments each performed in triplicate and using cells from different donors. *: p<0.05.
The transcription factor NF-κB is known to be activated within the TLR signalling cascade and is involved in the expression of many inflammatory cytokines [24, 25]. However, it has been shown that the TRIF- and IFN regulatory factor (IRF)3-mediated expression of IFN-β does not depend on the activation of the NF-κB signalling cascade, and that TLR3 activation by dsRNA results only in a delayed activation of NF-κB [22]. To test whether conidia induce NF-κB activation, HBECs were transfected with a plasmid expressing luciferase under the control of a promoter containing NF-κB binding sites. Figure 5a shows that incubation of HBECs with heat-inactivated resting conidia (106 conidia per millilitre) or TNF-α, serving as positive control, resulted in expression of luciferase, indicating that NF-κB is activated by conidia. However, inhibition of NF-κB with the NF-κB inhibitor helenalin resulted only in a partial inhibition of the induction of IP-10 expression by conidia whereas IFN-β expression was not affected at all (fig. 5b). To determine whether conidia induce the release of factors known to be under control of the NF-κB signalling cascade, we measured the release of IL-8 into the supernatant by ELISA. Only high doses of conidia (106 and 107 conidia per millilitre) induced a modest release of IL-8 by HBECs (fig. 5c).

Conidia activate nuclear factor (NF)-κB. Submerged human bronchial epithelial cell (HBEC) monolayers were a) transfected with a plasmid expressing luciferase under the control of a promoter containing NF-κB binding sites or b) incubated with the indicated concentrations of helenalin, and incubated with 106 heat-inactivated resting conidia per millilitre (a, b) or 10 ng·mL−1 tumour necrosis factor (TNF)-α (a). At 24 h post-inoculation, cells were lysed, substrate was added, and a) the luciferase activity was determined, or b) epithelial RNA was isolated and quantitative RT-PCR was performed to measure the induction of interferon (IFN)-β and IFN-γ inducible protein (IP)-10. c) Interleukin (IL)-8 concentrations in the supernatants of HBEC monolayers stimulated with the indicated concentrations of heat-inactivated resting conidia or with 10 ng·mL−1 TNF-α were analysed by ELISA. Data are presented as mean±sem of at least two different experiments each performed in triplicate and using cells from different donors. RLU: relative light units; ns: nonsignificant. *: p<0.05.
Together, these results show that RIP-1/TBK1 signalling is involved in the activation of IFN-β signalling in HBECs stimulated by conidia, whereas NF-κB signalling does not play a major role in the signalling cascade mediating the expression of IFN-β and IP-10.
TLR3 is activated by conidial dsRNA
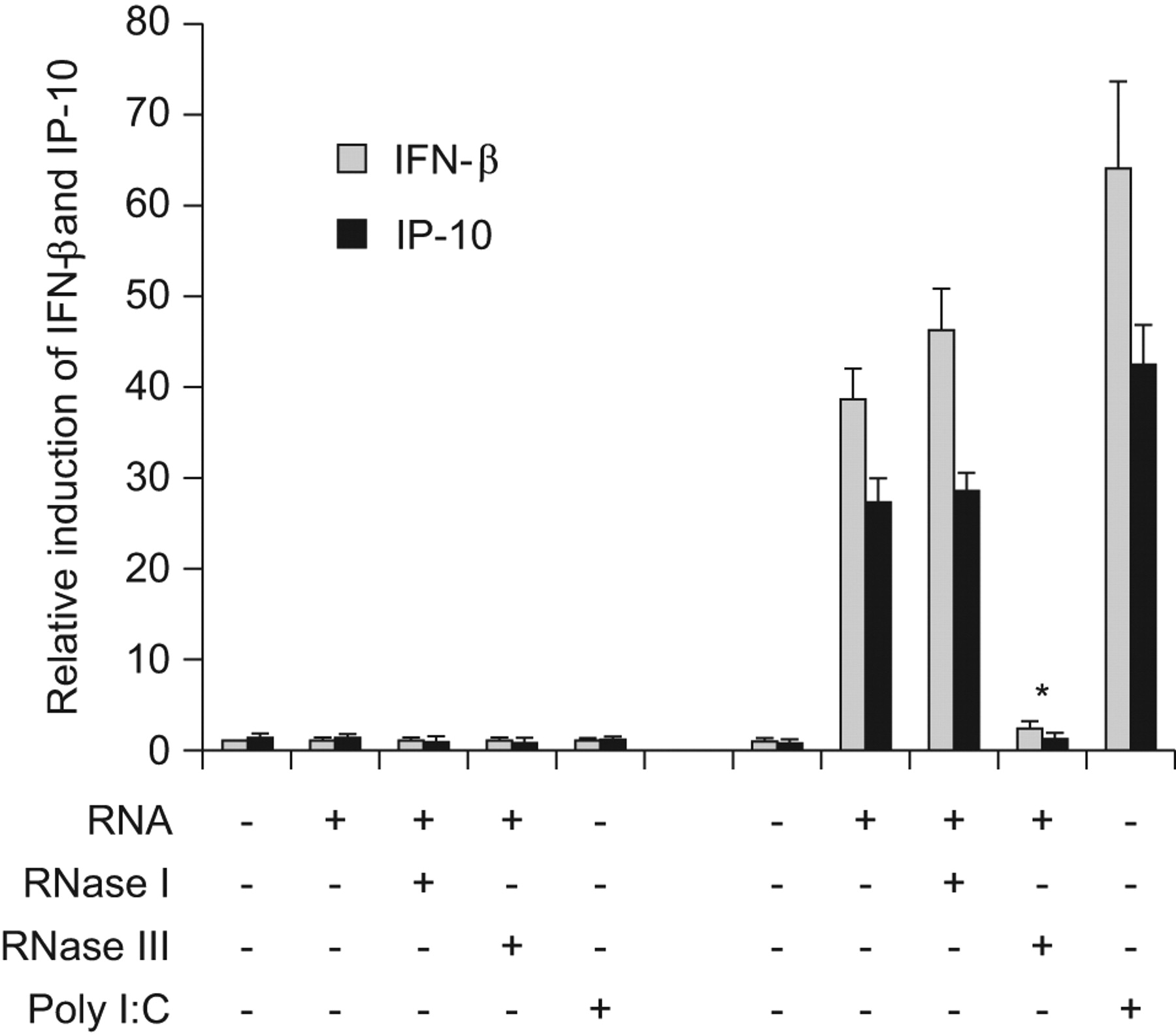
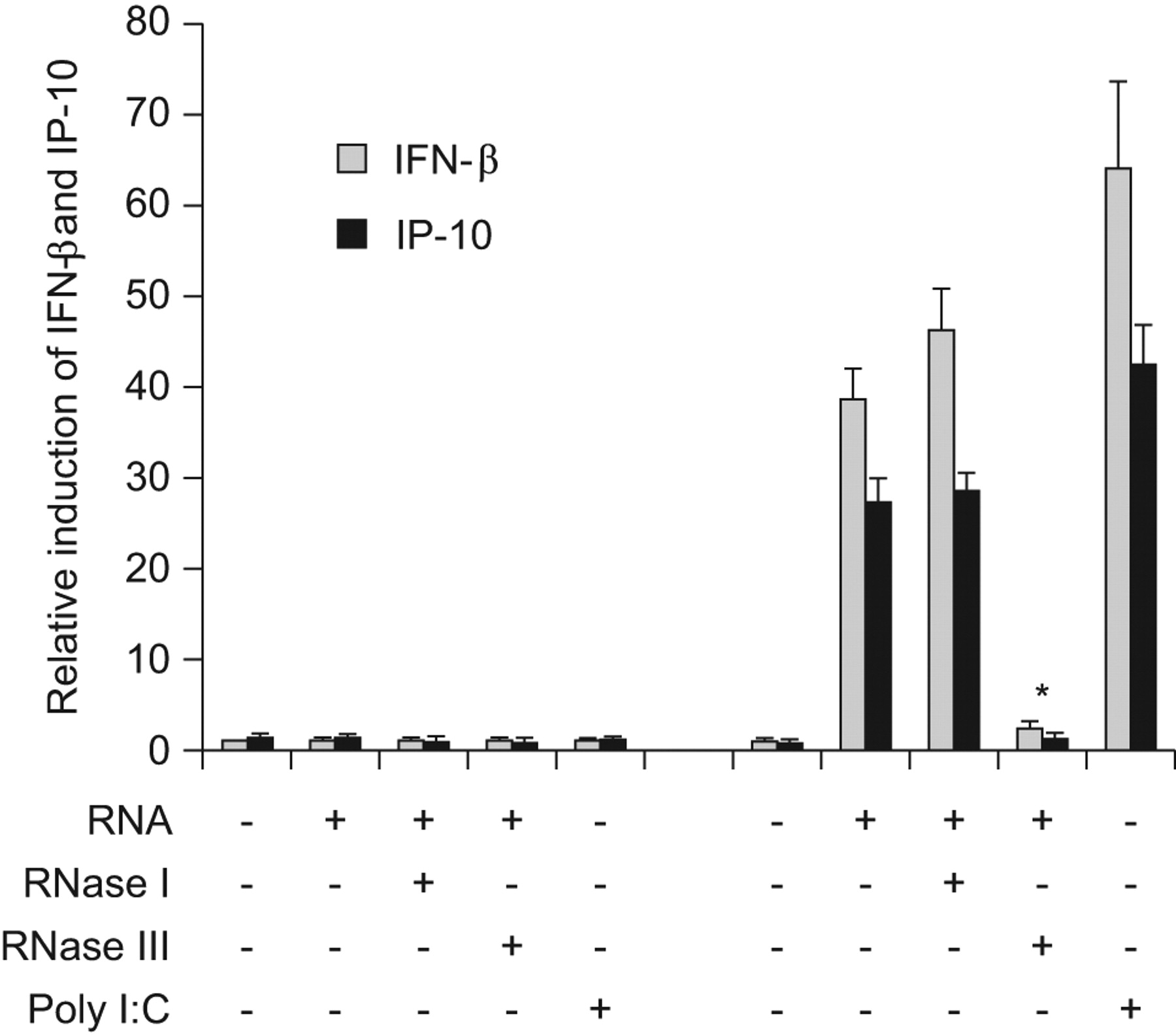
To test whether TLR3 contributes to the detection of dsRNA of conidia, HEK293 cells were transfected with a plasmid containing hTLR3 and incubated with native conidial RNA, or conidial RNA digested with RNase I or RNase III. HEK293 cells expressing hTLR3 showed a strong induction of IFN-β and IP-10 expression when stimulated with native RNA or dsRNA (RNase I-treated RNA), compared with HEK293 cells transfected with a control plasmid (fig. 6). Treatment of conidial RNA with RNase III before stimulation resulted in a complete loss of the stimulatory activity. Polyinosinic:polycytidylic acid, a specific ligand for TLR3, served as positive control.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Toll-like receptor (TLR)3 recognises double-stranded conidial RNA. Human embryonic kidney (HEK)293 cells were transfected with a plasmid expressing human TLR3 or a control plasmid, and incubated with culture medium containing 5 μg·mL−1 polyinosinic:polycytidylic acid (poly I:C) or 200 ng·mL−1 RNA isolated from conidia, either untreated, or digested with ribonuclease (RNase) I or III. At 24 h post-inoculation, HEK293 RNA was isolated and quantitative RT-PCR was performed to measure the induction of interferon (IFN)-β and IFN-γ-inducible protein (IP)-10. *: p<0.05.
DISCUSSION
This study examined the inflammatory response of differentiated airway epithelial cells to inactivated resting A. fumigatus conidia. The main finding is that airway epithelial cells are activated by resting conidia, but not by swollen conidia and hyphae, resulting in the induction of inflammatory mediators (IFN-β and IP-10) known to connect the innate and adaptive immune responses of the host [26, 27]. It is shown that conidial recognition involves the TLR3/TBK1/RIP-1 signalling cascade and that dsRNA is the pattern recognised by differentiated epithelial cells. These results give new insights into how airway epithelial cells contribute to the immune response of the host to inhaled resting conidia, and are different from mechanisms described previously for germinated conidia and hyphae [15, 28–30].
Fungal spores are potent allergens and associated with allergic and chronic diseases of the respiratory tract, such as asthma [31]. Moreover, inhaled conidia may cause life-threatening infections of the lung, such as allergic bronchopulmonary aspergillosis or invasive aspergillosis, in immunocompromised individuals. It is obvious that the host needs to clear inhaled conidia to prevent their germination and growth. Even though inhaled particles are trapped in the fluids and mucus lining the epithelial surfaces of the upper respiratory tract and are removed with the help of ciliated cells, it is evident that epithelial cells of the respiratory tract need to be actively involved in initiating and modifying the innate and adaptive immune response, as is known to occur for bacterial and viral pathogens. Indeed, previous studies have shown that ciliated and nonciliated respiratory epithelial cells, and the bronchial epithelial cell line 16HBE14o-, internalise A. fumigatus conidia [17, 19, 32, 33]. In addition, it has been shown in the epithelial cell line A549 and primary respiratory epithelial cells that internalised conidia are directed into the endosomal–lysosomal compartment where they are acidified and some are killed [18, 33]. Furthermore, it has been shown that the interaction of the respiratory epithelial cell line BEAS-2B with germinating but not resting (nongerminated) conidia results in the induction of pro-inflammatory cytokines, such as IL-8. Balloy et al. [15] showed that this induction is mediated via a signalling cascade involving phosphatidylinositol-3-kinase, p38 mitogen-activated protein kinase (MAPK) and extracellular signal-regulated kinase 1/2, independent of the MyD88 pathway. These results are consistent with a study showing that A. fumigatus induces innate immune responses in alveolar macrophages via MAPK signalling pathways independent of TLR2 and TLR4, and that recognition by TLR2/4 and MyD88 signalling are dispensable for the clearance of A. fumigatus in immunocompetent individuals [29]. In contrast, Mambula et al. [30] demonstrated that TLR2 and MyD88 mediate a response of macrophages to live resting conidia of A. fumigatus, demonstrated by the induction of TNF-α. Furthermore, a recent study showed that conidia of Histoplasma capsulatum induce IFN-β expression in murine macrophages in an IRF3-dependent manner [34]. Our finding that heat- or UV light-inactivated and, thus, metabolically inactive resting conidia are recognised by differentiated HBECs, resulting in the expression of the inflammatory mediators IFN-β and IP-10, offers new insights into the underlying signalling pathways of A. fumigatus recognition at an early state of fungal infection. Our study adds to those mentioned above that used live conidia, germinated conidia and hyphae with focus on pro-inflammatory cytokines, such as IL-8 and TNF-α, and the TLR2/4–MAPK signalling pathways [29, 30]. The results presented in this study suggest TLR3 as a receptor contributing to the activation of respiratory epithelial cells by recognising dsRNA of conidia. Our findings are consistent with a recent study showing that epithelial cells provide protection in the lungs of mice infected with A. fumigatus conidia in a TLR3/TRIF-dependent manner [35]. Previous studies also showed that TLR3 is expressed in primary respiratory epithelial cells and that TLR3 signalling is associated with IFN signalling pathways [10]. TLR3 is described as being localised to endosomal membranes where it recognises dsRNA [25, 36]. This is consistent with our finding that the activation of HBECs was dependent on endocytosis of the conidia. Furthermore, isolated conidial RNA was only able to induce IFN-β and IP-10 expression when it was transfected into HBECs, whereas no induction of IFN-β or IP-10 could be observed when the RNA was added directly to the culture medium. In addition, isolated conidial RNA strongly induced IFN-β and IP-10 in HEK293 cells overexpressing hTLR3, but not in control HEK293 cells. However, there are additional recognition mechanisms known for dsRNA involving cytosolic protein kinase (PK)R [37] and RNA helicases, such as RIG-1 and mda-5 [38, 39]. A contribution of these receptors to the activation of HBECs by conidia cannot be completely excluded. In fact, for transfected RNA, it is reasonable to suggest that cytosolic receptors for dsRNA mediate the induction of IFN-β and IP-10 independent of TLR3, as transfected RNA directly enters the cytosol of HBECs. Our finding that inhibition of RIP1 and TBK1 blocked the induction of IFN-β and IP-10 by inactive conidia excludes PKR as receptor for conidial dsRNA, as RIP1 and TBK1 are involved in TLR3 and RIG-1/mda-5 signalling, but not in PKR signalling [40]. TBK1 is essential for the induction of IRF3-regulated genes, such as the IFN-β gene [21], whereas phosphorylation of RIP-1 is involved in the late activation of NF-κB [24]. In addition, our results show that only the expression of IP-10 is partially dependent on NF-κB signalling. This is consistent with studies showing that the induction of IFN-β via the TLR3 signalling cascade does not rely on the activation of NF-κB, even though TLR3 signalling leads to a late activation of NF-κB [36]. As only dsRNA showed stimulatory activity in this study, we suggest that dsRNA derived from mycoviruses and conidial mRNA are responsible for the induction of IFN-β signalling in epithelial cells. Previous studies have shown that mycoviruses are efficiently transmitted by asexual spores containing up to 120 μg dsRNA per gramme of dry weight and that dsRNA derived from fungal mycoviruses is a potent inducer of IFN [41–44]. Furthermore, mRNA has been shown to be a potent ligand of TLR3, probably due to secondary double-stranded structures [45].
Besides its role in the defence against viruses, IFN-β plays an important role in the differentiation and activation of effector cells of the innate and adaptive immune system, such as macrophages, dendritic cells and T-cells [46]. IFN-β inhibits apoptosis of T-cells and enhances the expression of co-stimulatory factors, such as CD40 and CD86 in dendritic cells [47]. Another function of IFN-β is the induction of a group of genes called IFN response genes, including that encoding IP-10 [48], thereby linking innate and adaptive immunity [49]. IP-10 is chemotactic for monocytes/macrophages, T-cells and dendritic cells [26]. It is reasonable to conclude that the expression of IFN-β and IP-10 by epithelial cells plays an important role in directing monocytes/macrophages and other professional immune cells to the respiratory tract to ensure efficient clearance of inhaled and germinating spores. In addition, IP-10 is described as contributing to allergic pulmonary inflammation and airway hyperreactivity [27]. Thus, epithelium-derived IP-10 may contribute to the progression of diseases such as allergic bronchopulmonary aspergillosis. Our data are in contrast to a study of Aimanianda et al. [50] that demonstrated that the hydrophobic surface layer of nonswollen conidia prevents immune recognition of fungal spores. We suggest that internalisation, and lysosomal acidification and killing of conidia, as outlined here and in the Results section, might be a mechanism by which dsRNA is released from the conidia when entering the endosomal pathway in epithelial cells.
In conclusion, our study provides insight into the mechanisms allowing respiratory epithelial cells to react to resting A. fumigatus conidia. Conidial dsRNA induces the activation of the IFN-β signalling pathway and the expression of chemokines, such as IP-10, indicating an important role of the epithelium in initiating an early immune response to inhaled conidia.
Acknowledgments
We thank K. Kändler (Thomas Jefferson University, Philadelphia, PA, USA) for careful reading of the manuscript and K. Schröppel (Universität Erlangen, Erlangen, Germany) for providing the A. fumigatus (clinical isolate 27707).
Footnotes
Support Statement
This study was supported by grants from Deutsche Forschungs-gemeinschaft Transregio TR22 (TPA8) and the German Federal Ministry of Education and Research (BMBF) via 01KI07117.
Statement of Interest
None declared.
- Received June 22, 2010.
- Accepted July 6, 2011.
- ©ERS 2012
REFERENCES










