





Abstract
Epithelial-to-mesenchymal transition (EMT) has been implicated in the dysregulated epithelial wound repair that contributes to obliterative bronchiolitis (OB) after lung transplantation. Acquisition of Pseudomonas aeruginosa in the transplanted airway has been shown to be a risk factor for the development of OB. We investigated the potential of P. aeruginosa to drive EMT in primary bronchial epithelial cells (PBECs) isolated from lung transplant recipients.
Changes in the expression of epithelial and mesenchymal markers was assessed in cells challenged with clinical isolates of P. aeruginosa or co-cultured with P. aeruginosa-activated monocytic cells (THP-1) in the presence or absence of transforming growth factor (TGF)-β1.
P. aeruginosa did not drive or accentuate TGF-β1-driven EMT directly. Co-culturing P. aeruginosa-activated THP-1 cells with PBECs did not drive EMT. However, co-culturing P. aeruginosa-activated THP-1 cells with PBECs significantly accentuated TGF-β1-driven EMT.
P. aeruginosa, via the activation of monocytic cells, can accentuate TGF-β1-driven EMT. These in vitro observations may help explain the in vivo clinical observation of a link between acquisition of P. aeruginosa and an increased risk of developing OB.
- Bronchiolitis obliterans syndrome
- lung transplantation
- macrophages
- obliterative bronchiolitis
- transforming growth factor-β1
Over the last 25 yrs, lung transplantation has evolved from an experimental intervention to an accepted therapeutic option for selected patients with end-stage lung diseases. To date, ∼28,000 lung transplants have been reported to the international registry and, although early outcomes have significantly improved, long-term survival remains disappointing, being limited to a median of ∼5 yrs. The major cause of late allograft loss is due to the development of bronchiolitis obliterans syndrome (BOS) 1.
The pathological lesion of BOS is obliterative bronchiolitis (OB), which is characterised by inflammation and fibrosis in small and medium-sized airways, leading to airflow obstruction 2. The pathogenesis of the disease remains unclear, but it is hypothesised that repeated epithelial injury from both alloimmune and nonalloimmune mechanisms, and particularly a failure to re-establish an intact epithelium, contribute to fibrotic repair in the airway and disease progression 2–4. Recently, several groups have suggested a potential role for epithelial-to-mesenchymal transition (EMT) in the pathogenesis of OB 5–8. EMT is a process by which an epithelial cell alters both its phenotype and function to that of a mesenchymal cell, including acquiring the ability to produce matrix metalloproteinases and deposit extracellular matrix 9.
Transforming growth factor (TGF)-β1 is regarded as the “master switch” for fibrosis in several organs, including the lung 6, 10–14, and has been shown to be elevated in the bronchoalveolar lavage (BAL) of patients with BOS 15–17. TGF-β1 is a pleiotropic molecule, and its ability to drive EMT is dependent on both the amount of activated growth factor and the microenvironment in which it acts. There is established evidence that acute inflammatory cytokines are elevated in BAL of patients with BOS, including interleukin (IL)-8, tumour necrosis factor (TNF)-α and IL-1β 18, 19. Our group has recently shown that TNF-α and IL-1β accentuate TGF-β1-driven EMT, and cause dysregulated wound repair of injured epithelium, suggesting that a pro-inflammatory microenvironment may play a role in airway remodelling 5.
The cause of the insult responsible for injuring the epithelium is likely to be multifactorial, including oxidative stress 20, gastro-oesophageal reflux disease 21 and infections with viruses or bacteria 22–24. Recurrent bacterial infections are common in the transplanted lung and may contribute to both airway epithelial injury and repeated activation of the innate immune system. We have previously shown evidence that acquisition of Pseudomonas aeruginosa in the post-transplant lung is associated with an increased risk of BOS 22. However, how low-grade infection with P. aeruginosa leads to the airway remodelling characteristic of OB remains to be determined.
Based on these observations, we hypothesised that low-grade infection with P. aeruginosa in the transplanted lung may contribute to the airway remodelling characteristic of OB by driving or accentuating EMT.
MATERIALS AND METHODS
This study was performed in accordance with approval from the local research ethics committee (Newcastle and North Tyneside Local Regional Ethics Committee, Newcastle upon Tyne, UK) and informed written consent was obtained from all study patients.
Cell culture
Primary bronchial epithelial cells (PBECs) from stable lung transplant recipients were isolated by airway brushing at bronchoscopy, as described previously 25. Cells were cultured in Small Airway Growth Medium (Lonza, Slough, UK) on collagen coated flasks. A549 cells were maintained in Dulbecco’s modified Eagle’s medium (Sigma, Poole, UK). THP-1 cells (0.5×106 cells·mL−1) and alveolar macrophages isolated from the BAL of stable lung transplant recipients were cultured in RPMI-1640 medium (Sigma).
Co-culture experiments
THP-1 cells (1×106 cells·mL−1) activated with P. aeruginosa lysate (12.5 μL·mL−1) were added to PBECs with or without TGF-β1 (10 ng·mL−1) for 72 h and EMT was assessed, or THP-1 cells (1×106 cells·mL−1) were activated with P. aeruginosa lysate (12.5 μL·mL−1) for 6 h and then either the THP-1 cells themselves or the conditioned medium from THP-1 cells was added to PBECs with or without TGF-β1 (10 ng·mL−1) for 72 h and EMT was assessed.
P. aeruginosa whole-cell lysate preparation
P. aeruginosa whole-cell lysates were prepared from a reference strain (NCTC10662) and nine clinical isolates from our local repository of post-transplant patients (A–I). Strains were grown overnight on 1% horse blood agar plates, harvested into PBS and standardised to an optical density at 600 nm of 0.2. Bacterial suspensions were disrupted (using a Branson Digital Sonifier (Branson Ultrasonics BV, Eemnes, the Netherlands) at an amplitude of 10% for 3 min on ice) and incubated with deoxyribonuclease II (200 μg·mL−1) at 37°C for 1 h. Lysates were then treated with proteinase K (2 mg·mL−1) at 60°C for 2 h, boiled for 20 min (inactivating proteinase K) and stored at -80°C prior to use.
Flow cytometry
Following treatment, cells were pelleted and resuspended in 300 μL Phenol Red-negative RPMI media (Invitrogen, Paisley, UK). Cells were incubated at 37°C for 30 min, then 30 μL propidium iodide (1 mg·mL−1) was added prior to assessment of cell viability.
ELISA
Cytokine levels in supernatants were measured by sandwich ELISA performed with commercially available matched antibody pairs. The following cytokines were measured: IL-8, IL-1β and TNF-α (R&D Systems, Abingdon, UK).
Western Blotting
Protein concentrations were determined using the BCA protein assay kit (Perbio, Cramlington, UK).
Total cell lysates (10 μg) were separated on 4–12% bis-(hydroxyethyl)-amino-tris-(hydroxymethyl)-methane gels (Invitrogen) and electrophoretically blotted onto HyBond-P polyvinylidene difluoride (Amersham, Hatfield, UK). Membranes were incubated with primary antibodies and detected with horseradish peroxidase-labelled immunoglobulin G conjugates (Abcam, Cambridge, UK). Antibody complexes were visualised using SuperSignal West Pico chemiluminescent kit (Perbio). Results are normalised to β-tubulin as appropriate.
Trichloroacetic acid protein precipitation
Trichloroacetic acid (100% w/v) was added to culture media at a 1:4 ratio and incubated at 4°C for 10 min. The protein precipitate was pelleted by centrifugation at 14,000×g for 5 min. The protein pellet was washed twice in 200 μL cold acetone and dried by heating to 95°C for 10 min. The pellet was resuspended and separated under denaturing conditions by sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE). Membranes were incubated with primary antibodies and detected as described earlier.
Immunofluorescence
Cells fixed in 4% paraformaldehyde were incubated with primary antibodies and detected using appropriate fluorochrome-linked secondary antibodies. 4′,6-diamidino-2-phenylindole (DAPI) was used as a nuclear counterstain. Images acquired using a Leica TCS-SP-2UV microscope (Leica, Milton Keynes, UK) laser scanning confocal microscope (63× magnification).
Statistical analysis
The response of cells from each subject to a range of treatments were assessed and compared with untreated control cells. Changes in protein expression (relative band density) or protein secretion were quantified relative to untreated controls and expressed as mean±sem.
The significance of differences between groups was assessed by a one-way ANOVA using SPSS 14.0 (IBM, Feltham, UK). Differences with a p-value of <0.05 were considered statistically significant.
RESULTS
Characterisation of patient study group
Cultures of PBECs were obtained from six (two males and four females) lung transplant recipients with stable allograft function between 3 and 48 months post-transplant. No subjects showed any evidence of acute rejection (grade A2 or above, according to the International Society for Heart and Lung Transplantation classification) or infection, and were BOS stage 0 (>90% baseline forced expiratory volume in 1 s) at the time of sampling.
Investigating the effect of clinical isolates of P. aeruginosa on PBECs from lung transplant patients
PBECs stimulated with P. aeruginosa lysate showed no significant reduction in cell viability compared with untreated controls (p>0.05; n = 3; fig. 1a) and demonstrated a dose-dependent increase in the release of IL-8 up to 50 μl·mL−1 (p<0.05; n = 3), after which no significant increase was observed (p>0.05; n = 3; fig. 1b).
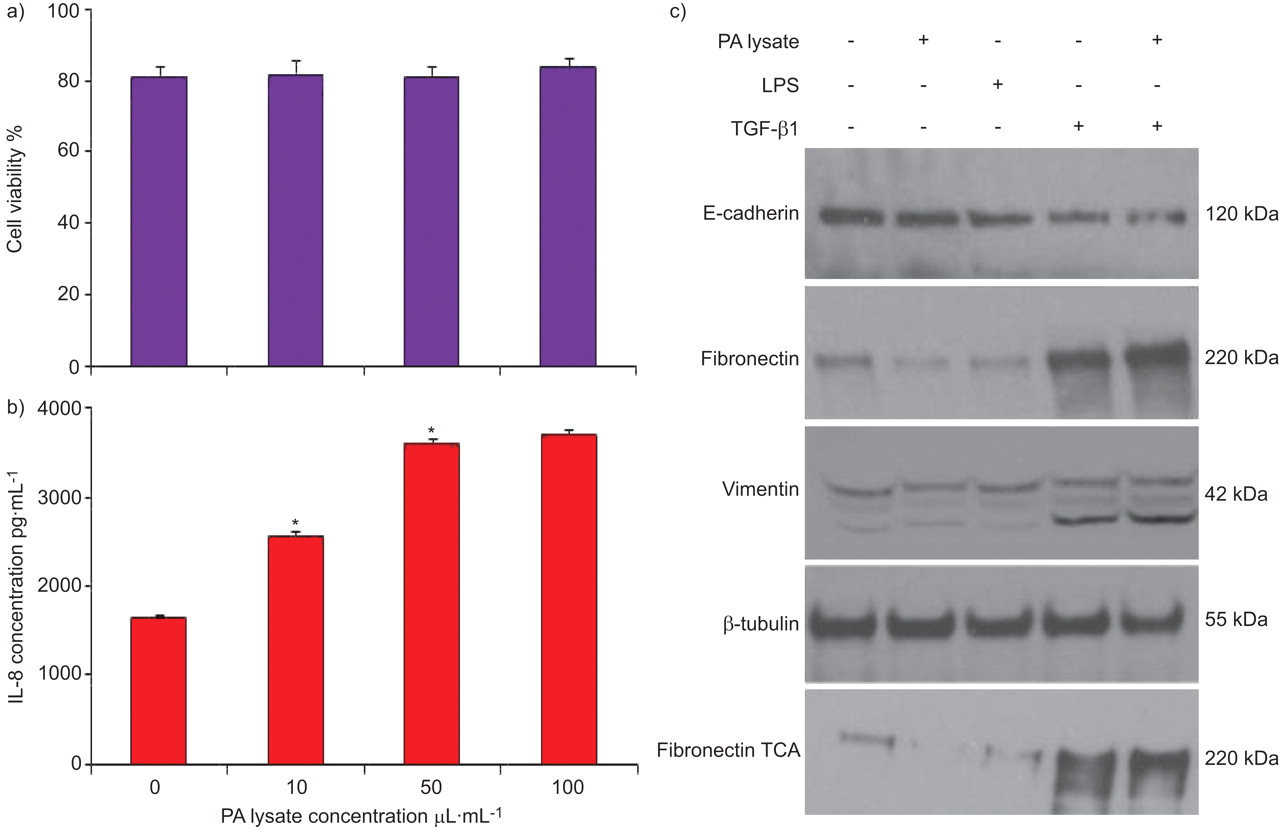
Effect of Pseudomonas aeruginosa (PA) on primary bronchial epithelial cells. Stimulation of primary bronchial epithelial cells (PBECs) with PA lysate (10, 50 or 100 μL·mL−1) had no affect on a) cell viability (p>0.05; n = 3) and induced a dose-dependent increase in b) the secretion of interleukin (IL)-8, with a maximal response at 50 μL·mL−1. (p<0.05; n = 3). c) Stimulation of PBECs with PA lysate (50 μL·mL−1) or Escherichia coli-derived lipopolysaccharide (LPS; 10 μg·mL−1) had no affect on protein expression or fibronectin deposition compared with control (p>0.05; n = 6). Treatment with transforming growth factor (TGF)-β1 (10 ng·mL−1) downregulated E-cadherin expression, increased fibronectin and vimentin expression, and increased fibronectin deposition compared with control cells (p<0.05; n = 6). Stimulation with PA lysate in the presence of TGF-β1 had no additional effect on protein expression or fibronectin deposition compared with TGF-β1 alone (p>0.05; n = 6). β-tubulin is shown as a loading control. TCA: trichloroacetic acid. Data are presented as mean±sem. *: p<0.05.
Stimulating PBECs with P. aeruginosa lysate or Escherichia coli O55 lipopolysaccharide (LPS) (Sigma) had no significant effect on epithelial phenotype (p>0.05; n = 6). In contrast, treatment with TGF-β1 (10 ng·mL−1) significantly reduced the expression of the epithelial marker E-cadherin, increased the expression of the mesenchymal markers vimentin and fibronectin, and increased the deposition of fibronectin (p<0.05; n = 6), consistent with the induction of EMT. Stimulation with P. aeruginosa lysate in the presence of TGF-β1 had no significant effect on fibronectin expression or deposition compared with TGF-β1 alone (p>0.05; n = 6; fig. 1c). These results suggest that although P. aeruginosa lysate provides pathogen-associated molecular patterns (PAMPs) to PBECs, as seen by the increased IL-8 secretion, it does not drive EMT by acting on epithelial cells directly, either alone or in synergy with TGF-β1.
Cytokine release from THP-1 cells and alveolar macrophages induced by clinical isolates of P. aeruginosa
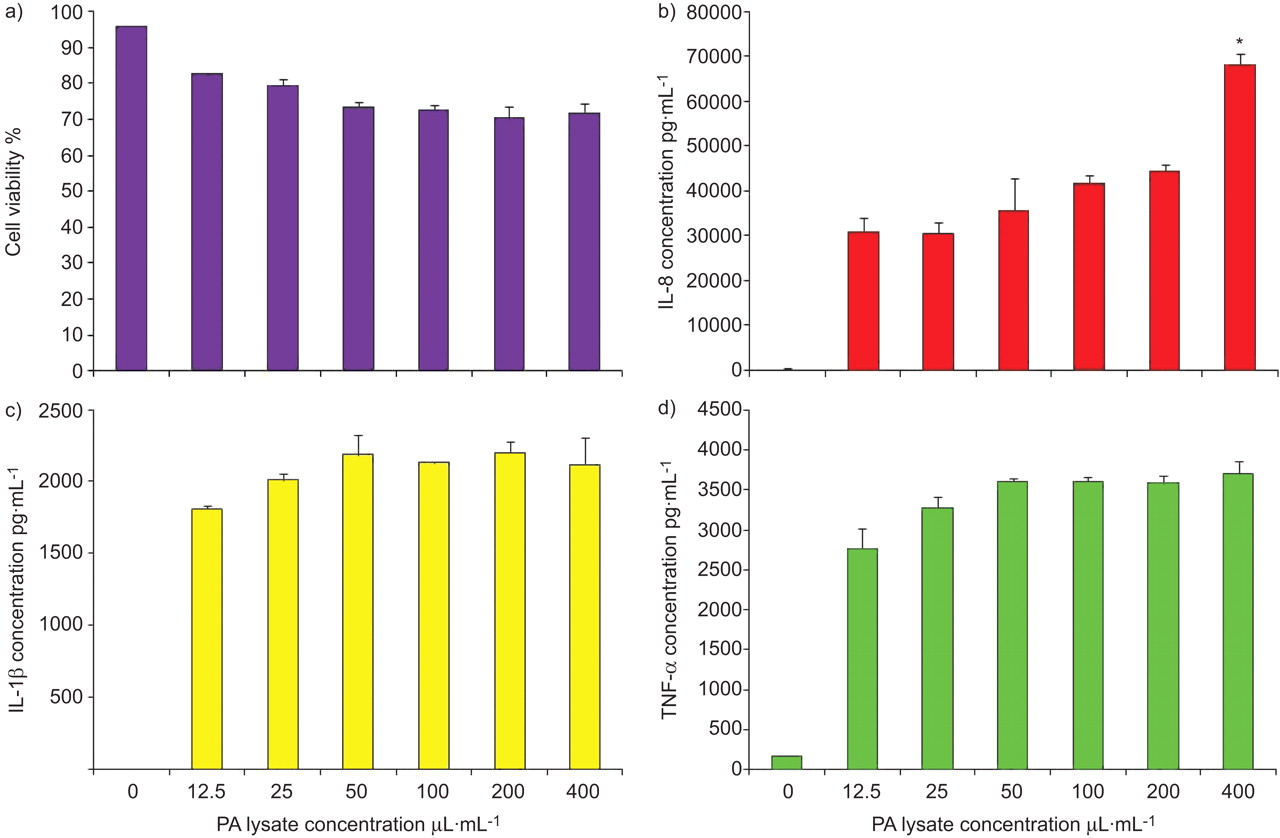
As P. aeruginosa does not directly affect EMT, we proceeded to determine if it could act indirectly via activation of immune cells and the generation of a pro-inflammatory microenvironment. Treatment of the THP-1 monocytic cell line with 12.5 μL·mL−1 P. aeruginosa lysate induced a significant increase in the release of IL-8 (fig. 2b), IL-1β (fig. 2c) and TNF-α (fig. 2d) (p<0.05; n = 3) while maintaining cell viability >80%. Concentrations >12.5 μL·mL−1 P. aeruginosa lysate induced little additional cytokine release and compromised cell viability. We therefore used a P. aeruginosa lysate concentration of 12.5 μL·mL−1 in subsequent experiments.

Cytokine release by THP-1 cells stimulated with Pseudomonas aeruginosa (PA). THP-1 cells (1×106 cells·mL−1) were stimulated with PA lysate (at 0, 12.5, 25, 50, 100, 200 or 400 μL·mL−1) for 24 h (n = 3). Cell viability was assessed by propidium iodide staining, and the production of tumour necrosis factor (TNF)-α, interleukin (IL)-8 and IL-1β was analysed by ELISA. a) Results showed a dose-dependent decrease in cell viability from 96% in control to 72% at a concentration of 400 μL·mL−1 PA lysate. b) PA lysate induced a dose-dependent increase in the release of IL-8. PA lysate induced a dose-dependent increase in c) IL-1β and d) TNF-α release up to a concentration of 50 μL·mL−1, after which no further increase was observed. Data are presented as mean±sem. *: p<0.05.
We compared the relative pro-inflammatory cytokine release from THP-1 cells in response to a laboratory reference strain of P. aeruginosa (NCTC10662) and nine clinical transplant isolates of P. aeruginosa (A–I). Stimulation with clinical isolates of P. aeruginosa induced a significantly greater secretion of IL-8 (mean±sem 68,815±4,498 versus 45,194±2,345 pg·mL−1; fig. 3a), IL-1β (1,294±83 versus 780±20 pg·mL−1; fig. 3b) and TNF-α (2,739±280 versus 1,202±20 pg·mL−1; fig. 3c) than the laboratory reference strain. Furthermore, pre-treatment of THP-1 cells with an anti-CD14 antibody (400 ng·mL−1) blocked LPS-induced TNF-α release (p<0.01; n = 3), returning levels to those comparable to control (p = 0.69; n = 3; fig. 3d). However, the anti-CD14 antibody (400 ng·mL−1) was able to inhibit only 39±3.6% (p<0.01; n = 3) of TNF-α release from THP-1 cells stimulated with P. aeruginosa lysate, suggesting P. aeruginosa is acting on multiple Toll-like receptors (TLRs) and providing us with a more representative model to that seen during infection in vivo. Hereafter, strain G was used as a representative P. aeruginosa clinical isolate.

Cytokine production by THP-1 cells stimulated with clinical isolates of Pseudomonas aeruginosa (PA) taken from patients with bronchiolitis obliterans syndrome. THP-1 cells (1×106 cells·mL−1) were stimulated with lysates of PA clinical isolates (A–I) and a lab reference strain (NCTC10662) lysate (all 12.5 μL·mL−1) for 24 h (n = 3), and the production of tumour necrosis factor (TNF)-α, interleukin (IL)-8 and IL-1β was analysed by ELISA. All clinical isolates of PA induced a significantly greater release of a) TNF-α, b) IL-8 and c) IL-1β than the lab reference strain of PA (p<0.05; n = 3). d) THP-1 cells were pre-treated with anti-CD14 antibody (at 100, 200 or 400 ng·mL−1) and stimulated with either Escherichia coli-derived LPS (100 ng·mL−1) or PA lysate (12.5 μL·mL−1) for 24 h, and the production of TNF-α analysed by ELISA (n = 3). Anti-CD14 antibody exerts a dose-dependent blocking effect on LPS-stimulated TNF-α release, returning it to control levels. There was a significant reduction in the PA-induced TNF-α release with the anti-CD14 antibody, but even at high doses, TNF-α release did not return to control levels. Data are presented as mean±sem.
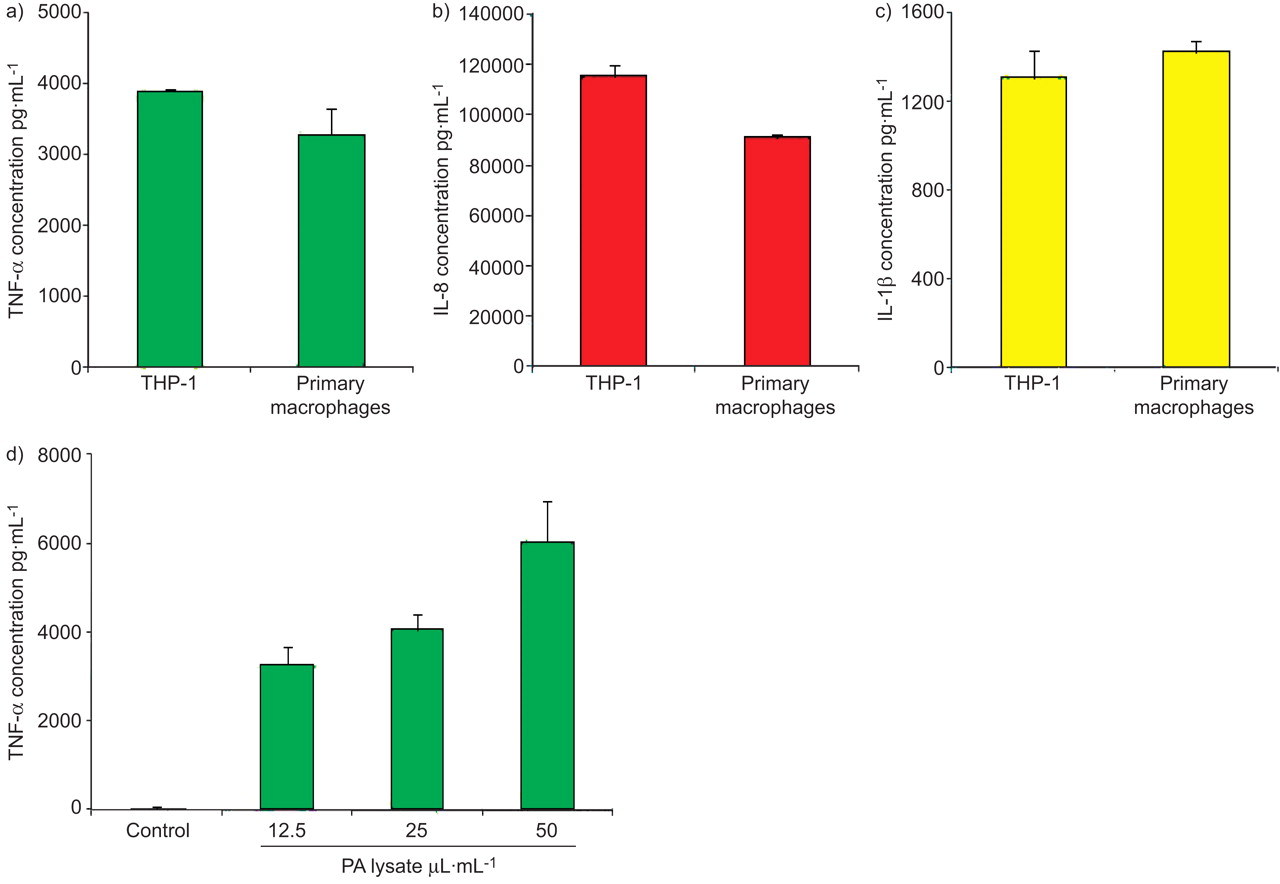
To confirm the suitability of the THP-1 monocytic cell line as a surrogate macrophage model in our co-culture system, it was important to compare the secretion of cytokines in response to activation with P. aeruginosa between THP-1 cells and alveolar macrophages isolated from the BAL of stable lung transplant recipients (fig. 4). IL-8 secretion was slightly higher in THP-1 cells compared with alveolar macrophages (116±4 versus 91±14 ng·mL−1; p = 0.02; n = 3). However, we found no significant difference in the levels of TNF-α (3,898±31 versus 3,283±372 pg·mL−1) and IL-1β (1,309±118 versus 1,425±48 pg·mL−1) secreted from THP-1 cells and alveolar macrophages, respectively (p<0.05; n = 3). We therefore proceeded to use THP-1 cells in all subsequent experiments.

Comparative assessment of cytokine release from THP-1 cells and alveolar macrophages stimulated with clinical isolates of Pseudomonas aeruginosa (PA). THP-1 cells and alveolar macrophages isolated from the bronchoalveolar lavage of stable lung transplant patients (both 1×106 cells·mL−1) were stimulated with PA lysate (12.5 μL·mL−1) for 24 h, and the secretion of a) tumour necrosis factor (TNF)-α, b) interleukin (IL)-8 and c) IL-1β was assessed by ELISA (n = 3). The secretion of all markers was similar between THP-1 cells and alveolar macrophages. d) Alveolar macrophages (1×106 cells·mL−1) were stimulated with PA lysate (at 0, 12.5, 25 or 50 μL·mL−1) for 24 h (n = 3), and the production of TNF-α was analysed by ELISA. PA lysate induced a dose-dependent increase in the release of TNF-α. Data are presented as mean±sem.
THP-1 cells stimulated with clinical isolates of P. aeruginosa accentuate TGF-β1 driven EMT
P. aeruginosa-activated THP-1 cells were co-cultured with epithelial cells to investigate the indirect effects of P. aeruginosa on EMT. Untreated cells maintain the classical “cobblestone” appearance of epithelial cells, express high levels of E-cadherin and express little to no vimentin or fibronectin. Co-culturing cells with untreated THP-1 cells or P. aeruginosa-activated THP-1 cells had no effect on morphology or EMT marker expression compared to control (p>0.05; n = 6). TGF-β1 treatment promoted a loss of cell-to-cell contact and elongation of cells, downregulation of E-cadherin expression, and an increase in vimentin and fibronectin expression (p<0.05; n = 6). Co-culturing cells with untreated THP-1 cells in the presence of TGF-β1 had no additional effect on EMT marker expression compared to TGF-β1 alone (p>0.05; n = 6). However, co-culturing cells with P. aeruginosa-activated THP-1 cells in the presence of TGF-β1 induced a more dramatic change in cell phenotype and significantly accentuated the change in EMT marker expression seen with TGF-β1 alone (p<0.05; n = 6; fig. 5). The results suggest that P. aeruginosa can accentuate EMT in an environment with elevated TGF-β1, such as that seen in the transplanted lung, but is unable to drive EMT in the absence of TGF-β1.

Accentuation of transforming growth factor (TGF)-β1-driven epithelial-to-mesenchymal transition by THP-1 cells stimulated with clinical isolates of Pseudomonas aeruginosa (PA). a) Control cells. b) Treatment with TGF-β1 (10 ng·mL−1) promotes a loss of cell-to-cell contact and elongation of A549 cells. c) Treatment with PA-activated THP-1 cells had little to no effect on epithelial morphology in the absence of TGF- β1. d) However, co-culturing with PA-activated THP-1 cells in the presence of TGF-β1 induces a dramatic change in cell phenotype to spindle-shaped myofibroblast-like cells. Pseudomonas PA-activated THP-1 cells had little to no effect on epithelial morphology in the absence of TGF-β1 (c). e) Untreated primary bronchial epithelial cells (PBECs) express high levels of E-cadherin and little to no vimentin or fibronectin. Co-culture PBECs with untreated THP-1 cells or PA-activated THP-1 cells has no effect on protein expression compared to control (p>0.05; n = 6). Treatment with TGF-β1 (10 ng·mL−1) downregulated E-cadherin expression, and increased vimentin and fibronectin expression compared with control (p<0.05; n = 6). Co-culture of PBECs with untreated THP-1 cells in the presence of TGF-β1 had no effect on protein expression compared to TGF-β1 alone (p>0.05; n = 6). However, co-culture of PBECs with PA-activated THP-1 cells in the presence of TGF-β1 further downregulated E-cadherin expression, and increase vimentin and fibronectin expression compared with TGF-β1 alone (p<0.05; n = 6).
Soluble products released from P. aeruginosa-activated THP-1 cells are responsible for accentuation of TGF-β1 driven EMT
In order to further understand how P. aeruginosa-activated THP-1 cells accentuate TGF-β1-driven EMT, the relative contribution of membrane-bound factors expressed on the surface of THP-1 cells and factors secreted from THP-1 cells on TGF-β1-driven EMT were investigated (fig. 6). Co-culturing PBECs with untreated THP-1 cells or P. aeruginosa-activated THP-1 cells had no effect on EMT marker expression compared to untreated control cells (p>0.05; n = 6). Similarly, media from untreated THP-1 cells or P. aeruginosa-activated THP-1 cells had no effect on protein expression compared to control (p>0.05; n = 6). The results suggest that the secretory products or membrane-bound factors from untreated or P. aeruginosa-activated THP-1 cells do not drive EMT in the absence of TGF-β1.

Investigation of the effect of secretory versus membrane-bound products from THP-1 cells on epithelial-to-mesenchymal transition (EMT). THP-1 cells (1×106 cells·mL−1) were incubated with Pseudomonas aeruginosa lysate (12.5 μL·mL−1) to activate the cells. After 6 h, the P. aeruginosa-activated THP-1 cells were pelleted, resuspended in fresh medium and added to primary bronchial epithelial cells (PBECs) in the presence or absence of transforming growth factor (TGF)-β1 (10 ng·mL−1), allowing us to investigate the effect of membrane-bound factors on the surface of THP-1 cells on EMT. The supernatant from the P. aeruginosa-activated THP-1 cells was also added to PBECs in the presence or absence of TGF-β1, allowing us to investigate the effect of soluble products released from THP-1 cells on EMT.
In contrast, treatment with TGF-β1 downregulated the expression of the epithelial markers cytokeratin-19 and E-cadherin, and increased the expression of vimentin and fibronectin (p<0.05; n = 6). Addition of untreated THP-1 cells or P. aeruginosa-activated THP-1 cells had no significant effect on EMT marker expression compared to TGF-β1 alone (p>0.05; n = 6).
However, treatment with medium from P. aeruginosa-activated THP-1 cells in the presence of TGF-β1 significantly accentuates the change in EMT marker expression seen with TGF-β1 alone (p<0.05; n = 6). The results suggest that secretory products released from P. aeruginosa-activated THP-1 cells, not membrane-bound factors on the surface of THP-1 cells, are responsible for accentuating TGF-β1-driven EMT in our co-culture model (figs 7 and 8).
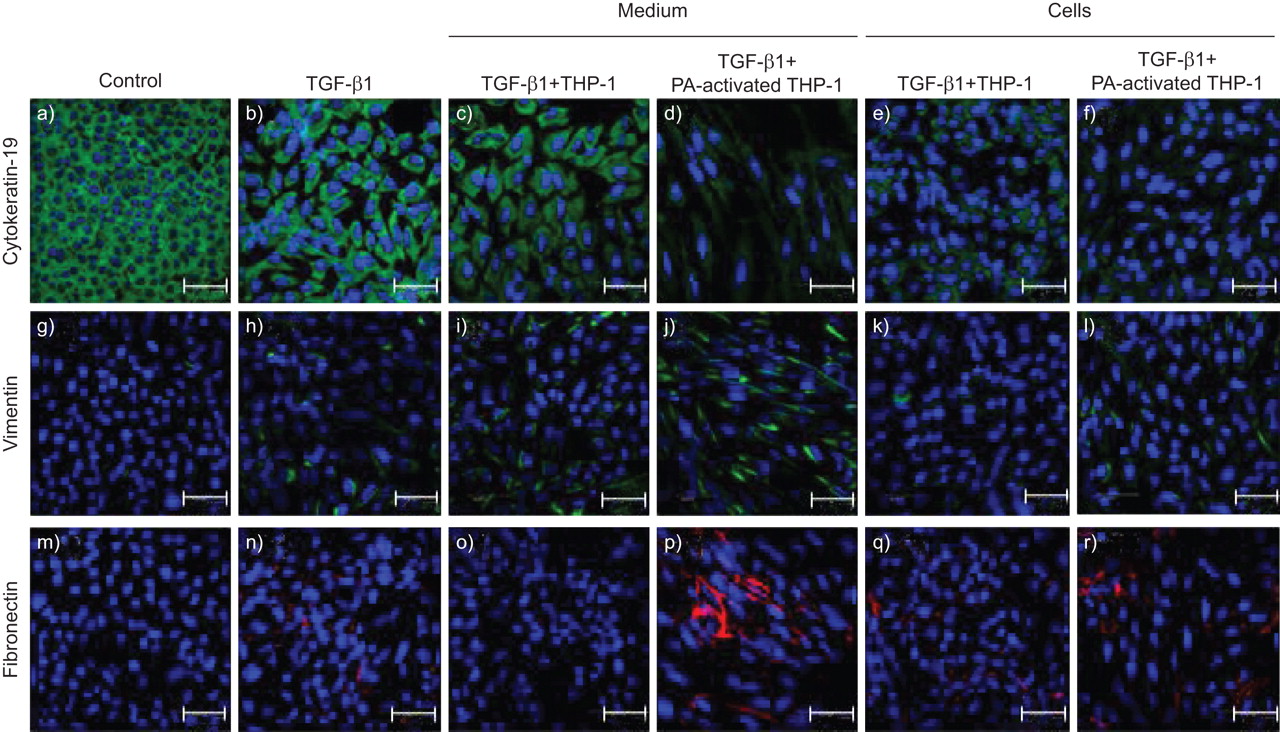
Secretory products from THP-1 cells activated with Pseudomonas aeruginosa (PA) lysates are responsible for driving epithelial-to-mesenchymal transition. Untreated primary broncial epithelial cells maintained the classic “cobblestone” morphology characteristic of epithelial cells, expressed high levels of a) cytokeratin 19, and do not express g) vimentin or m) fibronectin. Cells treated with transforming growth factor (TGF)-β1 (10 ng·mL−1) alone began to lose cell-to-cell contact, downregulated b) cytokeratin-19 expression, and increase h) vimentin and n) fibronectin expression. Addition of c, j, o) medium from untreated THP-1 cells, e, k, q) untreated THP-1 cells or f, l, r) PA-activated THP-1 cells had no affect on cell phenotype or protein expression compared to TGF-β1 alone. Addition of d, j, p) medium from PA-activated THP-1 cells accentuated the change in cell phenotype and protein expression compared to TGF-β1 alone. Images were acquired on a Leica confocal microscope (63× magnification). Scale bars = 48 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Accentuation of transforming growth factor (TGF)-β1-driven epithelial-to-mesenchymal transition (EMT) in primary bronchial epithelial cells (PBECs) is mediated by the soluble products released from activated THP-1 cells. a) Untreated PBECs express little to no vimentin or fibronectin, and express high levels of E-cadherin. Media from untreated THP-1 cells, media from Pseudomonas aeruginosa (PA)-activated THP-1 cells, untreated THP-1 cells or PA-activated THP-1 cells had no effect on EMT marker expression compared to control (p>0.05; n = 6). Treatment with TGF-β1 (10 ng·mL−1) downregulated E-cadherin expression, and increased vimentin and fibronectin expression compared with control (p<0.05; n = 6). Addition of untreated THP-1 cells or PA-activated THP-1 cells had no significant effect on EMT marker expression compared to TGF-β1 alone (p>0.05; n = 6). Addition of medium from untreated THP-1 cells increased fibronectin expression (p<0.05; n = 6) but had no significant effect on the expression of E-cadherin and vimentin compared to TGF-β1 alone (p>0.05; n = 6). Addition of media from PA-activated THP-1 cells further downregulated E-cadherin expression, and increased vimentin and fibronectin expression compared with TGF-β1 alone (p<0.05; n = 6). Changes in b) E-cadherin, c) fibronectin and d) vimentin expression (relative band density) were quantified (n = 6). Data are presented as mean±sem. *: p<0.05.
This observation was further confirmed by co-culturing epithelial cells with P. aeruginosa-activated THP-1 cells that were separated from the epithelial cells in a cell culture insert (Corning Inc., Corning, NY, USA). This technique allows only the secreted products released from THP-1 cells to affect the epithelial cells (supplementary figure E1). The results confirmed that the secreted products released from P. aeruginosa-activated THP-1 cells can accentuate TGF-β1-driven EMT (supplementary figure E2).
DISCUSSION
Viral and bacterial infections, which are common in the transplanted lung, can cause injury to the airway epithelium and activation of the pulmonary innate immune system. Such insults are believed to be important in driving crosstalk with alloimmune mechanisms and contributing to aberrant epithelial repair in the development of BOS. In particular, our group and others have shown that acquisition of the Gram-negative, opportunistic bacterium P. aeruginosa in the transplanted airway is associated with an increased risk of developing BOS 22–24. In this study, we explored the mechanism by which this organism might drive aberrant epithelial repair in the airway. Our data demonstrate that clinical isolates of P. aeruginosa cultured from the transplanted lung can, via the action of secretory products from activated immune cells, accentuate TGF-β1-driven EMT in airway epithelial cells isolated from lung transplant recipients. These in vitro findings may help explain the clinical observation that acquisition of P. aeruginosa in the transplanted airway is associated with an increased risk of developing BOS.
It is recognised that repeated sub-clinical injury and persistent inflammation in the airway epithelium coinciding with defective regeneration will favour excessive fibroproliferation, and obliteration of small and medium-sized airways in OB 26. In clinical practice, BOS is often first diagnosed (or progresses more rapidly) following a bacterial or viral infection, and these sources of acute inflammation may be very important in accentuating the processes already driving chronic lung allograft dysfunction. Our group has previously shown that TNF-α and IL-1β can cause dysregulated wound repair of injured epithelium by accentuating TGF-β1-driven EMT, demonstrating the link between acute inflammation and epithelial remodelling 5. Previous data from Vos et al. 27 suggest that even transient infection with P. aeruginosa in lung transplant recipients is associated with a significant increase in inflammation. We therefore hypothesised that acquisition of P. aeruginosa in the transplanted lung might produce sufficient inflammatory stimulus to accentuate dysregulated repair via EMT in the airway epithelium.
Our data shows that Pseudomonas aeruginosa does not drive EMT directly but can accentuate TGF-β1 driven EMT indirectly via activation of innate immune cells. Airway epithelial cells are able to respond to the presence of micro-organisms by expression of TLRs on their surface, yet these cells commonly show a hyporesponsiveness to TLR ligands 28, whereas pulmonary innate immune cells show a much more vigorous inflammatory response to TLR ligands. When activated, lung macrophages produce a multitude of growth factors and cytokines, including proteins that are present in elevated levels in the BAL of patients with BOS 18, 19, such as IL-8, TNF-α and IL-1β 29. There is growing interest in the role of the macrophage as an effector cell in allograft injury and fibrosis. In the murine heterotopic tracheal transplant model, depletion of recipient macrophages significantly abrogates obliteration of the transplanted airway 30. In this study, we used the THP-1 monocytic cell line as a model for lung innate immune cells, as the use of human macrophages isolated from either healthy controls or stable lung transplant patients was not feasible due to the large number of cells required for the co-culture experiments. Importantly, as part of the validation of our model, we showed that undifferentiated THP-1 cells show a similar inflammatory response to P. aeruginosa as alveolar macrophages isolated from the BAL of stable lung transplant recipients (fig. 4). It is also well established that THP-1 cells can be differentiated using phorbol myristate acetate into adherent macrophage-like cells 31. However, we required the THP-1 cells to remain in suspension to perform our co-culture experiments and, therefore, used undifferentiated cells.
Another important aspect of our model was the use of clinical isolates of P. aeruginosa. The whole bacterial lysates used in this study contain multiple PAMPs that signal via different TLRs and provides a more clinically relevant stimulus. This is further demonstrated by the observation that clinical isolates of P. aeruginosa induce a greater inflammatory response than the laboratory reference strain (NCTC10662).
The observations noted above suggest that targetting persistence of P. aeruginosa or its inflammatory consequences by limiting innate immune activation in the transplanted lung may be a potential therapy to limit dysregulated epithelial repair. Our group and others have shown that azithromycin treatment can reverse the decline in lung function in some patients with BOS 32–35. Azithromycin does not have any significant anti-pseudomonal activity but has been shown to exhibit extensive inhibition of the quorum-sensing systems, diminished virulence factor production and impairment of the oxidative stress response 36–38. However, recent data suggest that although azithromycin exhibits activity against P. aeruginosa biofilms, resistant mutants are readily selected, and tailored anti-pseudomonal therapies may be required to eradicate infection in patients receiving azithromycin treatment 39. This suggests that azithromycin is more likely to be exerting its benefits via its anti-inflammatory actions than by a direct action on P. aeruginosa colonisation.
Finally, it remains unclear in the literature if P. aeruginosa can be a primary driver of airway remodelling in the transplanted lung or is in fact accelerating an already established process. The data we present in this paper suggests that an elevated background level of TGF-β1 is required for P. aeruginosa to have its accentuating effects on EMT. This supports the hypothesis that P. aeruginosa may be having an accentuating effect on an already smouldering alloimmune injury to the airway epithelium.
Footnotes
↵This article has supplementary material available from www.erj.ersjournals.com
Support Statement
This work was supported by a research grant from the Medical Research Council UK (G0700861). A. De Soyza is supported by a HEFCE Senior Lectureship. A.J. Fisher is supported by a GlaxoSmithKline clinical fellowship award.
Statement of Interest
Statements of interest for A. De Soyza and A.J. Fisher can be found at www.erj.ersjournals.com/site/misc/statements.xhtml
- Received June 8, 2010.
- Accepted August 26, 2010.
- ©2011 ERS
REFERENCES










