





Abstract
The research question addressed in the current study was: do formoterol (1 and 10 nM) and montelukast (2 μM) possess interactive inhibitory effects on activated human neutrophils, particularly in relation to alterations in cyclic AMP and cytosolic Ca2+ fluxes?
Isolated human blood neutrophils were activated with the chemoattractant N-formyl-l-methionyl-l-leucyl-l-phenylalanine (fMLP) (1 μM) in combination with cytochalasin B (CB; 3 μM). Fura-2-acetoxymethyl ester-based spectrofluorimetry, lucigenin-enhanced chemiluminescence, colorimetric and flow cytometric procedures were used to measure cytosolic Ca2+ fluxes, production of superoxide, elastase release and beta-2 integrin (CR3) expression, respectively, while cyclic AMP and leukotriene (LT)B4 were assayed using competitive binding ELISA procedures.
Activation of the cells with fMLP/CB resulted in abrupt and sustained increases in cytosolic Ca2+, as well as release of elastase and production of superoxide and LTB4, and expression of CR3, all of which were attenuated by formoterol and montelukast individually, and especially by the combination of these agents. These anti-inflammatory effects of each agent, as well as the combination, were associated with significant increases in cyclic AMP.
The findings of the current study may explain the efficacy of montelukast and formoterol when used in combination with inhaled corticosteroids in the treatment of severe asthma, possibly by controlling neutrophil-driven inflammation of the airways.
Anti-inflammatory chemotherapy, most commonly inhaled corticosteroids, is the cornerstone of the pharmacotherapy of bronchial asthma 1. In patients not controlled on corticosteroids alone, the guideline recommendation is that an additional controller therapy should be added, either a long-acting β2-adrenoreceptor agonist (LABA) or a cysteinyl leukotriene receptor (cysLTR) antagonist 2. In patients who are still not adequately controlled, a combination of controller medications is added to the inhaled corticosteroids 2. With the possible exception of the effects of corticosteroids on β2-receptor expression and function 3–5, the clinical benefit of this triad of agents is thought to result from the collective effects of their distinct mechanisms of therapeutic activity, as opposed to meaningful, beneficial interactions between them. Corticosteroids possess broad spectrum anti-inflammatory activity, while LABAs and montelukast function primarily as bronchodilators and through antagonism of cysLTRs, respectively 6, 7. There is, however, evidence that LABAs and montelukast, in addition to their conventional therapeutic activities, possess anti-inflammatory properties 4, 8–10. If operative in vivo, these may complement the anti-inflammatory actions of corticosteroids in controlling severe asthma, especially those types of asthma in which the seemingly corticosteroid-resistant neutrophil 6 is the dominant type of inflammatory cell 11.
The pro-inflammatory activities of neutrophils, as well as other types of immune and inflammatory cells, are suppressed by pharmacological agents that elevate intracellular cyclic AMP (cAMP) 12. Because these cells possess β2-adrenoreceptors, they are amenable to the anti-inflammatory actions of β2-agonists, including the selective, long-acting agent, formoterol 4, 8. Montelukast has also been reported to increase intracellular cAMP by a mechanism distinct from antagonism of cysLTRs, which involves nonspecific inhibition of cyclic nucleotide phosphodiesterases (PDEs), resulting in cAMP-dependent inhibition of neutrophil pro-inflammatory activity 9. Although untested, it is plausible that the combination of formoterol and montelukast may exert augmentative, cAMP-dependent anti-inflammatory activity through the respective interactions of these agents with β2-adrenoreceptors and PDEs, especially PDE4 13.
In the current study, we have investigated the effects of formoterol and montelukast, individually and in combination, on several pro-inflammatory activities of activated neutrophils in relation to alterations in cAMP and Ca2+ handling by the cells, which is the primary focus of the study. In a limited series of experiments we have also included dexamethasone and salbutamol for comparison.
MATERIALS AND METHODS
Chemicals and reagents
Formoterol fumarate dehydrate and dexamethasone were purchased from Sigma-Aldrich (Pty) Ltd (Johannesburg, South Africa) and montelukast provided by Merck Research Laboratories (Rahway, NJ, USA), while salbutamol and rolipram were provided by GlaxoSmithKline Plc (Stockley Park, UK). With the exception of salbutamol (water soluble) all agents were dissolved to a stock concentration of 10 mM in dimethyl sulphoxide (DMSO) and used at final concentrations of 1 and 10 nM (formoterol), 2 μM (montelukast), 100 nM (salbutamol), 1 μM (dexamethasone) and 1 μM (rolipram) in the various assays of neutrophil function described below. The final concentrations of DMSO in each assay ranged from 0.1–0.2% and appropriate solvent controls were included with each experimental system.
The concentrations of montelukast used in the current study (1–2 μM) closely approximate peak serum levels of ∼1 μM attained during chemotherapy with this agent 14. The concentration of formoterol in the airways is, however, more difficult to ascertain. Assuming a maximum single dose of 24 μg and lung deposition of 18.6%, the total amount of formoterol reaching the airways would be ∼4.5 μg 15. Given a tidal volume of 500 mL, this would equate to a local concentration of 9 ng·mL−1, which is equivalent to 11 nM, close to the highest concentration of 10 nM used in the current study. Unless indicated, all other chemicals and reagents were purchased from Sigma-Aldrich.
The chemoattractant, N-formyl-l-methionyl-l-leucyl-l-phenylalanine (fMLP; 1 μM final), in combination with cytochalasin B (CB; 3 μM final) was used to activate the neutrophils. This relatively strong activator (fMLP/CB) was used because of the anti-inflammatory potency of formoterol and montelukast. An additional chemoattractant, platelet-activating factor (PAF; 200 nM) was used in an additional, limited series of experiments.
Preparation of neutrophils
The study was approved by the Faculty of Health Sciences research ethics committee of the University of Pretoria, Pretoria, South Africa and prior informed consent was obtained from all blood donors. These cells were isolated from heparinised venous blood (5 units of preservative-free heparin per mL of blood) from nonsmoking healthy adult volunteers. Each subject completed a detailed health questionnaire and underwent a health check (including measurement of blood pressure) by an experienced, qualified nursing sister prior to venepuncture. Neutrophils were separated from mononuclear leucocytes by centrifugation on Histopaque-1077 (Sigma Diagnostics, St Louis, MO, USA) cushions at 400×g for 25 min at room temperature. The resultant pellets were suspended in PBS (0.15 M, pH 7.4) and sedimented with 3% gelatin to remove most of the erythrocytes. Following centrifugation (280×g at 10°C for 10 min), residual erythrocytes were removed by selective lysis with 0.83% ammonium chloride at 4°C for 10 min. The neutrophils, which were routinely of high purity (>90%) and viability (>95%), were resuspended to 1×107 cells·mL−1 in PBS and held on ice until used.
Superoxide generation
This was measured using a lucigenin (bis-N-methylacridinium nitrate)-enhanced chemiluminescence (LECL) procedure. Briefly, neutrophils (106 cells) were pre-incubated without and with montelukast (2 μM) in 900 μL Hanks' balanced salt solution (HBSS, pH 7.4, indicator-free; Highveld Biological (Pty) Ltd, Johannesburg, South Africa) containing 0.2 mM lucigenin. Following pre-incubation, formoterol (1 or 10 nM) was added to the cells (or an equal volume of solvent to control systems) followed 20 s later by fMLP/CB, and LECL responses recorded using a Lumac Biocounter (model 2010; Lumac Systems Inc., Titusville, FL, USA). The final volume in each vial was 1 mL, and the results, which are expressed in relative light units (rlu), are the peak values for fMLP/CB-activated systems that were reached 40–50 s after the addition of the activator. There were four systems in each experiment: 1) neutrophils only (control); 2) neutrophils + formoterol; 3) neutrophils + montelukast; and 4) neutrophils + montelukast + formoterol.
Elastase release
Neutrophil degranulation was measured according to the extent of release of the primary granule enzyme, elastase. Neutrophils were incubated at a concentration of 2×106 mL−1 in HBSS without and with montelukast (2 μM) for 10 min at 37°C followed by addition of formoterol (1 or 10 nM) or solvent control to the relevant systems and 20 s later by fMLP/CB, and the tubes then incubated for 15 min at 37°C. The tubes were then transferred to an ice bath, followed by centrifugation at 400×g for 5 min to pellet the cells, and the supernatants were decanted and assayed for elastase using a micromodification of a standard colorimetric procedure. Briefly, 125 μL of supernatant were added to the elastase substrate, N-succinyl-l-alanyl-l-alanine-p-nitroanilide (3 mmol·L−1 in DMSO) in 0.05 M Tris-HCl (pH 8.0), and elastase activity was monitored spectrophotometrically at a wavelength of 405 nm. The effects of the corticosteroid dexamethasone (1 μM) on elastase release by fMLP/CB-activated neutrophils were also investigated in a more limited series of experiments. The corticosteroid was present with the cells during the 10 min pre-incubation period.
CR3 expression
Expression of CR3 on resting and fMLP/CB-activated neutrophils in the absence or presence of formoterol (10 nM) and montelukast (2 μM), individually and in combination, was measured flow cytometrically. Neutrophils (1×106 mL−1 final) were incubated in HBSS with and without montelukast for 10 min at 37°C followed by the addition of formoterol or solvent control to the relevant systems and 20 s later by fMLP/CB. The tubes were incubated for 1 min and the reaction stopped by dilution of the cells in ice-cold medium. Phycoerythrin-labelled monoclonal antibody to CD11b (Beckman Coulter, Miami, FL, USA) was added in a 10 μL volume to 1 mL of cell suspension and CR3 expression analysed using an Epics Altra Flow Cytometer equipped with a water-cooled Enterprise Laser (Beckman Coulter) and the results expressed as mean fluorescence intensity.
Spectrofluorimetric measurement of cytosolic Ca2+
Fura-2 acetoxymethyl ester (fura-2AM) was used as the fluorescent, Ca2+-sensitive indicator for these experiments 16. Neutrophils (1×107 mL−1) were incubated with fura-2AM (2 μM) for 30 min at 37°C in PBS, washed and resuspended in HBSS. The fura-2-loaded cells (2×106 mL−1) were then pre-incubated for 10 min at 37°C without and with montelukast (2 μM) after which they were transferred to disposable reaction cuvettes which were maintained at 37°C in a Hitachi 650 10S fluorescence spectrophotometer with excitation and emission wavelengths set at 340 and 500 nm, respectively. After a stable baseline was obtained (±1 min), formoterol (10 nM only) or solvent control was added to the relevant systems followed 20 s later by fMLP/CB, and alterations in fluorescence intensity monitored over a 5–10 min time course. Cytosolic Ca2+ concentrations were calculated as described previously 16. In a more limited series of experiments, the following were also investigated: 1) the effects of the short-acting β2-receptor agonist, salbutamol (100 nM final, predetermined in preliminary experiments) without and with montelukast (1 μM) on PAF (200 nM)-activated increases in cytosolic Ca2+ concentrations in neutrophils; 2) the effects of formoterol (10 nM) alone or in combination with the prototype PDE4 inhibitor rolipram (fixed, final concentration of 1 μM); and 3) the effects of dexamethasone (1 μM) on fMLP/CB-activated Ca2+ fluxes in neutrophils. Montelukast, rolipram and dexamethasone were present with the cells during the 10 min pre-incubation period, while salbutamol or formoterol were added 20 s prior to the activation of the cells.
Measurement of leukotriene B4 and cAMP
Competitive binding immunoassay procedures (Correlate-EIATM; Assay Designs Inc., Ann Arbor, MI, USA) were used to measure leukotriene (LT)B4 in the supernatants of fMLP/CB-activated neutrophils, while cAMP was assayed in the extracts of unstimulated cells in the absence and presence of formoterol and montelukast, individually and in combination. In the case of LTB4, neutrophils (2×106 mL−1) were pre-incubated for 10 min at 37°C in the absence and presence of montelukast (2 μM), after which formoterol (1 or 10 nM) was added to the relevant systems, followed 20 s later by fMLP/CB. Incubation was terminated 3 min later by the addition of an equal volume of ice-cold HBSS to the tubes, which were then held in an ice bath prior to pelletting the cells by centrifugation. The cell-free supernatants were then diluted (1:8) and assayed for LTB4, with the results expressed as pg LTB4 per 107 cells. As with the other assays of neutrophil pro-inflammatory activity, each experiment consisted of four systems: 1) neutrophils only (control); 2) neutrophils + formoterol; 3) neutrophils + montelukast; and 4) neutrophils + formoterol + montelukast. In a more limited series of experiments, the following were also investigated: 1) the effects of salbutamol (100 nM) without and with montelukast (1 μM) on the production of LTB4 by PAF (200 nM)-activated neutrophils; and 2) the effects of dexamethasone (1 μM) on the production of LTB4 by neutrophils activated with fMLP/CB.
In the case of cAMP, neutrophils (2×106 mL−1) were pre-incubated for 10 min at 37°C without or with montelukast (2 μM), after which formoterol (10 nM only) was added to the relevant systems and the reactions terminated 20 s later by the addition of an equal volume of ice-cold HBSS to the tubes. Following centrifugation, the supernatants were discarded and cAMP extracted from the cell pellets by addition of 0.1 M HCl for 15 min followed by centrifugation to remove the cell debris and the supernatants decanted and assayed for cAMP. These results are expressed as pmol cAMP per 107 cells.
Statistical analysis
With the exception of the results of the fura-2 fluorescence experiments, some of which are presented as representative traces, the results of each series of experiments are presented as the mean±sem values, in which n equals the number of different donors used in each series of experiments, with the number of replicates for each drug concentration and drug-free control system for each experiment shown in the figure legends. Levels of statistical significance were determined by comparing the absolute values for each drug-treated system with the corresponding values for the relevant drug-free control systems for each assay using repeated measures ANOVA with appropriate multiple comparisons post-test.
RESULTS
Superoxide production, elastase release and LTB4
These results for neutrophils activated with fMLP/CB in the absence and presence of formoterol (1 and 10 nM) and montelukast (2 μM) individually and in combination are shown in figure 1. Treatment of neutrophils with formoterol or montelukast significantly (p<0.05) attenuated the production of superoxide and LTB4 in particular, as well as the release of elastase, with the combination, especially in the case of 1 nM formoterol, being significantly (p<0.05) more effective in most cases, albeit additive rather than synergistic, than either of the individual agents. The exception was superoxide production, for which the effects of the combination differed only modestly from those of montelukast alone. The magnitude of inhibition of elastase release observed in the presence of montelukast, although statistically significant, was less than we had reported previously 9, probably because of the higher concentration of CB used in the current study.

Effects of formoterol (1 and 10 nM) and montelukast (2 μM) individually and in combination on the generation of superoxide (a), release of elastase (b), and production of leukotriene (LT)B4 (c) following activation of neutrophils with N-formyl-l-methionyl-l-leucyl-l-phenylalanine (fMLP) in combination with cytochalasin B (CB). For purposes of comparison, the data are presented as the mean±sem per cent of control of a minimum of four different experiments using cells from four different donors. #: p<0.05 for comparison of the formoterol and montelukast combination with the system treated with montelukast alone; *: p<0.05 for comparison of the formoterol and montelukast combination with the system treated with formoterol alone. The absolute values for the responses of control (drug-free), unstimulated neutrophils for superoxide generation, release of elastase and production of LTB4 were 3,782±475 rlu, 34±2 milli-units enzyme per 107 cells, and 37±5 pg LTB4 per 107 cells, respectively; the corresponding values for the control systems activated with fMLP/CB were 54,954±1,926 rlu, 980±16 milli-units enzyme per 107 cells and 7,926±1,170 pg LTB4 per 107 cells.
Treatment of neutrophils with dexamethasone had minimal effects on the production of LTB4 and release of elastase from fMLP/CB-activated neutrophils. In the case of elastase the values for cells activated with fMLP/CB in the absence and presence of dexamethasone were 950±15 and 932±15 milli-units elastase per 107 cells, respectively. The corresponding values for LTB4 release were 7,213±110 and 7,242±150 pg LTB4 per 107 cells (data from three separate experiments). The effects of salbutamol (100 nM) and montelukast (2 μM) individually and in combination on the release of LTB4 by PAF-activated neutrophils were also investigated. The results for unstimulated cells and for PAF-activated control cells and cells treated with salbutamol alone, montelukast alone, or the combination of these agents were 24±2, 681±29, 335±55, 132±38 and 65±17 pg LTB4 per 107 cells, respectively (p<0.05 for comparison of each drug-treated system with the control system and for the combination with the individual agents).
CR3 expression
The level of CR3 expression for resting neutrophils was 27±2 mean fluorescence intensity. The corresponding levels of expression for control, fMLP/CB-activated cells and for the corresponding systems treated with formoterol (10 nM) only, montelukast (2 μM) only, or the combination of these two agents were 92±2, 71±3, 81±2 and 60±3 mean fluorescence intensity respectively. The levels of CR3 expression were significantly less (p<0.05) than those of the control system for all three drug-treated systems, while the levels of CR3 expression in the system treated with both formoterol and montelukast was significantly less (p<0.05) than that of the systems treated with the individual drugs.
cAMP
These results are shown in figure 2. Treatment of neutrophils with either formoterol (10 nM) for 20 s, or montelukast (2 μM) resulted in significant (p<0.05) elevations in intracellular cAMP. Although higher than that observed with the individual agents, the increase in cAMP which resulted from treatment of the cells with the combination of montelukast and formoterol was not significantly different from that observed with the individual agents (fig. 2). We have previously reported that montelukast potentiates the increase in cAMP following exposure of neutrophils to salbutamol 9.
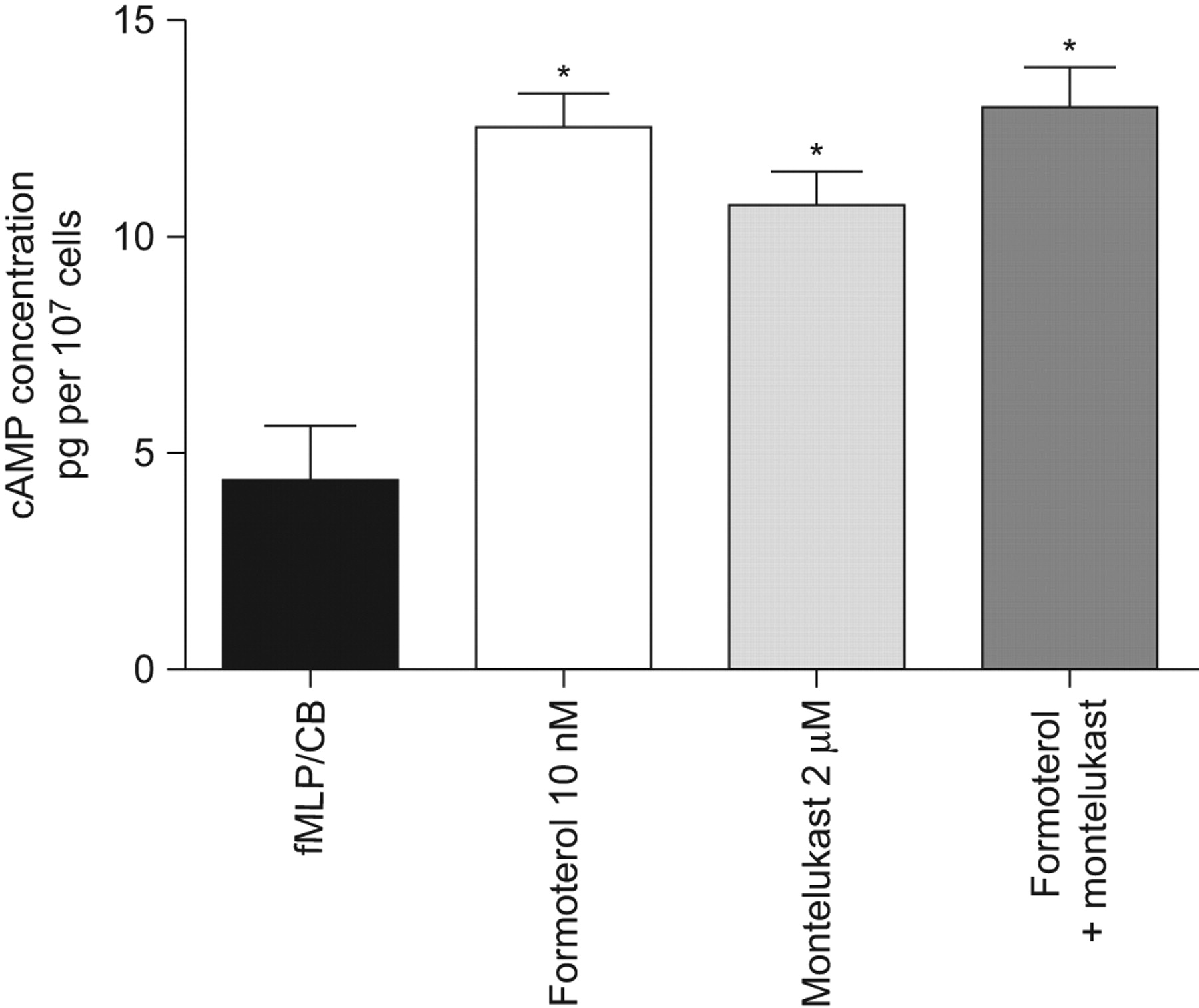
Effects of formoterol (10 nM) and montelukast (2 μM) individually and in combination on neutrophil intracellular cyclic AMP (cAMP) levels. The results are expressed as the mean±sem values (data from five different experiments using cells from five different donors). Exposure of the cells to either formoterol or montelukast was accompanied by significant (p<0.05) elevations in cAMP, while the effects of the combination of agents, although greater, did not differ significantly from those of the individual agents. fMLP: N-formyl-l-methionyl-l-leucyl-l-phenylalanine; CB: cytochalasin B. *: p<0.05.
Cytosolic calcium
The alterations in cytosolic Ca2+ concentrations in fMLP/CB-activated neutrophils were quantified using the strategy represented schematically in figure 3. The results shown in figure 4 are traces from a representative experiment which depicts the alterations in cytosolic Ca2+ (fura-2 fluorescence) following activation of the cells with fMLP/CB in the absence and presence of formoterol (10 nM) and montelukast (2 μM) individually and in combination. Activation of the cells with fMLP/CB resulted in an abrupt increase in cytosolic Ca2+, with the peak response being sustained for 1–2 min, followed by a gradual subsidence which levelled off within 1–2 min reaching a sustained plateau at a level considerably higher than the pre-activation, basal value for cytosolic Ca2+. Treatment of the cells with either montelukast (2 μM) for 10 min or formoterol (10 nM) for 20 s prior to the addition of fMLP/CB caused partial, but significant (p<0.05) attenuation of the peak, plateau cytosolic Ca2+ response, while accelerating the rate of decline, and decreasing the magnitude of the second, sustained plateau response. Again, the combination of montelukast and formoterol was found to be significantly (p<0.05) more effective than the individual agents in promoting clearance of Ca2+ from the cytosol of fMLP/CB-activated neutrophils. These results are shown in table 1. Similar results (not shown) were obtained with the nonselective PDE inhibitor rolipram (1 μM), in the presence or absence of formoterol (10 nM). Addition of PAF (200 μM) to neutrophils resulted in an abrupt increase in cytosolic Ca2+, which was followed by a sustained plateau phase and subsequent gradual decline towards basal levels. Pretreatment of neutrophils with salbutamol (100 nM) 20 s prior to addition of the stimulant did not alter the magnitude of the initial peak response, or the duration of the sustained plateau phase. However, the rate of decline in cytosolic Ca2+ concentrations was accelerated. In the presence of montelukast (1 μM), the initial plateau phase was markedly attenuated and the rate of decline in cytosolic Ca2+ concentrations increased significantly. The combination of salbutamol and montelukast was more effective than either agent alone. These results are shown in figure 5 and table 2. Dexamethasone did not detectably alter Ca2+ fluxes in fMLP/CB-activated neutrophils (results not shown).

Schematic representation of the strategy used to measure the peak cytosolic Ca2+ concentration, duration of the peak plateau phase and magnitude of the second sustained increase in cytosolic Ca2+ concentration in activated neutrophils.

N-Formyl-l-methionyl-l-leucyl-l-phenylalanine (fMLP)/cytochalasin B (CB)-activated fura-2 fluorescence responses of control neutrophils and cells treated with formoterol (10 nM) and montelukast (2 μM) individually and in combination. fMLP/CB was added as indicated by arrows when a stable baseline was obtained (±1 min). The traces are from an individual representative experiment (four in the series using cells from four different donors).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of salbutamol (100 nM; – – – –) and montelukast (1 μM; · · · · ·), alone and in combination (- - - - -), on the cytosolic Ca2+ concentrations of platelet-activating factor-activated (arrow) neutrophils. –––––: controls.
DISCUSSION
Add-on therapy with montelukast has been reported to confer benefit on patients whose asthma is poorly controlled with inhaled corticosteroid monotherapy, or with the combination of a LABA and an inhaled steroid 17–22. In this setting, it is believed that montelukast targets inflammatory mechanisms, specifically those orchestrated by cysteinyl leukotrienes and their receptors, thereby complementing the therapeutic actions of inhaled corticosteroids and β2-agonists. However, this may not be the only mechanism by which add-on therapy with montelukast confers benefits on patients with poorly controlled asthma. The recently described effects of montelukast on PDEs 9 raise the possibility that this agent may act in concert with the other anti-asthma agents, particularly β2-agonists, to augment cAMP-mediated anti-inflammatory activity and, possibly, bronchodilatory activity.
To test this hypothesis, we investigated the effects of formoterol (1 and 10 nM) and montelukast (1–2 μM), individually and in combination, on the pro-inflammatory activities of human neutrophils. Treatment of neutrophils with either agent resulted in significant inhibition of the generation of superoxide in particular, as well as expression of CR3 and release of elastase following activation of the cells with fMLP/CB, with the combination being more effective than the individual agents, most notably in the case of CR3 expression and elastase release, the effects being additive as opposed to synergistic. In asthma, phagocyte-derived reactive oxygen species and elastase have been identified as possible mediators of bronchial hyperactivity and obstruction, and airway remodelling, respectively 23, 24, while interference with CR3 expression will restrict the migration of neutrophils into the airways. In the case of production of LTB4 by activated neutrophils, the individual agents were found to be extremely potent inhibitors of production of this eicosanoid with small, but nevertheless significant, augmentative effects observed with the combination of formoterol and montelukast. Notwithstanding the well-recognised involvement of cysteinyl leukotrienes in asthma, it is noteworthy that LTB4 also appears to play an important role in the pathogenesis of severe persistent asthma, as well as aspirin- and exercise-induced asthma, allergic rhinitis and atopic dermatitis 25. Moreover, corticosteroids, which generally do not affect LTB4 production by immune and inflammatory cells, have been reported to upregulate the expression of the BLT1 receptor on corticosteroid-resistant cells such as neutrophils, monocytes and effector memory CD8+ T-cells 24–27. In the current study, dexamethasone, an agent that does not elevate intracellular cAMP concentrations, had no detectable effects on either elastase or LTB4 release by activated neutrophils.
Although of limited relevance in the treatment of chronic asthma, or asthma in which inflammation is well-controlled, we also measured the effects of the short-acting β2-agonist, salbutamol, on LTB4 production by neutrophils activated with PAF, a less potent stimulus than fMLP/CB. Salbutamol also inhibited the production of LTB4 by PAF-activated neutrophils, an effect that was augmented by inclusion of montelukast, probably by interactive cAMP-dependent mechanisms as described previously 9.
To probe the mechanism of formoterol/montelukast-mediated inhibition of the pro-inflammatory activity of neutrophils, we investigated the effects of these agents on alterations in cAMP and cytosolic Ca2+. Exposure of neutrophils to either formoterol or montelukast was accompanied by substantial increases in cAMP. In the case of montelukast, these were higher than we had reported previously 9. This is most probably attributable to the longer exposure of the cells to the drug (10 min in the current study compared to 5 min in the previous study). Although the magnitude of the increase in cAMP observed with the combination was slightly higher than that observed with the individual agents, the difference did not achieve statistical significance. This may be due to the fact that we used a relatively brief exposure time (20 s) at which the effects of formoterol were maximal; longer exposure times may have revealed significant effects of the combination.
In neutrophils, the anti-inflammatory actions of agents that elevate intracellular cAMP are achieved primarily by activation of cAMP-dependent protein kinase (PKA). PKA in turn accelerates restoration of Ca2+ homeostasis and downregulation of pro-inflammatory activities following exposure of the cells to receptor-linked, Ca2+ mobilising stimuli by several mechanisms, including: phosphorylative inactivation of phospholipase C 28; inactivation of inositol triphosphate receptors on intracellular Ca2+ stores 29; upregulation of the Ca2+ sequestering/resequestering endo-membrane Ca2+-ATP 30; and inhibition of p38 MAP kinase, resulting in interference with the activation of 5-lipoxygenase 31 and attenuation of an autocrine, LTB4-mediated secondary wave of Ca2+ uptake by the cells 32.
In the current study, formoterol and montelukast, in keeping with previous studies 8, 9, facilitated the clearance of Ca2+ from the cytosol of neutrophils activated with fMLP/CB without affecting mobilisation of the cation from intracellular stores. The two agents were most effective when used in combination. These observations are compatible with a mechanism whereby formoterol and montelukast, by targeting β2-adrenoreceptors and PDEs, respectively, exert their individual and combined anti-inflammatory activities via cAMP-dependent, accelerated restoration of Ca2+ homeostasis. This contention is supported by the following observations: 1) exposure of fMLP/CB-activated neutrophils to the type 4 PDE inhibitor rolipram, in combination with formoterol, resulted in decreases in cytosolic Ca2+ concentrations similar to those observed with montelukast and formoterol; and 2) montelukast potentiated the effects of salbutamol on clearance of Ca2+ from the cytosol of PAF-activated neutrophils. In keeping with its lack of effects on the release of elastase and LTB4, dexamethasone did not affect Ca2+ fluxes in fMLP/CB-activated neutrophils.
In conclusion, the results of the current study identify an interactive, cAMP-dependent mechanism by which formoterol and montelukast may support the anti-inflammatory actions of inhaled corticosteroids by targeting the intransigent neutrophil in patients with poorly controlled asthma.
Footnotes
Support Statement
This study was supported, in part, by a research grant from the Investigator Initiated Studies Programme, Merck Research Laboratories, Rahway, NJ, USA.
Statement of Interest
Statements of interest for C.M. Gravett, C. Feldman and R. Anderson, and for the study itself, can be found at www.erj.ersjournals.com/site/misc/statements.xhtml
- Received October 6, 2009.
- Accepted April 5, 2010.
- ©ERS 2010
REFERENCES










