





Abstract
Pleiotropic effects of statins have been reported to include inhibition of matrix metalloproteinase (MMP) release from macrophages and endothelial cells. We evaluated whether statins would inhibit MMP release from human lung fibroblasts, which play a major role in remodelling processes.
Monolayer and three-dimensional (3D) collagen gel cultures of fibroblasts were used. Cytokines (tumour necrosis factor-α and interleukin-1α) were used to induce MMP release and mRNA expression. Collagen degradation induced by cytokines and neutrophil elastase (NE) was evaluated by quantifying hydroxyproline. Atorvastatin inhibited MMP-1 and -3 release and mRNA expression in both culture systems. Similar results were obtained with simvastatin and fluvastatin.
In 3D cultures where cytokines also stimulated MMP-9 release, atorvastatin also inhibited MMP-9 release. In 3D cultures, cytokines together with NE induced collagen degradation, which was also inhibited by atorvastatin. The effect of atorvastatin was reversed by mevalonate and geranylgeranyl-pyrophosphate but not by farnesyl-pyrophosphate.
The current data suggest that statins may modulate remodelling processes mediated by fibroblasts by inhibiting MMP release.
Chronic obstructive pulmonary disease (COPD) is expected to become the third leading cause of death worldwide by the year 2020 1. The defining feature of COPD is loss of expiratory airflow that can result from at least two important structural alterations in the lungs, emphysema with its destruction of alveolar wall and loss of lung elastic recoil, and peribronchial fibrosis, which narrows airways. At present, therapy can partially alleviate symptoms, but has relatively little impact on the progressive structural alterations that compromise lung function 2. Novel therapeutic strategies that have the potential to alter lung structure, therefore, are of interest as possible approaches to alter the long-term natural history of COPD. One type of therapy that is attracting interest for its potential in COPD is the use of statins, which have been associated with reduced acute events and mortality 3–5, as well as with reduced decline in lung function in retrospective database studies 6.
Statins are a class of cholesterol-lowering drugs that inhibit 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA). Recently, pleiotropic effects of statins have been reported, including anti-inflammatory and antioxidant effects 7, 8, as well as the inhibition of matrix metalloproteinase (MMP) release from vascular cells and macrophages 9–11. One potential mechanism for statin efficacy in COPD could be effects on inflammatory cells. Such effects could be a mechanism that could alter both acute events and long-term progression. An alternate mechanism, which could be either independent or interactive, is the effect of statins on MMPs. Through these effects, statins could directly modify lung extracellular matrix (ECM) and, thereby, could modify the alterations in lung structure that compromise lung function.
The MMPs are a large family of proteolytic enzymes 12 that are produced by both inflammatory cells and structural cells in the lung including alveolar epithelial cells and fibroblasts 13. Fibroblasts are believed to be major cells responsible for maintenance and repair of ECM 14. This is accomplished by the production and assembly of ECM macromolecules, by the release of growth factors for other cells and by the release of proteolytic enzymes, particularly MMPs. This allows fibroblasts to modulate both ECM production and destruction, as well as the response of other cells.
The current study was designed to determine if statins could modulate MMP release by human lung fibroblasts. To investigate this, we utilised two types of lung fibroblasts, the widely used human fetal lung fibroblast strain (HFL-1) 15, which allows our results to be compared with those of other investigations that have used identical or similar strains and, in addition, fibroblasts from normally appearing adult lung tissue that was removed during surgery. We have, moreover, used three statins, atorvastatin, fluvastatin and simvastatin, to assure that the effects were not specific for a single agent. In addition, to determine if the effect of statins could modulate ECM degradation in addition to MMP release and to determine if the results obtained in the conventionally used monolayer culture would be observed in other culture conditions, the effect of atorvastatin was evaluated using three-dimensional (3D) collagen gels in which collagen degradation was induced by cytokines and neutrophil elastase (NE). Finally, the signalling pathway by which atorvastatin exerts its effect on MMP release was investigated in both culture systems.
MATERIALS AND METHODS
Materials
Native type I collagen (rat tail tendon collagen) was extracted from rat tail tendons by a previously published method 16, 17. Commercially available reagents were obtained as follows: Dulbecco's modified Eagle's medium (DMEM) and fetal calf serum (FCS) were from Invitrogen Life Technologies (Grand Island, NY, USA); human NE was from Elastin Products Company, Inc. (Owensville, MO, USA); recombinant human MMP-1 and -3 Western blotting standards, recombinant human tumour necrosis factor (TNF)-α and interleukin (IL)-1α and monoclonal anti-human pro/active MMP-1 (Clone 36665) and MMP-3 (Clone 10D6) antibodies were from R&D Systems, Inc. (Minneapolis, MN, USA); horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG was from Rockland (Gilbertsville, PA, USA); fluvastatin and simvastatin were from Calbiochem (San Diego, CA, USA); mevalonate, farnesyl pyrophosphate (FPP), geranylgeranyl pyrophosphate (GGPP), chloramine-T, p-dimethylaminobenzaldehyde and 3-(4,5-dimethylthiazolyl-2)-2,5- diphenyltetrazoliumbromide (MTT) were from Sigma (St. Louis, MO, USA).
Atorvastatin was a kind gift from Pfizer Inc. (New York, NY, USA) and was dissolved in methanol to a stock solution of 2.5 mM.
Cell culture
HFL-1 (lung, diploid, human) was obtained from the American Type Culture Collection (ATCC; #CCL-153; Rockville, MD, USA). HT1080 cells (connective tissue fibrosarcoma, diploid, human) were also purchased from ATCC (#CCL-121). HLFs were isolated from alveolar lung tissue removed at surgery for suspected malignancy that appeared to be free of cancer under a protocol approved by the Human Studies Committee of the University of Michigan (Ann Arbor, MI, USA). The cells were cultured on 100-mm tissue culture dishes (Falcon; Becton-Dickinson Labware, Lincoln Park, NJ, USA) with DMEM supplemented with 10% FCS, 100 μg·mL−1 penicillin, 250 μg·mL−1 streptomycin sulfate (penicillin-streptomycin; Invitrogen), and 2.5 μg·mL−1 amphotericin B (Geneva Pharmaceuticals, Inc., Dayton, NJ, USA). Cells were cultured at 37°C in a humidified atmosphere of 5% CO2 and passaged every 3 to 5 days at a 1:4 ratio. In all experiments, HFL-1 cells between the 14th and 18th passage were used. For HLF, passages between the seventh and 11th were used. HT1080 cell culture media were used as a positive control for gelatine zymography.
To evaluate the release of MMP in monolayer culture, fibroblasts were cultured in DMEM with 10% FCS in 60-mm dishes (Falcon) at 0.5×106 cells for 24 h. The medium was then changed to serum-free (SF)-DMEM. After 24 h, cells were washed with phosphate-buffered saline (PBS) once and treated with statins for 1 h before stimulation with cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) in SF-DMEM. After 24-h incubation, the media were harvested for gelatine zymography and Western blot analysis.
Collagen gel culture
Collagen gel cultures were conducted according to the previously published methods 17 with a cell density of 3×105 fibroblasts·mL−1 gel. The gels were incubated in SF-DMEM either with or without designated reagents at 37°C and 5% CO2 for 3 days.
Hydroxyproline assay
Hydroxyproline, which is directly proportional to type I collagen content, was measured by spectrophotometry according to the previously published methods 18. Briefly, the collagen gels were solubilised by heating in 0.1M HCl (50 μL/3 gels). Samples (20 μL) were mixed with 30 μL 3.3M NaOH and autoclaved at 120°C for 20 min. After oxidisation with 0.056M chloramine-T for 20 min, samples were reacted with Ehrlich's reagent at 65°C for 20 min. The absorbance was measured at 540 nm with BenchMark Microplate Reader and Microplate Manager III software (Bio-Rad, Hercules, CA, USA).
Western blot analysis of metalloproteinase
To assess the release of MMP-1 and -3, Western blot analysis was performed according to the previously published method 19, 20. Briefly, the supernatants (4 mL per condition) from monolayer and 3D collagen gel cultures were precipitated with 50% (vol/vol) cold ethanol and re-suspended in 50 μL double-distilled H2O. Because secreted MMPs were assessed, no internal standard to allow for variable dilution was used. Rather, a constant volume, 35 μL of each sample was subjected to 10% sodium dodecyl sulfate (SDS)–polyacrylamide gel elecrophoresis (PAGE) under nonreducing conditions and then proteins were transferred to polyvinyl difluoride membranes (Bio-Rad). This then allowed comparison of the intensity of the final bands in a manner similar to immunoassay 21, 22. After blocking with 5% nonfat milk, blots were incubated overnight at 4°C with primary antibodies (1 μg·mL−1 mouse anti-human MMP-1 or MMP-3 monoclonal antibody). Target proteins were detected using HRP-conjugated goat anti-mouse immunogloblin G in conjunction with an enhanced chemiluminescence detection system (Amersham Biosciences UK Limited, Little Chalfont, UK) using a Typhoon Scanner (Amersham Biosciences). Band intensity was quantified using ImageJ software version 1.34s (National Institute of Health; NIH, USA).
Gelatine zymography
Supernatant media from monolayer and 3D collagen gel cultures (500 μL per culture condition) were concentrated 10-fold by precipitation with cold ethanol and re-suspended in 50 μL double-distilled H2O. Gelatine zymography was performed by a modification of previously published procedures 20, 23, 24. Briefly, 25 μL of each sample were separated in 10% SDS–PAGE containing 0.1% gelatine (Invitrogen, Carlsbad, CA, USA) under nonreducing conditions. After electrophoresis, the gels were soaked in 2.5% Triton-X 100 and incubated in metalloproteinase activation buffer containing CaCl2 and ZnCl2 for 16 h at 37°C. The gels were stained with 0.4% Coomassie blue (Bio-Rad) and rapidly destained with destaining buffer (30% methanol, 10% acetic acid). The gels were dried between cellophane sheets (Amersham, Piscataway, NJ, USA). Zones of proteolysis appeared as clear bands against a blue background and were scanned by Gel Doc 2000 (Bio-Rad). Band intensity was quantified using ImageJ software.
RNA preparation and complementary DNA synthesis
To determine whether changes in MMP mRNA levels were present, quantitative real-time PCR was performed. HFL-1 fibroblasts were cultured in 60-mm dishes at 1.5×106 cells for 24 h. Cells were then washed by PBS twice and treated with statins for 1 h before the stimulation by cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) in SF-DMEM for 6 h. Total RNA was isolated by a single-step guanidinium–thiocyanate–phenol–chloroform extraction procedure designed by Chomcynski and Sacchi 25, and the total amount of RNA was quantified spectrophotometrically (Shimadzu Scientific Instruments, Inc., Columbia, MD, USA). 1 µg of total RNA was transcribed according to the previously published methods 20, 24.
Quantitative real-time PCR
Gene expression was measured with the use of an ABI Prism 7500 Sequence Detection System (Applied Biosystems) by a slight modification of previously published methods 20, 26. The same primers and probes for MMP-1, -3 and -9 were used as described by Fang et al. 20. rRNA was simultaneously tested using TaqMan Ribosomal RNA Control Reagents (Applied Biosystems).
Cell viability assay
Cell viability was evaluated by MTT assay using previously described methods 27. Cells were treated with various concentrations of reagents used in this study with MTT solution at a final concentration of 1 mg·mL−1 for 6 h at 37°C. After incubation, dimethyl sulfoxide was added into each well. The absorbance of each sample at 540 nm was determined by BenchMark Microplate Reader (Bio-Rad).
Statistical analysis
Data are expressed as mean±sem. Each experiment was repeated at least three times. The number of replicates is specified in each figure. Experiments with multiple comparisons were evaluated by one-way ANOVA followed by post hoc analysis by pairwise comparisons of values with Tukey's test to adjust for multiple comparisons. Probability p-values <0.05 were considered significant.
RESULTS
Effect of statins on MMP release in monolayer culture
HFL-1 cells
To test if statins inhibit the release of MMPs in HFL-1, we first investigated three separate statins, not only to demonstrate the reproducibility of our findings, but to be certain that any effects observed were not specific for a single compound. To accomplish this, cells were cultured in 60-mm dishes and treated with atorvastatin (2.5 μM), fluvastatin (1.0 μM) or simvastatin (0.5 μM) with and without stimulation by cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) for 24 h. These concentrations of statins were chosen as they are considered to be close to those detected in the human body when administered therapeutically. Supernatants from each culture condition were assessed for MMP-1 and -3 release by Western blot analysis, and for MMP-2 and -9 by gelatine zymography. As expected, the cytokines stimulated MMP-1 and -3 release (fig. 1a–d⇓). Bands corresponding to the latent forms (55 kDa) were observed, but lower molecular weight bands corresponding to active MMPs were not observed. All three statins inhibited both MMP-1 (fig. 1a and b⇓) and MMP-3 (fig. 1c and d⇓) release stimulated by cytokines (p<0.05 compared with the cytokine-treated controls). In monolayer culture, MMP-9 release was not observed under any condition. However, HFL-1 cells released a gelatinase corresponding to the latent form of MMP-2 (fig. 1e and f⇓). The effect of atorvastatin on the release of the latent form of MMP-2 stimulated by cytokines was modest (fig. 1e and f⇓). None of the reagents affected cell viability evaluated by MTT assay (data not shown).

Effect of statins on matrix metalloproteinase (MMP) release by fibroblasts in monolayer culture. Human fetal lung fibroblast-1 cells cultured in monolayer were treated with atorvastatin (Atr; 2.5 μM), fluvastatin (Flu; 1.0 μM) and simvastatin (Sim; 0.5 μM). 1 h later, cytokines (CK; tumour necrosis factor-α 5 ng·mL−1 and interleukin-1α 2 ng·mL−1) were added. Supernatant media harvested after 24 h were subjected to Western blot analysis for MMP-1 and MMP-3, and gelatine zymography for MMP-2 and MMP-9. a) MMP-1 western blot. The band corresponding to the latent form is indicated by the arrow. b) Quantification of MMP-1 staining. c) MMP-3 western blot. The band corresponding to the latent form of MMP-3 is indicated by the arrow. d) Quantification of MMP-3 staining. e) Gelatine zymography for MMP-2 and MMP-9. f) Quantification of MMP-2 staining. Densitometry data are shown as mean±sem from three separate experiments performed on different occasions. STD: MMP-1 and MMP-3 standards; CTR: control; HT1080: supernatant from HT1080 cell monolayer culture as a positive control. *: p<0.05 compared with the cytokine-treated controls.
To determine whether the inhibition of MMP-1 and -3 expression by atorvastatin was associated with changes in gene expression, quantitative real-time PCR was performed after 6 h of incubation with statins and cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1). Atorvastatin alone slightly stimulated both MMP-1 (fig. 2a⇓) and MMP-3 (fig. 2b⇓) mRNA expression. However, this effect was modest compared with that of the cytokines, which stimulated MMP-1 and -3 mRNA 121.6±3.8- and 46.9±1.0-fold of control, respectively. When cells were treated with atorvastatin before the stimulation by cytokines, the effect of the cytokines on both MMP-1 and -3 mRNA was inhibited to 12.3±1.4- and 7.1±0.4-fold of control, respectively (p<0.01 compared with the cytokine treated groups). Consistent with the results observed with gelatine zymography, no MMP-9 mRNA was detected (data not shown).
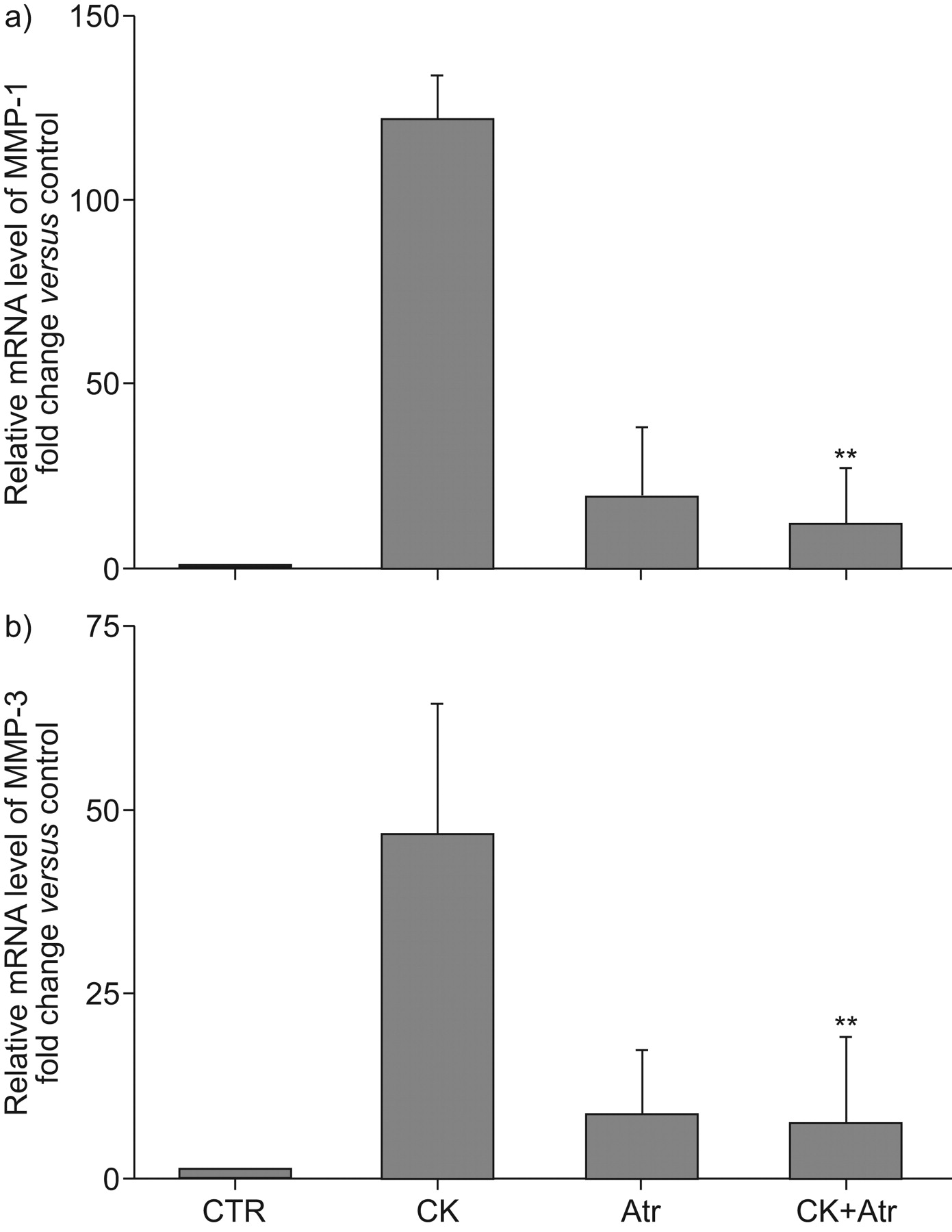
Effect of atorvastatin (Atr) on a) matrix metalloproteinase (MMP)-1 and b) MMP-3 mRNA expression. Human fetal lung fibroblast-1 fibroblasts cultured in monolayer were treated with and without atorvastatin (2.5 μM). 1 h later, cytokines (CK; tumour necrosis factor-α 5 ng·mL−1 and interleukin-1α 2 ng·mL−1) were added and then cultured for 6 h. mRNA levels were quantified by Taqman RT-PCR. The vertical axes show mRNA expression level normalised to the amount of rRNA and expressed as fold of control. The data presented are mean±sem from three separate experiments, each performed in duplicate. CTR: control. **: p<0.01 compared with cytokine treated groups.
Adult lung fibroblasts
Because fibroblasts may be functionally heterogeneous, we also investigated the effect of atorvastatin on the inhibition of MMP release from normal adult human lung fibroblasts (HLF) after 24 h of culture. In contrast to HFL-1 cells, which did not release any detectable MMP-1 or -3 under control conditions, HLF cells released a small amount of MMP-1 and -3 corresponding in molecular size to the latent form of 55 kDa (online supplementary material fig. 1⇑). As with HFL-1 cells, the cytokines also stimulated both MMP-1 and -3 release from HLF cells (online supplementary material fig. 1a–d⇑). Also, as with HFL-1 cells, atorvastatin inhibited HLF release of both MMP-1 (online supplementary material fig. 1a and b⇑) and MMP-3 (online supplementary material fig. 1c and d⇑) release stimulated by cytokines (p<0.05 compared with the cytokine-treated controls). The release of MMP-2 was similar to that of HFL-1 cells and, as with HFL-1 cells, MMP-9 was not detected in the supernatant of HLF cells in monolayer cultures (online supplementary material fig. 1e and f⇑).
Role of the mevalonate pathway
Inhibition of MMP-1 and MMP-3 protein release
Incubation of cells with HMG-CoA reductase inhibitors, such as the statins, blocks production of mevalonate 28, 29. Mevalonate metabolism yields a series of isoprenoids, including FPP and GGPP. To confirm that this is the pathway by which atorvastatin inhibits MMP-1 and -3 release, we tested whether mevalonate, FPP or GGPP could reverse atorvastatin inhibition of cytokine-stimulated HFL-1 MMP-1 and -3 release. HFL-1 cells were cultured in monolayer and treated with atorvastatin (2.5 µM) with or without mevalonate (100 µM), FPP (5 µM) or GGPP (5 µM) followed by stimulation with cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) for 24 h. In the presence of mevalonate, GGPP or FPP alone, there was no detectable level of MMP-1 release (fig. 3a⇓) and this corresponded to the results under control conditions (data not shown). The addition of mevalonate and GGPP almost completely blocked the inhibitory effect of atorvastatin on the release of MMP-1 (fig. 3a and b⇓; p<0.05 compared with cytokine-treated groups after incubation with atorvastatin). In contrast, FPP had no effect.

Effect of mevalonate (Mev) and isoprenoids on atorvastatin inhibition of a) MMP-1 and b) MMP-3 release. Human fetal lung fibroblast-1 cells were cultured in monolayer and treated with atorvastatin (2.5 µM) with or without Mev, geranylgeranyl pyrophosphate (GGPP) or farnesyl pyrophosphate (FPP). 1 h later, cytokines (CK; tumour necrosis factor-α 5 ng·mL−1 and interleukin-1α 2 ng·mL−1) were added. Supernatant media harvested after 24 h were subjected to Western blot analysis for MMP-1 and MMP-3. a) MMP-1 and c) MMP-3 Western blots. The band corresponding to the latent form is indicated by the arrow. Quantification of b) MMP-1 and d) MMP-3 staining. Densitometry data are shown as mean±sem for three separate experiments performed on separate occasions. *: p<0.05.
No MMP-3 was detected when cells were incubated under control conditions (data not shown) or with mevalonate, GGPP or FPP added alone (fig. 3c⇑). Mevalonate and GGPP blocked the inhibitory effect of atorvastatin on MMP-3 release that was stimulated by cytokines (fig. 3c and d⇑; p<0.05 compared with cytokine-treated groups after incubation with atorvastatin). FPP was without effect.
Inhibition of MMP-1 and MMP-3 mRNA expression
To determine whether the effects of mevalonate and GGPP were associated with changes in gene expression, quantitative real-time PCR assay was performed. HFL-1 cells were treated with atorvastatin (2.5 µM) with or without mevalonate (100 µM) or GGPP (5 µM), and then stimulated with cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) for 6 h. Neither mevalonate nor GGPP alone altered MMP-1 or -3 mRNA expression levels (fig. 4a and b⇓, respectively; p<0.01 compared with cytokines plus atorvastatin treated groups). However, the addition of mevalonate or GGPP blocked the inhibitory effect of atorvastatin on MMP-1 and -3 mRNA expression significantly (fig. 4a and b⇓, respectively). FPP did not have any effect on MMP-1 or -3 mRNA expression (data not shown).

Effect of mevalonate (Mev) and isoprenoids on atorvastatin inhibition of a) matrix metalloproteinase (MMP)-1 and b) MMP-3 mRNA expression. Human fetal lung fibroblast-1 cells were cultured in monolayer and treated with atorvastatin (2.5 µM) with or without Mev or geranylgeranyl pyrophosphate (GGPP). 1 h later, cytokines (CK; tumour necrosis factor-α 5 ng·mL−1 and interleukin-1α 2 ng·mL−1) were added and cultured for 6 h. mRNA levels were quantified by Taqman RT-PCR. Vertical axes show the mRNA expression level normalised to the amount of rRNA and expressed as fold of control. The data presented are mean±sem from three separate experiments, each performed in duplicate. **: p<0.01.
Culture in 3D collagen gels
Effect of atorvastatin on collagen degradation
The response of fibroblasts to exogenously added mediators can vary depending on the culture condition. In 3D cultures in floating collagen gels, which are thought to more closely mimic in vivo conditions, we have previously shown that stimulation of fibroblasts with cytokines in the presence of NE can result in degradation of the collagen gel matrix 19. To determine if atorvastatin inhibition of fibroblast MMP release could lead to inhibition of matrix degradation, we prepared 3D collagen gels populated with HFL-1 fibroblasts. When stimulated by cytokines (TNF-α 5 ng·mL−1 and IL-1β 2 ng·mL−1) to induce MMP release in the presence of NE (10 nM), which leads to activation of MMPs, the collagen gels were degraded as we have reported previously 19. The hydroxyproline content of the gels was significantly reduced after 3 days (hydroxyproline content 38±4% of control (fig. 5⇓); p<0.05 compared with control). This decrease was almost completely inhibited by atorvastatin (hydroxyproline content 83±8% of control (fig. 5⇓); p<0.05 compared with cytokine + NE). Hydroxyproline content of gels treated with atorvastatin + cytokines (without NE) or atorvastatin + NE (without cytokines) was the same as that of control (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of atorvastatin on collagen gel degradation induced by cytokines (CK) together with neutrophil elastase (NE). Human fetal lung-1 fibroblasts (3×105 cells·mL−1) were cast into three-dimensional collagen gels and released into floating media (serum-free Dulbecco’s modified Eagle’s medium) containing atorvastatin (2.5 μM). 1 h later, CK (tumour necrosis factor-α 5 ng·mL−1 and interleukin-1α 2 ng·mL−1) were added; NE was added the next day. The gels were cultured for three more days and hydroxyproline content in the gels was determined. The data presented are mean±sem from three separate experiments, each of which included triplicate gels for each condition. *: p<0.05.
Effect of atorvastatin on MMP release
To confirm the results obtained in monolayer culture on plastic, we also assessed the effect of atorvastatin on MMP release in 3D culture in collagen gels. There were modest differences in MMP release under control conditions. For example, detectable MMP-1 was released in latent form under control conditions in 3D culture, which contrasts with the undetectable levels in monolayer culture. Importantly, as with monolayer culture, there was marked increase in release of MMP-1 following cytokine stimulation (online supplementary material fig. 2⇑). As in monolayer culture, MMP-3 was undetectable under control conditions in 3D culture (online supplementary material fig. 3⇑). MMP-2, which was released in the latent form in monolayer culture, was released in both latent and active forms under control conditions in 3D culture (online supplementary material fig. 4⇑). MMP-9, which was not released under any conditions tested in monolayer culture, was not released under control conditions in 3D culture, but its release was stimulated by cytokines (TNF-α 5 ng·mL−1 and IL-1α 2 ng·mL−1) (online supplementary material fig. 4⇑). Importantly, the cytokines stimulated the release of all the MMPs assessed, MMP-1, -3 and -9, and this release was inhibited (p<0.05 compared with cytokines) in all cases by atorvastatin (online supplementary material figs 2⇑, 3⇑ and 4⇑). There was, moreover, a reduction in the conversion of latent MMPs to the active forms in the presence of NE for MMP-1 (p<0.05), -3 (p<0.01) and -9 (p<0.001). A modest effect, which was not quantified, was also observed in inhibiting MMP-2 release (online supplementary material fig. 4a⇑).
Role of the mevalonate pathway in inhibition of collagen gel degradation
To confirm that atorvastatin inhibited collagen gel degradation by the same pathway by which it inhibited MMP release in monolayer culture, we assessed the ability of mevalonate (100 μM), FPP (5 μM) and GGPP (5 μM) to block atorvastatin inhibition of collagen gel degradation induced by the combination of cytokines and NE. After 3 days of culture in the presence of cytokines and NE, collagen gel hydroxyproline content was reduced by more than half (online supplementary material fig. 5⇑; p<0.05 compared with control), indicating degradation of the collagen gels. Atorvastatin almost completely inhibited this degradation (p<0.05 compared with cytokines + NE). Mevalonate and GGPP had no effect when added alone, but almost completely reversed atorvastatin inhibition of collagen degradation caused by cytokines and NE (fig. 3⇑; p<0.05 compared with control). In contrast, FPP had no effect when added alone and did not reverse the inhibitory effect of atorvastatin on collagen gel degradation.
Role of the mevalonate pathway in 3D culture
Because fibroblasts in 3D culture release MMP somewhat differently than in monolayer culture, we also investigated whether mevalonate or isoprenoids blocked the effect of atorvastatin on MMP release in 3D gel culture (online supplementary material fig. 6). Although statistical significance was not achieved, there was a trend for both mevalonate and GGPP to block the inhibitory effect of atorvastatin on the amount of detectable active MMP-1 released in the presence of cytokines and NE (online supplementary material fig. 6a and b). Both mevalonate and GGPP significantly blocked the inhibitory effect of atorvastatin on MMP-3 release (online supplementary material fig. 6c and d; p<0.01) and on the release of MMP-9 and its conversion to an 84 kDa size that corresponds to the active form MMP-9 (online supplementary material fig. 6e and f; p<0.01). FPP was consistently without effect.
Adult HLF in 3D culture
Collagen degradation
To confirm that collagen gel degradation occurs in the gels populated with adult HLF and that MMP release by these cells is also blocked by atorvastatin, cultured HLF cells were cast into 3D cultures and treated with cytokines and NE to induce degradation. As observed with HFL-1 cells, in the presence of cytokines and NE, hydroxyproline content of the gels was significantly reduced after 3 days of culture (online supplementary material fig. 7; p<0.05 compared with control). This decrease was inhibited by atorvastatin (p<0.05 compared with cytokines + NE). The role of the mevalonate pathway also appeared to be the same in HLF cells, as mevalonate and GGPP had no effect when added alone, but reversed the effect of atorvastatin on the inhibition of collagen degradation caused by cytokines and NE (online supplementary material fig. 7; p<0.05 compared with control).
DISCUSSION
The current study demonstrates that HMG-CoA reductase inhibitors can inhibit cytokine-induced release of MMPs from human lung fibroblasts cultured in both monolayer and 3D collagen gels. Release of both MMP-1 and -3 from fibroblasts in monolayer culture was inhibited by atorvastatin, simvastatin and fluvastatin. Atorvastatin also inhibited the cytokine-induced expression of MMP-1 and MMP-3 mRNA expression. In 3D gel culture, atorvastatin inhibited the degradation of collagen gels that was induced by cytokines in the presence of NE. In contrast to monolayer culture, in 3D gel culture, fibroblasts were induced by cytokines to release MMP-9, and these were also inhibited by atorvastatin. All of these changes were reversed by the addition of mevalonate and GGPP, which are intermediates in the HMG-CoA reductase pathway that bypass the step inhibited by statins. Interestingly, the intermediate FPP did not reverse these effects of atorvastatin. Taken together, the current data suggest that statins may modulate remodelling processes mediated by lung fibroblasts by inhibiting the release of MMPs.
Since the classic studies of Laurell and Eriksson 30, proteases have been believed to play a major role in the pathogenesis of COPD. In particular, the MMPs have been suggested to play a prominent role. In support of this concept, mice deficient in the macrophage elastase MMP-12 have been reported to be resistant to the development of cigarette smoke-induced emphysema 31, although MMP-12 is not prominent in human emphysema 32. Increased MMP-9 expression in the lungs has been reported to be associated with emphysema, in both experimental 33–37 and clinical 38 studies. Moreover, immunohistochemical analysis of COPD lungs showed an increased expression of collagenase 1 (MMP-1) and collagenase 2 (MMP-8) 39, and alveolar epithelial cells from patients with emphysema express MMP-1 mRNA and protein, and exhibit collagenase activity 40.
MMPs are expressed in low to undetectable levels in normal lung, but are readily detected in many lung diseases in addition to emphysema. In this context, the expression of MMPs is not restricted to inflammatory cells; virtually all cells, including alveolar epithelial cells and fibroblasts, can make MMPs 40, 41. Alveolar fibroblasts, which comprise ∼40% of alveolar cells 42, are believed to be the major cells responsible for the production and remodelling of ECM. They are one of the major sources of MMPs, which are secreted following lung injury. Therefore, inhibition of MMP release from fibroblasts could confer a new therapeutic approach to prevent the lung destruction observed in emphysema. Whether inhibition by statins could lead to toxicities, e.g. by inhibiting lung tissue turnover in fibrotic areas, of course, is a potential toxicity that would need to be considered.
Statins were originally developed for their cholesterol-lowering properties and efficacy in cardiovascular disease. However, evidence of pleiotropic effects of statins has been accumulating and suggests immunomodulatory, antioxidant, antithrombogenic and vascular actions 7, 29. Several statins have been reported to inhibit the expression of MMPs in vascular cells and macrophages in vitro 9–11. Consistent with this effect on MMPs, statins have been reported to inhibit the development of emphysema in model systems. In a rat model of cigarette smoking-induced emphysema, Lee et al. 43 found that simvastatin inhibited lung parenchymal destruction and MMP-9 expression. In a mouse model of emphysema induced by elastase, simvastatin reduced mRNA expression for interferon-γ, TNF-α and MMP-12 in the whole lung 44.
The current study extends these observations by providing a basis for statins to alter lung structural remodelling by a direct effect on fibroblast MMP release. Further, our study demonstrates that the effect of the statins is mediated through the mevalonate pathway, which is likely to involve GGPP but not FPP. This observation is similar to other studies, which also observed that the addition of FPP did not reverse the effect of statins 45, 46. The mechanism for the failure of FPP to reverse these statin effects remains unexplained.
As with the normal lung, fibroblasts in culture release very low amounts of MMP-1, -2, -3 and -9. The release of these MMPs, however, can be readily induced by the inflammatory cytokines TNF-α + IL-1β, and it is the induced MMP release that statins were demonstrated to inhibit in the current study. This inhibition occurred both in the more conventionally used monolayer culture system (on plastic dishes) and in culture in 3D collagen gels, a system that is believed to more closely resemble tissue conditions 14, 16, 47. In this system, degradation of the ECM can be quantified, and statins also inhibited this process. Some modest differences were observed between cells grown in monolayer and 3D culture. Part of these differences could be related to MMP retention in the collagen gel in the 3D system. This is unlikely, however, to account for the production of MMP-9 in 3D culture, which was not observed in monolayer culture. While there were some differences between the culture systems, the consistency of the inhibitory effects of statins, which were also observed in both the fetal strain of fibroblasts HFL-1 that has been widely used to explore lung fibroblast biology and in a strain of normal adult fibroblasts, suggests that the inhibitory effects of statins are robust.
In the current study, statins slightly increased the release of MMP-1 when added alone, and this was confirmed at both the mRNA and protein levels. Atorvastatin also slightly increased MMP-3 mRNA when added alone; however, MMP-3 was not detected at the protein level. The mechanisms that lead to mild stimulation when added alone and marked inhibition when added together with cytokines are unclear, but similar effects have been observed in other regulatory networks when agents can have multiple effects. These observations, therefore, suggest that statins might be modulating MMP release at several levels.
In summary, the current study demonstrates that statins can inhibit MMP release from HLFs in both monolayer and 3D collagen gel cultures. This effect is mediated through the inhibition of the HMG-CoA reductase pathway. Through actions on fibroblast MMP release, statins may modulate remodelling processes mediated by fibroblasts. The use of statins, therefore, may have therapeutic potential in diseases characterised by alterations in tissue architecture, particularly diseases characterised by tissue destruction, such as emphysema.
Support statement
Funded by a grant from Pfizer Pharmaceaticals.
Statement of interest
A statement of interest for S.I. Rennard, and this study itself can be found at www.erj.ersjournals.com/misc/statements.dtl
Acknowledgments
The authors thank the excellent secretarial support of L. Richards.
Footnotes
-
This article has supplementary material accessible from www.erj.ersjournals.com
- Received October 12, 2007.
- Accepted September 4, 2009.
- © ERS Journals Ltd
References










