





Abstract
The aim of this study was to investigate the presence of epithelial neutrophil-activating peptide (ENA)-78 in pleural effusions, as well as the chemoattractant activity of pleural ENA-78 on neutrophils.
Pleural effusion and serum samples were collected from 75 patients who presented to the respiratory institute (19 with malignant pleural effusion, 21 with tuberculous pleural effusion, 18 with infectious pleural effusion and 17 with transudative pleural effusion). The concentrations of ENA-78, myeloperoxidase and neutrophil elastase were determined, and the chemoattractant activity of ENA-78 for neutrophils both in vitro and in vivo was also observed.
The concentrations of ENA-78, myeloperoxidase and neutrophil elastase in infectious pleural effusion were significantly higher than those in malignant, tuberculous and transudative groups, respectively (all p<0.01). Infectious pleural fluid was chemotactic for neutrophils in vitro and anti-ENA-78 antibody could partly inhibit these chemotactic effects. Intrapleural administration of ENA-78 produced a marked progressive influx of neutrophils into pleural space.
Compared with noninfectious pleural effusion, ENA-78 appeared to be increased in infectious pleural effusion. Our data suggested that ENA-78 was able to induce neutrophil infiltration into pleural space and might be responsible for pleural neutrophil degranulation.
The inflammatory process results in increased pleural vascular permeability, leading to the accumulation of fluid enriched in proteins and to the recruitment of cells with the development of exudative pleural effusion (PE) 1. PE is characterised by the presence of specific subsets of leukocytes 1, 2 which, together with pleural mesothelial cells, contribute to the local production of cytokines and chemokines 3, 4. Analysis of malignant and tuberculous PEs usually shows a lymphocytic preponderance; whereas infectious PEs, including empyema and parapneumonic effusion, are typically associated with an influx of neutrophils 5, 6.
Neutrophils are recruited to sites of inflammation by a variety of chemoattractants, including complement factors (C5a), interleukin (IL)-8, arachidonic acid metabolites (leukotriene B4), and infectious peptides (N-formyl-methionyl-leucyl-phenylalanine), which are generated and released locally at sites of injured tissue. Neutrophils release reactive oxygen and nitrogen species into the extracellular milieu, in addition to releasing their granular content, which contains several serine and neutral proteases that can produce tissue injury 7. Chemokines are potent proinflammatory molecules, and their regulation must therefore be tightly controlled 8. One of these, a CXC chemokine, is epithelial neutrophil-activating peptide (ENA)-78 (also named CXCL5), which contains a Glu-Leu-Arg motif essential for neutrophil-stimulating activity 9, 10. ENA-78 has been implicated in the pathogenesis of a number of pulmonary diseases 11–13. The present study was performed to determine total and differential leukocyte counts and the concentrations of ENA-78 and two indicators of neutrophil degranulation (myeloperoxidase (MPO) and neutrophil elastase (NE)) in pleural fluid and serum of patients with PEs of various aetiologies, in order to: 1) determine whether PE ENA-78 is produced in the pleural space; 2) establish the relationship between this chemokine to the neutrophil number and activation state in PE; and 3) explore the chemoattractant activity of ENA-78 on neutrophils.
MATERIALS AND METHODS
Subjects
A prospective design study was performed in our institute of respiratory diseases (First Affiliated Hospital, Guangxi Medical University, Guangxi, China) from October, 2006 to March, 2008. The study protocol was approved by our institutional review board for human studies and informed consent was obtained from all subjects. 75 consecutive patients with PE of various causes were recruited in the present study.
Malignant PE was collected from 19 patients (aged 34–74 yrs) with newly diagnosed lung cancer with PE. Histologically, 14 cases were adenocarcinoma and 5 were squamous cell carcinoma. A diagnosis of malignant PE was established by demonstration of malignant cells in PE and/or on closed pleural biopsy specimen.
21 patients (aged 17–68 yrs) were proven to have tuberculous PE, as evidenced by: a compatible clinical history associated with presence of acid-fast bacilli in PE specimen or by demonstration of granulomatous pleurisy on closed pleural biopsy specimen in the absence of any evidence of other granulomatous diseases; an exudative lymphocytic effusion with an adenosine deaminase level >40 U·L−1, along with a positive purified protein derivative skin test result and the exclusion of any other potential causes of pleurisy; or following antituberculosis chemotherapy, the resolution of PE and clinical symptoms was observed.
18 PE patients (aged 16–57 yrs) were classified as infectious PE (including 16 empyema and two parapneumonic effusion). Empyema was defined as an effusion that met one or more of the following criteria: purulent fluid on macroscopic examination, positive Gram stain and/or growth of organisms in culture, and PE with pH<7.2 or glucose <3.3 mmol·L−1 in association with pneumonia. Parapneumonic effusion was defined as those with a glucose concentration >3.3 mmol·L−1 and pH >7.2, and no organisms seen on Gram stain or found on PE culture in a patient with pneumonia.
17 patients (aged 17–70 yrs) with PE were classified as transudates on the basis of Light's criteria 14.
Patients were excluded if they had received any invasive procedures directed into the pleural cavity or if they had suffered chest trauma within 3 months prior to hospitalisation or had a PE of undiagnosed cause. At the time of sample collection, none of the patients had received any antituberculosis therapy, anticancer treatment, corticosteroids or other nonsteroidal anti-inflammatory drugs.
Sample collection and processing
PE samples were collected from each subject in heparin-treated tubes, using a standard thoracocentesis technique within 24 h after hospitalisation. 10 mL of venous blood was drawn simultaneously for obtaining sera. The PE specimens were immersed in ice immediately and were then centrifuged at 1,200×g for 5 min. The cell-free supernatants of PE and serum were frozen at -80°C immediately after centrifugation, for later determining concentrations of ENA-78, MPO and NE. Total and differential cell counts, and detections of concentrations of pleural protein, glucose, lactate dehydrogenase and adenosine deaminase, were performed in addition to cytological and microbiological examination of pleural fluid. A pleural biopsy was performed when the results of pleural fluid analysis were suggestive of tuberculosis or malignancy.
Measurement of ENA-78, MPO and NE
The concentrations of ENA-78, MPO and NE in both PEs and sera were measured by sandwich ELISA kits according to the manufacturers’ protocols. The ELISA kits for ENA-78 and MPO were purchased from R&D Systems Inc. (Minneapolis, MN, USA), and the ELISA kits for NE were purchased from Cell Sciences® (Canton, MA, USA). All samples were assayed in duplicate. The lower detection limits were: 15 ng·L−1 (ENA-78); 0.1 μg·L−1 (MPO); and 0.4 μg·L−1 (NE).
Neutrophil chemotaxis assay
Human neutrophils were prepared from peripheral blood by Ficoll–Hypaque density gradient in 0.9% saline (Sigma Chemical Co., St. Louis, MO, USA). Neutrophils were separated from erythrocytes by hypotonic lysis and then suspended in Hanks' balanced salt solution (HBSS) with calcium/magnesium (Gibco; Grand Island, NY, USA) at 2.0×106 cells·mL−1 with >95% viability by trypan blue exclusion. 200 μL of the neutrophil suspension (1.0×106 cells·mL−1 in RPMI-1640 media) were loaded into the upper well of a 24-well Transwell chamber (Costar, Corning NY, USA) while 30 µL pleural fluid from five patients with malignant PE, five with tuberculous PE, five with empyema or five with transudates, was placed in the bottom chamber. The two wells were separated by a polycarbonate filter paper with a pore size of 5 µm and the chamber was incubated at 37°C for 45 min. At the end of incubation, the filter was fixed, stained and mounted on a glass microscope slide. Migration was assessed by counting the number of cells that had migrated beyond a certain depth into the filter (50 µm). To correct for interdonor variation, migration data of test samples were compared with their corresponding control values (HBSS alone) and expressed as a percentage above the control value. To demonstrate that ENA-78 was responsible for neutrophil migration, blocking experiments were performed by mixing the PE with 10 µg·mL−1 of anti-ENA-78 monoclonal antibody (mAb) or immunoglobulin G2a irrelevant isotype control (R&D System Inc.).
Effects of intrapleural injected ENA-78 on neutrophil recruitment
After an additional study protocol was approved by our institutional review board and informed consent was obtained from the subjects studied, a total of 10 patients with tuberculous PE were included in this section of the study. Immediately after collection of PE samples, 10 μg of recombinant human ENA-78 (R&D Systems Inc.) in vehicle (0.1% human serum albumin in 0.9% saline) was injected into the pleural space of five patients and vehicle only was injected into the pleural space of the other five patients. Intrapleural injection of ENA-78 or vehicle was randomised. The dose of ENA-78 was based upon a preliminary study involving two PE patients. PE collection for determining neutrophil number was repeated 6, 12 and 24 h after the injection of ENA-78 or vehicle.
Statistical analysis
Data were expressed as mean±sem or median (25th–75th percentile). Comparisons of the data between different groups were performed using Kruskal–Wallis one-way ANOVA on ranks. For the levels of ENA-78, MPO and NE in PE and in corresponding serum, paired data comparisons were made using a Wilcoxon signed-rank test. The correlations between variables were determined by Spearman’s rank correlation coefficients. The effects of intrapleural injected ENA-78 or vehicle on neutrophil recruitment were compared through one-way repeated-measures ANOVA. Analysis was completed with SPSS version 14.0 Statistical Software (Chicago, IL, USA), and a p-value <0.05 was considered significant.
RESULTS
Characteristics in PEs
Cytological characteristics in PEs are illustrated in table 1⇓. Subjects with lung cancer showed a large proportion of lymphocytes and macrophages in PE. Importantly, on cytological examination, malignant cells were found in 13 subjects. Subjects with tuberculosis showed a marked elevation of total cell counts and a large proportion of these cells were lymphocytes, with some neutrophils and macrophages. Absolute lymphocyte counts evidenced the highest values in tuberculous PE, showing a significant increase in comparison with each of the PEs induced by the other causes (all p<0.05). Also, as expected, total cell counts in infectious PE were the greatest among the four groups (all p<0.05) and neutrophils were the predominant cell type; significantly greater than those in the PEs with any other causes (all p<0.01).
Cytological characteristics in pleural effusions(PE)
Concentrations of ENA-78, MPO and NE in PEs
As shown in table 2⇓ and figure 1⇓, the concentration of ENA-78 in infectious PE (median 2,639.2 (25th–75th percentile, 448.8–5,014.6) ng·L−1) was significantly higher than those in malignant PE (88.0 (52.2–337.8) ng·L−1), tuberculous PE (73.0 (33.9–131.5) ng·L−1) and transudative PE (36.1 (22.3–74.2) ng·L−1), all with p<0.01. It should be mentioned that, although ENA-78 concentration in infectious PE was much higher than those in the other PEs, we were unable to identify a cutoff value for ENA-78 that could be used to diagnose infectious PE.

Comparison of concentrations of epithelial neutrophil-activating peptide (ENA)-78 in pleural effusions with different aetiologies. #: p = 0.001; ***: p<0.001. Statistical analysis by Kruskal–Wallis one-way ANOVA on ranks.
Concentrations of epithelial neutrophil-activating peptide (ENA)-78, myeloperoxidase (MPO) and neutrophil elastase (NE) in pleural effusions and sera
Similar results were observed with MPO and NE studies, and the concentrations of both these enzymes changed in parallel among the groups. Additionally, as shown in table 2⇑, the concentrations of both MPO and NE in infectious PE were significantly higher than those in the any other groups (all p<0.01).
The comparisons of levels of ENA-78, MPO, and NE in PEs with their corresponding concentrations in sera are also shown in table 2⇑. The concentrations of ENA-78, MPO and NE in sera were not different from one another among the four groups (all p >0.05). ENA-78 concentration was much lower in PEs than in sera from patients with malignant, tuberculous and transudative PEs (all p<0.05); in contrast, ENA-78 in infectious PE tended to be increased when compared with the corresponding serum but the difference did not reach statistical significance (p = 0.102). MPO concentration was much lower in malignant and transudative PEs, and higher in tuberculous and infectious PEs than in their corresponding sera (all p<0.05). NE concentration in malignant, tuberculous and infectious, but not transudative, PE, was much higher than that in their corresponding serum.
Correlation between ENA-78 and inflammatory cells, MPO and NE
We noted that pleural ENA-78 levels were positively correlated with the number of neutrophils (n = 75, r = 0.526; p<0.001)(fig. 2a⇓) but not with lymphocytes (n = 75, r = 0.167; p = 0.153), macrophages (n = 75, r = 0.022; p = 0.854), mesothelial cells (n = 75, r = -0.250; p = 0.066) and malignant cells (n = 13, r = 0.017; p = 0.957). It was further noted that the pleural ENA-78 levels were positively correlated with PE MPO levels (n = 75, r = 0.714; p<0.001)(fig. 2b⇓) and PE NE levels (n = 75, r = 0.739; p<0.001)(fig. 2c⇓). The correlations between pleural ENA-78 and neutrophil numbers, MPO levels and NE levels within the different diagnostic groups are shown in table 3⇓. It was noted that PE ENA-78 levels were positively correlated with MPO levels and NE levels even when PEs with various aetiologies were analysed separately.

The concentrations of epithelial neutrophil-activating peptide (ENA)-78 correlated with a) neutrophil numbers (n = 75, r = 0.526; p<0.001), b) myeloperoxidase (MPO; n = 75, r = 0.714; p<0.001) and c) neutrophil elastase (NE; n = 75, r = 0.739; p<0.001) in pleural effusions. Correlations were determined by Spearman’s rank correlation coefficients.
Correlation of ENA-78 and neutrophil numbers or neutrophil activation markers in pleural effusion (PE)
Infectious PE was chemotactic for neutrophils
The finding that the concentration of pleural ENA-78 correlated best with the number of neutrophils prompted us to test the chemoattractant activity of PEs on human neutrophils. The results showed that exudates, especially empyema, but not transudates, exerted a potent chemoattractant activity for neutrophils (fig. 3⇓). In order to determine whether ENA-78 was responsible for the migration of neutrophils, the ability of an anti-ENA-78 mAb to neutralise the chemoattraction of neutrophils was tested. The anti-ENA-78 mAb significantly suppressed neutrophil chemotaxis in all exudative PE groups. These data provided indirect evidence that ENA-78 was capable of inducing neutrophil recruitment into the pleural space.

Exudative pleural effusions are chemotactic for neutrophils in vitro. Pleural effusions from patients with lung cancer (n = 5), tuberculous pleurisy (n = 5), empyema (n = 5) and with transudates (n = 5) were used to stimulate chemotaxis of peripheral blood neutrophils isolated from healthy adults. ENA-78: epithelial neutrophil-activating peptide. Data are expressed as percentage of control. □: chemotaxis in the absence of anti-ENA-78 monoclonal antibody; ░: irrelevant isotype control; ▓: chemotaxis in the presence of anti-ENA-78 monoclonal antibody. #: p<0.01 compared with the corresponding group without anti-ENA-78 monoclonal antibody; ¶: p<0.01 compared with each of the other pleural effusion groups. The comparisons were determined by Kruskal–Wallis one-way ANOVA on ranks.
Recruitment of neutrophils into PE caused by ENA-78
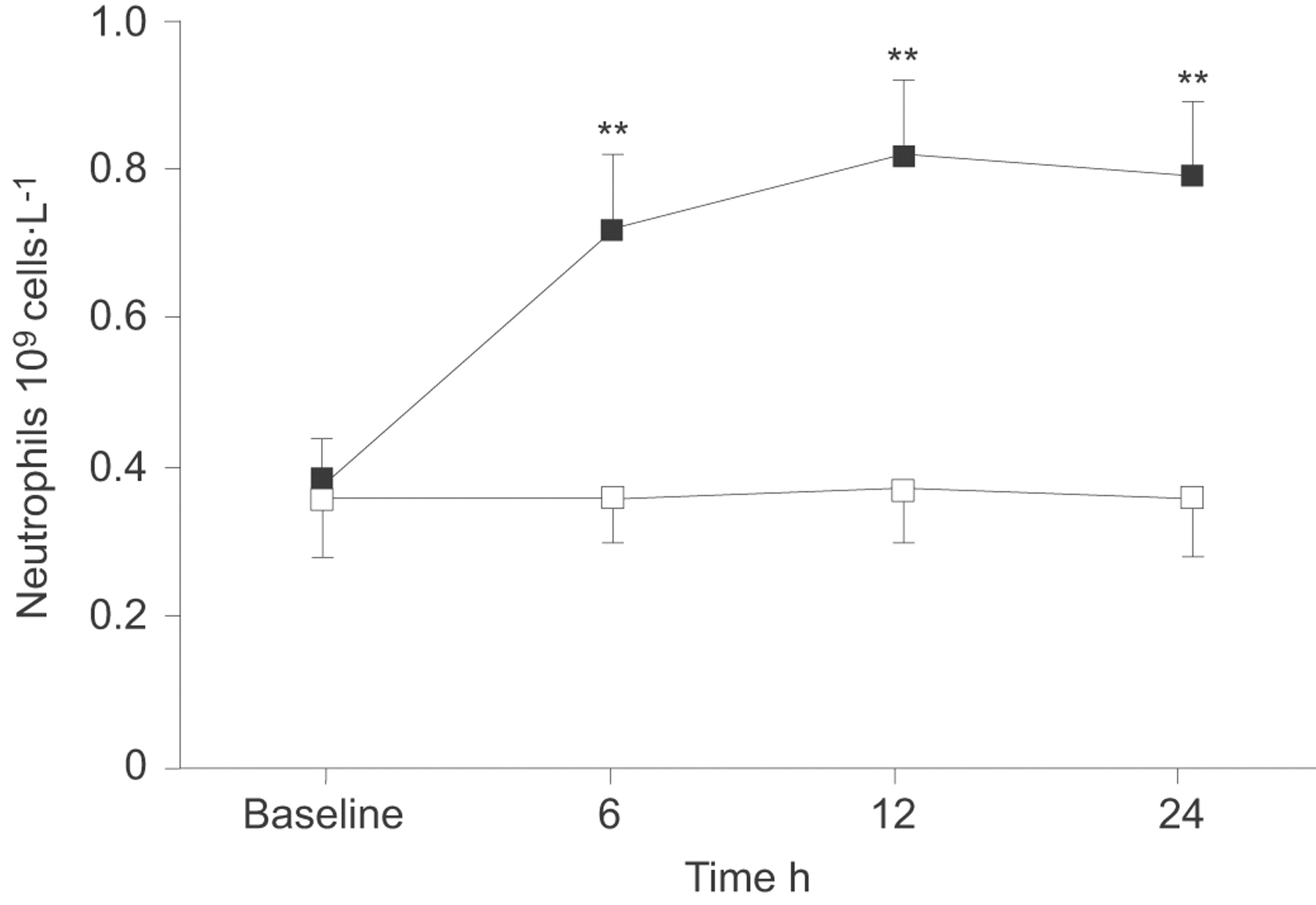
In order to investigate the direct chemoattractant capacity of ENA-78 to recruit neutrophils in vivo, 10 μg human recombinant ENA-78 were injected into the pleural space of patients with tuberculous pleurisy, and then observed the changes in neutrophil numbers. Compared with the baseline value (0.38×109±0.06 cells·L−1), a significant increase in the number of neutrophils started to be observed at 6 h (0.72×109±0.10 cells·L−1; p<0.01) after intrapleural ENA-78 injection; The number of neutrophils increased with time, reaching a maximum at 12 h (0.82×109±0.10 cells·L−1; p<0.01), and lasting for at least 24 h (0.79×109±0.10 cells·L−1; p<0.01) (fig. 4⇓). After vehicle only was injected into the pleural cavity, we did not observe increases in neutrophil counts in PE obtained at three time points when compared with baseline measurement before injection (all p>0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Changes of neutrophil numbers in pleural effusion from patients with tuberculous pleurisy, who were intrapleurally injected with recombinant human epithelial neutrophil-activating peptide (ENA)-78 (▪, n = 5) or vehicle (□, n = 5). Data are presented as mean±sem. **: p<0.01 compared with within-group change from baseline measurements determined by one-way repeated-measures ANOVA.
DISCUSSION
The development of PE is often associated with an increase of inflammatory cells in the pleural space. PEs caused by diverse disease entities are usually present with the predominance of a particular type of leukocyte. Infectious PEs, including empyema and parapneumonic effusion, are typically associated with an influx of neutrophils, whereas tuberculous and malignant PEs are rich in lymphocytes 5, 6.
Inflammation within the pleural space could be mediated by a number of proinflammatory molecules. ENA-78 is a 78-amino-acid protein displaying the four highly conserved cysteine residues that are common feature of all CXC chemokines, including IL-8. The overall amino acid sequence identity of ENA-78 with IL-8 is only 22% 15. ENA-78 is a potent activator of neutrophil function, inducing chemotaxis, enzyme release and rise of intracellular calcium 9 by acting via the type-B IL-8 receptors 16 and, thus, may play an important role in the recruitment of leukocytes to inflammatory lesions in a manner similar to that demonstrated for IL-8 and other neutrophil-activating proteins 17, 18.
We did not identify the cellular origins of pleural ENA-78 in the present study. The primary aim of this study was to explore the presence of ENA-78 in PE, the chemoattractant activity of PE ENA-78 on neutrophils, and the relationship of ENA-78 to the activation state of neutrophils in PE. Originally, ENA-78 was characterised from the IL-1β- and tumour necrosis factor (TNF)-α-stimulated alveolar type II epithelial cell line A549 9. Now, it is well known that ENA-78 expression is inducible by a variety of inflammation mediators, including lipopolysaccharide, IL-1 and TNF-α, in epithelial cells 9, 19, monocytes 20, 21, platelets 22, endometrial stromal cells 23, endothelial cells 24 and macrophage 25. In the present study, our data have shown for the first time that ENA-78 could be detected by ELISA in PEs and that the concentration of ENA-78 in infectious PE was significantly higher than those in malignant, tuberculous and transudative PEs. These findings suggested that more pleural sources of ENA-78 exist in infectious patients. Local production has been reported for some chemokines, such as IL-8 26, 27, monocyte chemotactic peptide-1 27, 28 and macrophage-derived chemokine 29, in inflammatory or neoplastic pleural disorders. Likewise, PE ENA-78 can also be produced by inflammatory cells recruited into the pleural space. Interestingly, although ENA-78 concentration was much lower in PEs than in sera from patients with malignant, tuberculous and transudative PEs, ENA-78 in infectious PE tended to be increased when compared with the corresponding serum. Taken together, these results favour the concept of a local production of ENA-78 in infectious PE, rather than a passive diffusion of this chemokine from plasma to the pleural compartment. Alternatively, it was not possible for ENA-78 to exude into peripheral circulation from the pleural space in patients with malignant, tuberculous or transudative PE, since ENA-78 concentrations in these PEs were much lower than those in the corresponding sera.
The mechanism by which neutrophils infiltrate into pleural cavity has not yet been elucidated completely. The finding of higher concentration of ENA-78 in infectious PE than in any of the other groups was consistent with the massive neutrophilic infiltration of PE in this condition. If ENA-78 was the major factor contributing to neutrophil influx into the pleural space, strong correlations between PE neutrophil counts and ENA-78 would be expected. As a matter of fact, our results have demonstrated a positive correlation between neutrophil numbers and ENA-78 level, when all PEs studied were analysed together. Indeed, an in vitro migration assay in the present study further confirmed that exudative PEs could induce the migration of neutrophils and that an anti-ENA-78 mAb inhibited the ability of the PEs to stimulate neutrophil chemotaxis. The important findings in this study also showed that intrapleural administration of 10 μg human recombinant ENA-78, not vehicle, to patients with tuberculous PE produced a marked progressive influx of neutrophils into pleural space. Therefore in the present study, we have provided the direct evidence, for the first time, that intrapleural injection of ENA-78 was able to chemoattract neutrophil recruitment into the pleural space.
We noted that the effect of inhibiting ENA-78 in infectious pleural fluid only reduced neutrophil chemotaxis by ∼50%, suggesting that ENA-78 is one of several chemokines that aid neutrophil recruitment into the pleural space. Actually, it has been reported in earlier studies that IL-8 concentrations in infectious PE were much higher than those in PEs with any other causes, and that there was a significant positive correlation between pleural IL-8 concentrations and neutrophil counts 27, 30. Moreover, it has been demonstrated that IL-8 was a major neutrophil chemotactic factor in pleural liquid of patients with empyema 27, 30. Since ENA-78 and IL-8 exhibit different patterns of expression in cells, such as human monocytes and endothelial cells 20, 24, the comparison of chemotactic activity between ENA-78 and IL-8 needs to be established in a future study. In addition, it is likely that backup redundancy exists among the many chemokines. It is also necessary to design in vivo animal experiments using gene knockout mice or chemokine antagonists to explore such redundancy in pleural infection.
Neutrophils release reactive oxygen and nitrogen species into the extracellular milieu, in addition to releasing their granular content, which contains several serine and neutral proteases that can produce tissue injury. Among the neutrophil proteases, NE has been implicated in both chronic and acute inflammatory damage, and oxidant species, partly produced by the action of another granular enzyme, MPO, potentiate its effects 31, 32. Therefore, it is likely that the products derived from neutrophil activation are related to the evolution of infectious PE from the noncomplicated to the complicated state. The effect of ENA-78 on neutrophil degranulation has been demonstrated in vitro 9, 33 and there are also data to support its occurrence in vivo 25. In agreement with earlier studies reported by other authors, we found the highest PE concentrations of both MPO and NE in infectious PEs 34, with a profile similar to that for ENA-78. In the present study, we have shown that both MPO and NE correlated positively with ENA-78, thus suggesting a role for ENA-78 in pleural neutrophil degranulation.
It should be noted that although the ENA-78 level in infectious PE was higher than that in PE caused by the other causes, it is not possible to differentiate infectious PE from other kinds of PE due to the wide distribution and obvious overlap of these chemokine concentrations among each group.
In conclusion, compared with noninfectious PE, ENA-78 appeared to be increased in infectious PE. ENA-78 was able to induce neutrophil infiltration into pleural space and might be responsible for pleural neutrophil degranulation.
Support statement
This study was supported in part by research grants 30660064 and 30872343 from the National Natural Science Foundation of China; in part by research grant 0728137 from the Natural Science Foundation of Guangxi Zhuang Autonomous Zone; and in part by research grant 200621 from the Bureau of Health, Guangxi Zhuang Autonomous Zone, China.
Statement of interest
None declared.
- Received July 22, 2008.
- Accepted November 6, 2008.
- © ERS Journals Ltd
References










