





Abstract
Clinical and experimental studies indicate an association between chorioamnionitis and bronchopulmonary dysplasia in preterm infants. The present authors hypothesised that, in the rabbit, antenatal infection may impair lung development after birth, despite effective maternal antibiotic therapy.
Pregnant rabbits received an intra-uterine inoculation of 103 Escherichia coli colony forming units or vehicle at the end of gestation (day 29). Intravenous ceftriaxone therapy was initiated 8 h after inoculation for a period of 8 days. Pups born between 60 and 84 h after inoculation were kept with their mother until sacrifice on days 0, 1, 5, 8 and 15.
Blood cultures from antenatally infected animals were sterile at birth. Postnatal growth was significantly impaired by day 8. Lung morphometry showed a significant decrease of alveolar surface density and interstitial density, with a significant increase of alveolar airspace volume, indicating impaired alveolarisation for the first 2 weeks of postnatal life. Inflammatory and apoptotic processes were not detected in the lung at birth or subsequently.
Intra-uterine infection in rabbits is, therefore, responsible for concomitant postnatal growth retardation and abnormal pulmonary development despite early and effective antenatal antibiotic therapy. This may constitute an alternative model to study the consequences of antenatal infection on postnatal growth and lung development.
Despite recent major advances in the management of premature infants, including surfactant administration and noninvasive mechanical ventilation, most immature neonates, often weighing less than 1,000 g at birth, are prone to develop lung sequelae corresponding to bronchopulmonary dysplasia (BPD) 1. BPD is mainly related to arrested lung development, including decreased expression of key factors regulating alveolar and vascular development 2, 3, and marked reduction in total alveolar number 4. Numerous risk factors have been identified, including mechanical ventilation, oxygen therapy, patent ductus arteriosus and nosocomial infection 5. However, there is growing evidence that antenatal infection may play a crucial role in the pathogenesis of BPD 6. Watterberg et al. 7 reported that 80% of infants without respiratory distress syndrome, who nevertheless developed BPD, were born in a context of chorioamnionitis. Moreover, maternal stage of chorioamnionitis was correlated with severity of BPD 8. Experimental models have highlighted the role of inflammation induced by intra-amniotic endotoxin in altered lung development in fetal sheep 9, 10 or rats 11. However, these models of selective amniotic fluid inflammation are quite different from the characteristic features associated with the pathophysiology of chorioamnionitis, which consist of active bacterial infection.
In the present study, a model of intra-uterine infection in the rabbit was used to evaluate the effects of infection on neonatal lung development. This model of Escherichia coli chorioamnionitis represents an active infection with relevance to human pathology, as E. coli is involved in numerous neonatal infections 12, 13. The impact of intra-uterine infection on newborn lung development was evaluated by studying lung morphometry and cell death, and the impact on inflammation status was studied by inflammatory cell counts and interleukin (IL)-6 determination in bronchoalveolar lavage (BAL) fluid from birth until 2 weeks of postnatal life.
METHODS
Antenatal infection
The study was conducted in pregnant and newborn New Zealand rabbits, with the RS218 (O18: K1: H7) strain of E. coli K1 isolated from the central nervous system (cerebrospinal fluid) of a human newborn. This bacterial strain has already been characterised for virulence factors and invasion genes 14. The minimum inhibitory concentration (MIC) of this strain for ceftriaxone was 0.06 mg·L−1. The intra-uterine inoculation protocol, as previously described 12, was approved by the Animal Care Committee of the University of Nantes experimental therapy department (Nantes, France). A total of 14 female New Zealand white rabbits weighing 3.7–4.5 kg (CEGAV, Saint Marc d'Egrenne, France) were obtained 10 days before the end of their usual 31–33-day gestation period and housed at the Nantes School of Medicine (Nantes, France) in appropriate cages with a special place for nidation. They were provided ad libitum with water and food consisting of antibiotic-free granules. On day 29 of gestation, the animals were anesthetised with 25 mg·kg−1 of intramuscular ketamine. Laparotomy was then performed via a 2-cm vertical midline incision below the gravid uterus. After exposing the uterus, a single injection of E. coli inoculum (103 colony forming units (CFU) in 1 mL of 0.9% NaCl solution) or sterile vehicle was injected under visual control into one of the uterine horns facing a fetoplacental unit. Slight aspiration was performed before injection to verify correct intra-amniotic positioning of the needle. The incision was then closed in layers and the animals were returned to their cages. All pregnant rabbits (control and infected; n = 7 for both groups) were treated with 300 mg·kg−1·day−1 intravenous ceftriaxone (Rocephine®; Roche, Meylan, France) via a catheter inserted into a marginal vein of the ear. Treatment began 8 h after intra-uterine inoculation and lasted a total of 8 days. These experimental conditions provided a high rate of antenatal infection, due to contiguous propagation of infection 12, with a sufficient number of live births to allow evaluation of postnatal development. Ceftriaxone was chosen due to the current increase in penicillinase-secreting E. coli strains. Spontaneous birth occurred 60–84 h after inoculation and rabbit pups were suckled until euthanasia. They were exsanguinated at birth (day 0), day 1, day 5, day 8 and day 15, in both the control and infected groups under deep anesthesia with intraperitoneal pentothal. Blood was obtained by cardiac puncture only after animals failed to blink an eyelid or move a leg muscle following local stimulation. A total of five to 10 rabbit pups per group were used for either BAL or morphometric study.
Assessment of lung morphometry
Lungs were gently extracted en bloc and fixed with 4% paraformaldehyde through a polyethylene tracheal cannula at a constant pressure of 30 cm water for 10 mins. The trachea was then ligated and the lung was immersed in 4% paraformaldehyde for 24 h. Lung volumes were measured by the displacement method in the fixative solution 15. Both lungs of each rabbit were paraffin-embedded and 4-μm frontal sections were cut in the medial part of the lungs and stained with hematoxylin-eosin-saffron (HES). All lung lobes were used. All morphometric evaluations were performed by one observer blinded to group assignment. A microscope (Leitz, Weszlar, Germany) connected to a television screen by a video camera (Sony, Tokyo, Japan) was used. Volume densities of pulmonary parenchymal structures (alveolar airspace, airways, blood vessels >20 μm in diameter and interstitial tissues) and alveolar surface density were measured using the point counting and mean linear intercept methods, as previously described 16. Light-microscope fields were quantified at an overall magnification of ×400, using a 42-point, 21-line eyepiece graticule placed on the television screen. A total of 20 fields (10 per lung) were evaluated per animal by a systematic sampling method from a random starting point. Morphometric evaluations were repeated by a second independent observer in order to test reproducibility. To correct for shrinkage associated with fixation and paraffin processing, area values were multiplied by 1.22, a factor calculated during a previous evaluation 17. All morphometric data were expressed as relative and absolute values, as described by Burri et al. 18. Relative values (volume density or surface density) were those obtained directly from morphometric measurements of tissue sections. Absolute values (total volume or surface area per lung) were determined by multiplying the relative values by lung volume.
Assessment of inflammation
BAL
BAL was performed as previously described 17. Briefly, rabbits were exsanguinated and the thorax was opened to expose the lungs and trachea. A short length of tubing was inserted into the trachea and ligated. A total of 8–10 separate aliquots of warm saline (37°C) were used for BAL. Volumes ranged from 1 mL per aliquot for just-born rabbits to 6 mL per aliquot for day-15 rabbits. Total BAL volumes were 10, 16, 21 and 30 mL for rabbits at days 0, 5, 8 and 15, respectively. Total lavage volume was measured and aliquots were saved for subsequent analysis. Cells recovered from alveolar lavage fluid after centrifugation (300 ×g for 7 min) were resuspended in isotonic saline at a concentration of 106 cell·mL−1. Cytodifferentiation was performed on cytospin preparations stained with Diff-Quick (Scientific Products, McGraw Park, IN, USA).
Histologic and immunohistochemical evaluation of inflammation
Lung inflammation was evaluated on HES-stained sections of at least eight fetuses in each group with respect to neutrophil infiltrates, congestion and presence of oedema.
The placenta of each fetoplacental unit was rinsed with sterile saline and fixed in 3% formalin and embedded in paraffin. Paraffin sections (5 μm) were stained with standard hematoxylin and eosin techniques. In order to further study the inflammatory response, immunodetection of macrophages was performed on placentas. A mouse monoclonal anti-human monocyte-macrophage antibody (MCA874G; Serotec, Oxford, UK) was used for macrophage detection.
IL-6 determination
Levels of immunoreactive IL-6 were determined in plasma and BAL by ELISA using a commercially available kit (Diagnostic Products Corp., CA, USA).
Cell death
Cell death was assessed in at least four pups of each group, using a DNA staining assay. The DNA-specific dye Hoechst 33258 (Calbiochem, Merck Biosciences, VWR, Fontenay-sous-Bois, France), 1 μg·mL−1 in Hank's balanced salt solution without phenol red, was used on formalin-fixed paraffin sections of infected and infected-and-treated newborns (n = 5). Sections were mounted with Prolong Antifade medium (Molecular Probes, Eugene, OR, USA). Fluorescence was observed with a fluorescence microscope (Axiovert 200-M; Carl Zeiss, Göttingen, Germany) equipped with an ApoTome slider, which eliminates image blurring. Cells were visualized with a ×63/1.4 oil immersion lens. Image processing was performed using an AxioCam MR CCD camera and AxioVision 4.0 software (Carl Zeiss, Göttingen, Germany).
Bacteriological analysis
Blood cultures
At the time of sacrifice, blood was withdrawn from the hearts of dams and pups for quantitative cultures; 100-μL blood samples were mixed with 900 μL of heparinized (5,000 IU·L−1) physiological serum. After centrifugation, 100 μL of the pellet were plated on MH agar (Difco; Becton Dickinson, Franklin Lakes, NJ, USA) and incubated at 37°C for 24 h before performing bacterial counts. This method allows detection of an inoculum of 102 CFU·mL−1.
Pulmonary and placental cultures
At the time of sacrifice, lung and placenta were homogenised in 500 μL of sterile saline solution and diluted to 10−2 and 10−4 to prevent the possibility of carry-over. Next, 50 μL of undiluted homogenate and 50 μL of each dilution were spread on EμMB plates using a spiral system plater (Interscience, BV, Louvain, Belgium). After 24 h of incubation at 37°C, viable bacteria were counted and the results were expressed as log10CFU·g−1 of tissues. The lower detection limit for this method was 1 CFU per 50 μL of undiluted tissue homogenate.
Two pregnant rabbits were sacrificed 8 h after inoculation and placental and fetal blood cultures (n = 17) were performed in order to evaluate the course of infection after initiation of antibiotic therapy.
Statistical analysis
Results are expressed as mean±sd. Global differences were evaluated using a Kruskal–Wallis test and comparisons between the two groups were performed with a Mann–Whitney test. Percentages were compared using Fisher's exact test. A p-value p<0.05 was considered to be significant.
RESULTS
Effects of E. coli infection on rabbits
Maternal aspects: pregnancy outcome, survival, bacteriological results
There was no loss of pregnancies after intra-uterine inoculation. At the time of initiation of antibiotics (8 h after bacterial challenge), maternal blood cultures were sterile but 5 (29%) out of 17 fetal blood cultures were positive and 15 (88%) out of 17 placental cultures were positive with a mean quantitative value of 4.5±1.4 log10CFU·g−1 of tissue. By contrast, placental cultures remained negative at birth and all cultures of maternal blood sampled after 8 days of antibiotics were sterile, indicating that maternal infection was controlled. All but one of the dams survived during the follow-up period.
Neonatal aspects: survival, growth, bacteriological results
At birth, mortality was higher in the infected group than in the control group with 33 (50%) out of 66 stillborn infected pups from seven infected dams and 13 (25%) out of 51 stillborn control pups from seven control dams (p = 0.01). Mean number of fetuses per dam was 8.3±2.3, (7.3±2.5 in the control group and 9.2±1.8 in the infected group; p = 0.12). The litter size after excluding stillbirths was 5.4±1.9 in the control group and 4.8±1.4 in the infected group; p = 0.4). Twelve pups died spontaneously after birth and before day 11 in the infected group, while only one died in the control group over the same period (p<0.001). Mean birth weight did not differ between infected and control newborns. Postnatal growth was significantly different between groups at day 8 and day 15, as mean body weight was about 60% lower in the infected group (p<0.05; fig. 1⇓). All blood cultures from pups collected at birth or later were sterile.

Growth of newborn rabbits in infected and control groups. Pregnant rabbits received an intra-uterine inoculation of 103 Escherichia coli or saline on day 28 of gestation followed by antibiotics (see Methods section). Pups were born spontaneously and were suckled by their mothers until sacrifice on days 0, 1, 5, 8 and 15. Growth was significantly different between infected (▪) and control (•) animals on days 8 and 15. Data are expressed as mean±sd. *: p<0.05.
Morphometric evaluation of lung structures
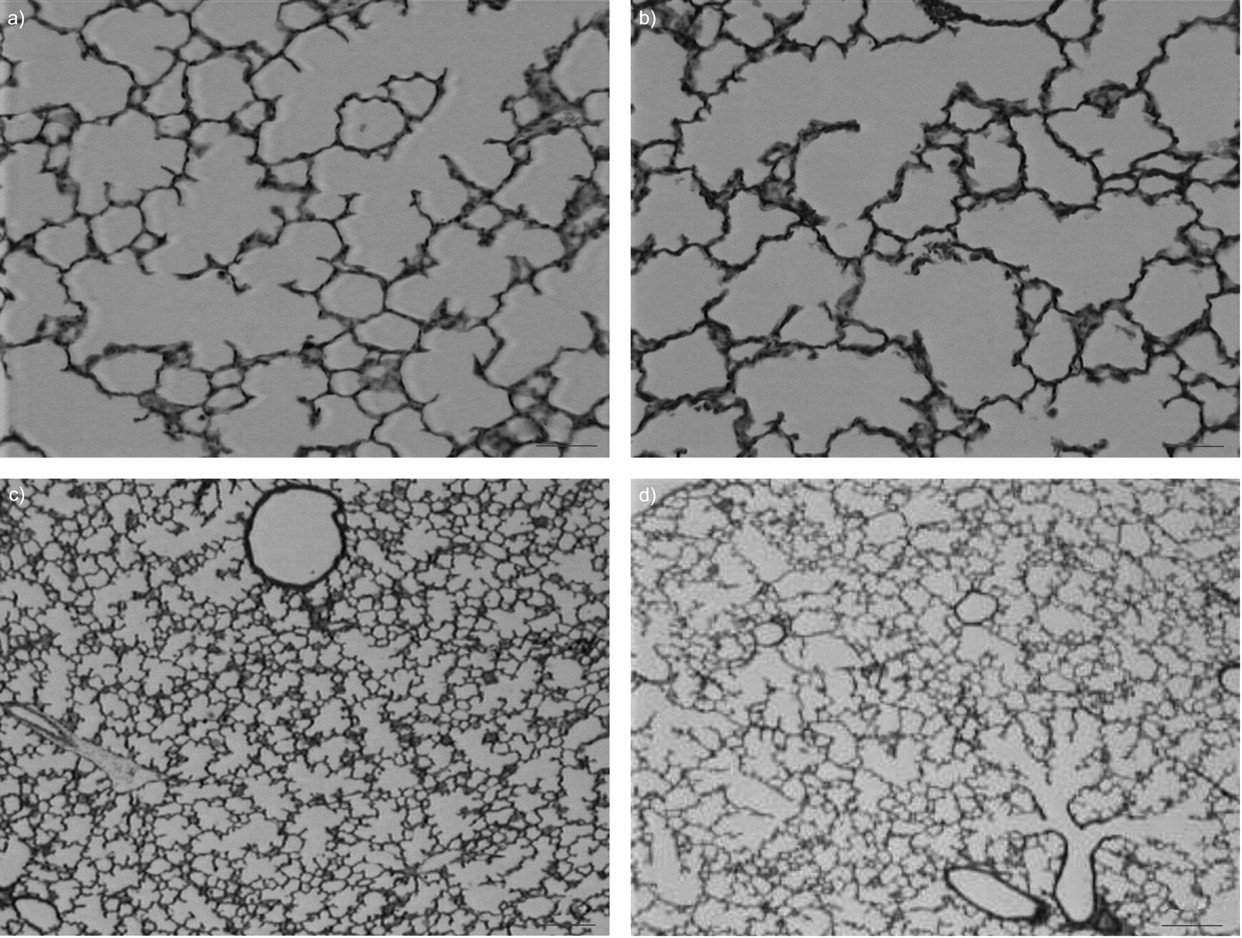
Examination of postnatal lung sections under light microscopy showed that infected rabbit lungs exhibited a diffuse, simplified lung structure with enlarged air spaces and fewer secondary septa (fig. 2⇓). Quantification by morphometric analysis (all results from day 0 to day 15 are presented in figs 3⇓–⇓⇓6⇓) highlighted the following features.

Histological appearance of the lung showing the effect of intra-amniotic bacterial inoculation on airspace formation in rabbit pups. A timed pregnant rabbit received an inoculum of 103 colony forming units of Escherichia coli in the uterine horns followed by antibiotics, and delivered at normal term. Rabbit pups were killed postnatally at different times. Light micrographs of haematoxylin-eosin-saffron-stained 4-μm sections of lung parenchyma in control (a, c) and infected (b, d) pups at day 5 (a, b) and day 8 (c, d). Infected rabbit lungs exhibited diffuse, simplified lung structure with enlarged air spaces and fewer secondary septa. a and b) Scale bars = 50 μm; c and d) scale bars = 100 μm.

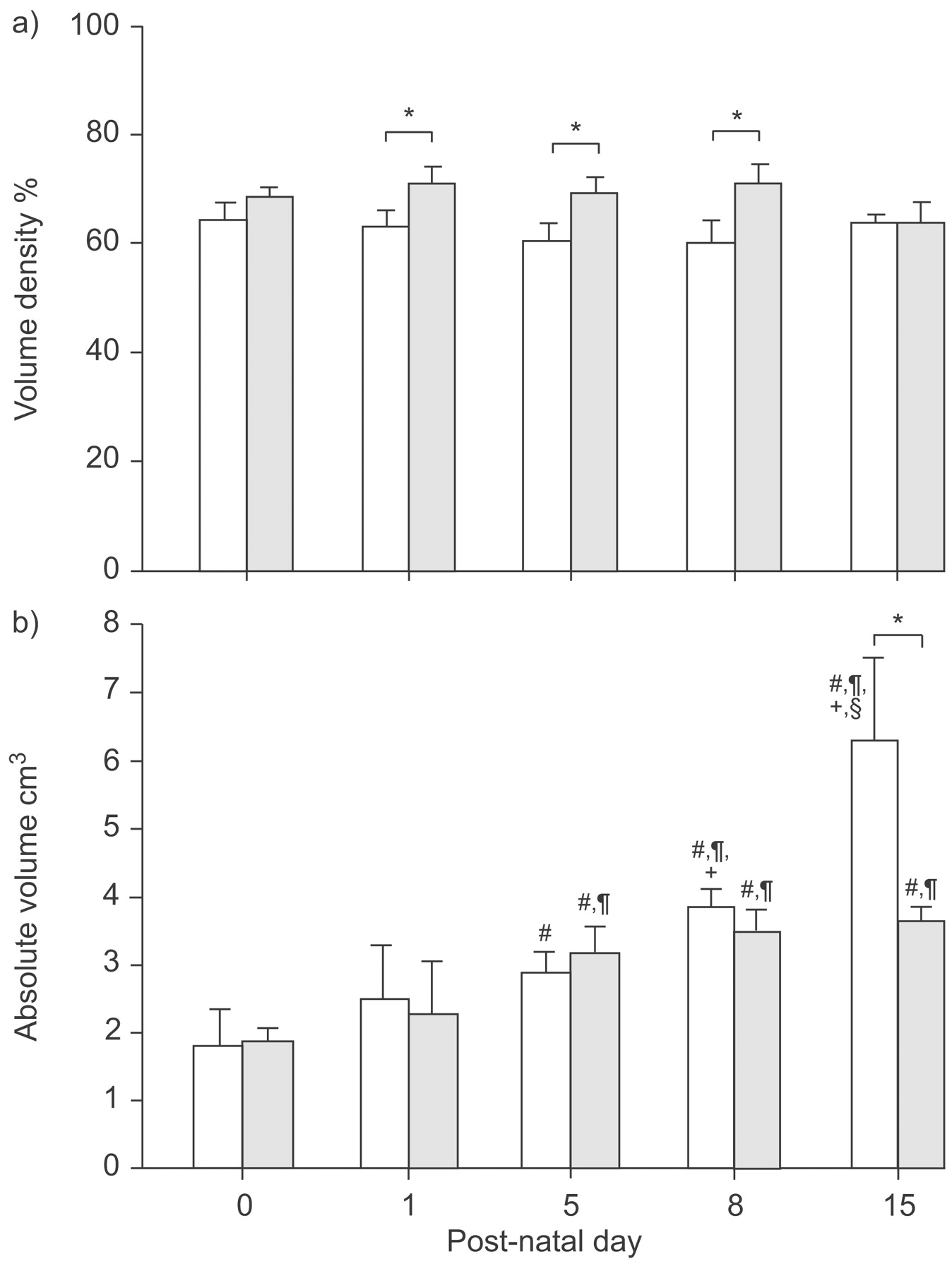
Absolute and specific lung volumes. A timed pregnant rabbit received an inoculum of 103 CFU Escherichia coli (░) or its diluent (□) in the uterine horns followed by antibiotics and delivered at normal term. Rabbit pups were sacrificed postnatally on different days. Lung volume of infected and control newborn rabbits was measured by the displacement method in the fixative solution. Specific lung volume was calculated by dividing lung volume by body weight and multiplying the result by 100. Values are presented as mean±sd of measurements on three to six individual lungs for each group. Lung volume increased constantly in control pups but stopped increasing on day 5 in infected pups. Specific lung volume decreased in controls and in infected pups, although more slightly in infected pups, resulting in an increased specific volume in infected pups on days 8 and 15. #: p<0.05 compared with day 0; ¶: p<0.05 compared with day 1; +: p<0.05 compared with day 5; §: p<0.05 compared with day 8. *: p<0.05 when infected are compared with controls.

Alveolar surface area density and absolute area in rabbit pup lungs antenatally exposed to Escherichia coli (E. coli) infection. A timed pregnant rabbit received an inoculum of 103 CFU Escherichia coli (E. coli) (░) or its diluent (□) in the uterine horns followed by antibiotics and delivered at normal term. Rabbit pups were sacrificed postnatally on different days. Results are expressed as a) density or b) absolute values. Antenatal infection significantly decreased alveolar surface area density from day 1 and prevented the increase in absolute value from day 5 to day 15. Data are presented as mean±sd of measurements on three to six individual lungs for each group. #: p<0.05 compared with day 0; ¶: p<0.05 compared with day 1; +: p<0.05 compared with day 5; §: p<0.05 compared with day 8. *: p<0.05 when infected are compared with controls.

Alveolar airspace volume density and absolute values of rabbit pup lungs antenatally exposed to Escherichia coli (E. coli) infection. A timed pregnant rabbit received an inoculum of 103 CFU E. coli (░) or its diluent (□) in the uterine horns followed by antibiotics and delivered at normal term. Rabbit pups were sacrificed postnatally on different days. Results are expressed as a) airspace volume density and b) absolute values. Antenatal infection significantly decreased alveolar volume density from day 1 to day 8 and arrested the increase in absolute value from day 5 to day 15. Data are presented as mean±sd of measurements on three to six individual lungs for each group. #: p<0.05 compared with day 0; ¶: p<0.05 compared with day 1; +: p<0.05 compared with day 5; §: p<0.05 compared with day 8.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relative, absolute, and specific values of interstitium volume in rabbit pup lungs antenatally exposed to Escherichia coli (E. coli) infection. A timed pregnant rabbit received an inoculum of 103 CFU E. coli (░) or its diluent (□) in the uterine horns followed by antibiotics and delivered at normal term. Rabbit pups were sacrificed postnatally on different days. Results are expressed as a) density and b) absolute values. Antenatal infection decreased volumetric density of alveolar interstitium on days 5, 8 and 15 and arrested its growth from day 5 in absolute values. Data are presented as mean±sd of measurements on three to six individual lungs for each group. #: p<0.05 compared with day 0; ¶: p<0.05 compared with day 1; +: p<0.05 compared with day 5; §: p<0.05 compared with day 8. *: p<0.05 when infected are compared with controls.
Lung volume
Absolute lung volume increased significantly in both groups from day 0 to day 5 (p<0.05) but stopped increasing at day 5 in infected pups (no significant difference between day 5 and day 8 or day 15). In contrast, absolute lung volume continued to increase in the control group, as the value at day 8 was significantly higher than at day 5 and the value at day 15 was significantly higher than at day 8. Consequently, absolute volume was lower in infected animals than in the control group, with a significant difference on days 8 and 15 (p<0.05). Specific lung volume tended to increase slightly in both groups from day 0 to day 1 although this did not reach significance, and then decreased significantly in the control group (Kruskal–Wallis test; p<0.05), while no significant decrease was observed in the infected group (Kruskal–Wallis test; p = 0.08). Consequently, specific lung volume was higher in infected than in control pups on days 8 and 15 (p<0.05; fig. 3⇑).
Alveolar surface area
Infected rat pups had a significantly lower (about 25%) alveolar surface density (p<0.05) from day 1 to day 15. The absolute alveolar surface area did not increase between days 5 and 15 in infected pups in contrast with a marked and significant increase in control rabbits (p<0.05). As a result, absolute alveolar surface area was significantly lower in infected pups compared with controls on days 5 (-30%), 8 (-40%) and 15 (-55%; p<0.05; fig. 4⇑).
Alveolar airspace volume
The alveolar airspace volume density was significantly increased in infected pups compared with controls (p<0.05 from day 1 to day 8). Absolute alveolar volume did not increase in infected pups between days 5 and 15 but the difference compared with controls was significant only at day 15 (p<0.05; fig. 5⇑).
Interstitial volume
The interstitial volume density was decreased in infected pups from days 5 up to 15. In alveolar surface area and volume absolute values, an increase of absolute interstitial volume was observed from days 5 to 15 in control pups. In contrast, absolute interstitial volume did not increase in infected pups between days 5 and 15. Consequently, a significant difference between infected and control pups from -40% on day 1 to -52% on day 15 was observed (fig. 6⇑).
Evaluation of inflammatory response
Histological examination
Histological examination of chorionic plates at birth showed diffuse neutrophil infiltrates, suggesting an inflammatory process. The macrophage nature of infiltrating cells was supported by immunohistochemical staining with MAC 874 (data not shown).
Histological examination on HES-stained sections of infected pup lungs on days 0, 8 and 15, or of paired control animals, did not show any obvious sign of inflammation, as oedema or inflammatory infiltrate were not observed (fig. 2⇑).
BAL cellularity
Total cellularity in BAL did not differ significantly between infected and control groups. The percentage of polymorphonuclear neutrophils was slightly but significantly increased on days 0 and 5 in infected pups (table 1⇓).
Effects of antenatal infection on bronchoalveolar lavage(BAL) cell count from day 0 to day 15
IL-6 assay
IL-6 levels determined in plasma and in BAL did not differ between infected and control pups (table 2⇓).
Effects of antenatal infection on interleukin(IL)-6 levels in serum and bronchoalveolar lavage (BAL) fluid on post-natal days 0, 5 and 15
Cell death
Staining with Hoechst 33258 dye demonstrated chromatin condensation or fragmentation in only a few cells at days 1, 5, 8 in both infected and control groups (data not shown). The proportion of apoptotic cells was too low to perform quantitative evaluation in either group.
DISCUSSION
The present study provides evidence for the first time that in vivo intra-amniotic infection by E. coli in the rabbit affects postnatal growth and lung development for a long time, despite early appropriate maternal antibiotic therapy that rapidly eradicated infection.
The first finding was that absolute lung volume increased to a greater extent in control pups than in infected pups, although specific lung volume, i.e. expressed relative to body weight, was higher in infected pups. This finding was the result of increased distensibility that can be observed when alveolar septation is either increased or decreased. Determination of alveolar volume and surface area clearly indicated arrest of lung development; the alveolar airspace volume density was slightly but significantly increased in infected pups whereas alveolar surface density was decreased in infected pups, corresponding to fewer alveoli per unit volume. This was confirmed when these values were reported as absolute values, i.e., taking in account the lung volume; an arrested increase of alveolar surface area and alveolar volume was clearly observed from day 5 to day 15 in infected pups, in contrast with the constant increase observed over the same period in control pups. The decrease in alveolar volume in the presence of a higher specific lung volume in infected pups is indicative of an emphysema-like condition, consistent with the morphological appearance. These data were corroborated by determinations of interstitium volume; interstitium volume density was lower in infected pups, which was probably due to a decreased number of septa (and consequently interstitial tissue) and the increase in absolute interstitium volume also appeared to be arrested in infected pups compared with controls. The present results, therefore, show that the infection–antibiotic treatment procedure provides an alternative model to reproduce key morphometric features of altered lung development similar to those observed in BPD.
This impaired alveolar development mainly occurred after birth but significant differences in alveolar airspace volume and surface density were observed as early as day 1, before any weight difference was observed. This excludes the possibility that this alveolar impairment was exclusively due to growth retardation, at least at this time. Moreover, the absolute alveolar surface area, which is probably the best parameter to assess alveolarisation, was also impaired by day 1. This suggests that antenatal inflammation plays a role in alveolarisation impairment. Epidemiological and clinical data support a strong association between chorioamnionitis, systemic infections, colonisation of the airways and the development of chronic lung disease, especially in preterm infants with mild or no respiratory distress syndrome 19, 20. That impaired lung development resulting directly from in utero infection is supported by experimental findings, mainly obtained in a lamb model of endotoxin-induced inflammation, which accelerated lung maturation and surfactant synthesis 9, 10, 21–23 and generated structural changes in lung parenchyma with a marked reduction of alveolar number 9. A single antenatal endotoxin injection appears to be sufficient to trigger lung maturation in lambs 22 and impaired alveolarisation in rats 11. Moreover, Kramer et al. 24 reported that an acute inflammatory response in the fetal lamb resolved rapidly, although unactivated inflammatory cells persisted in BAL for a longer period of time than observed in the present model. Subsequently, the same authors reported that the increase in apoptotic cells resolved within 72 h.
The present model is based on transient E. coli infection. The study design does not allow verification that each pup was actually infected, which obviously constitutes a limitation. However, these experimental conditions provide a high rate of antenatal infection, which is propagated by contiguity, as already published 12 and this conclusion is supported by the high level of positive placental cultures and high losses in the infected group. The present model of transient E. coli infection can, therefore, be considered to induce very similar consequences to those observed in the previous lamb and rat models, as bacteriological cultures were sterile in newborns and dams at birth, indicating that the infectious process was eradicated at birth, while nearly all placental cultures were positive on initiation of antibiotics. The percentage of polymorphonuclear neutrophils in BAL was slightly but significantly increased in infected pups at day 0 and day 5, suggesting transient local inflammation prior to birth, which is consistent with a previous report that early antibiotic therapy after inoculation was sufficient to reverse inflammation 13. Finally, no signs of apoptosis were observed in the lungs, which appears to be consistent with the findings reported by Kramer 24 and the present authors’ previous findings concerning rapid reversal of apoptosis induced by antenatal infection in response to antibiotics 13.
The rabbit has already been used as an experimental model for E. coli intra-uterine infection 12, 13, 25–27. In comparison with other models, the inflammation-induced impairment of alveolar development in the present model is original, as it induced complete and transient infection and uses a species in which the chronology of alveolar development closely resembles that of humans. In both species, the process of secondary alveolar septation starts shortly before birth and continues throughout a prolonged postnatal period. In the rabbit, alveolarisation starts on day 28 for a full term of 31–33 days 28 and continues for a long time after birth with a dramatic increase in the first 2 weeks of postnatal life 29. Similarly, human alveolar development extends from about 36 weeks of pregnancy until the second year of life 30. In this respect, alveolar development in the rabbit more closely resembles that of humans than the other classical animal models such as rats and mice, in which alveolar development is an entirely postnatal process, or sheep, in which it is mainly antenatal. Finally, as this model uses a small inoculum allowing pup survival, it allows evaluation of lung development over a long developmental period that includes most of the active alveolarisation process.
Other causes may account for the observed impairment of alveolar development. First, as a higher number of stillborns was observed in the infected group, toxic substances released as a result of these in utero deaths may have played a potential role on the antenatal alveolar development of the surviving fetuses. Secondly, a high postnatal mortality was observed in the infected group together with a decline in weight gain after day 5. The present model can therefore be considered to be a combination of antenatally acquired impaired alveolar development mainly due to a transient inflammatory process, amplified postnatally by altered growth.
The effects of maternal infection on fetal growth restriction have recently been reported. Infection can lead to host epigenetic modifications of imprinted genes via changes in DNA methylation with developmental modifications, potentially inducing long-term disabilities in various organ systems 31. In the present study, fetal growth was not impaired; birth weight was not significantly different between litters. This loss of weight gain occurred after birth, which is an intriguing finding, as chorioamnionitis or antenatal infections are not currently recognised as factors of impaired postnatal growth. Several hypotheses related to the experiment and more-general factors could explain these findings. First, the number of pups per litter could have been unbalanced between the infected and control groups. However, this was not the case, as the number of fetuses and pups per dam was the same in the two groups. The number of fetuses was slightly but not significantly higher antenatally in the infected group and, due to the large number of stillborns, was slightly but not significantly higher postnatally in the control group, which rules out the hypothesis that more pups in the infected group received less milk due to their number. A second hypothesis is that the quality or quantity of milk could have been insufficient for normal growth of the pups; this could be due to maternal undernutrition resulting from a change in their feeding behaviour due to antenatal infection. The only way to test this hypothesis would have been to perform a cross-nursing experiment. Unfortunately, preliminary experiments revealed that dams refused to adopt other pups, leading to a high level of cannibalism. This impossibility to perform cross-nursing is clearly a limitation of the study. Alternatively, a modification of maternal behaviour leading to less nursing care, which could have profound consequences, including modification of hypothalamic-pituitary-adrenal reactivity in the pups 32, can also hypothesised. This hypothesis cannot be verified in the absence of video recordings. A third hypothesis would be a modification of gut microbial ecology induced by antibiotics, which is known to interfere with weight gain 33. However, this hypothesis is less likely as all rabbits received ceftriaxone antenatally. However, impaired postnatal growth is an essential feature of this experimental model and further investigations must be conducted to elucidate its mechanisms.
There is growing evidence that antenatal infectious/inflammatory processes affect normal alveolarisation in infants 6, 20. The present experimental findings support this hypothesis and provide an experimental model of antenatal infection in the rabbit that reproduces the morphometric features of impaired alveolar development associated with impaired postnatal growth. This model may be useful to investigate the developmental mechanisms responsible for these disorders and to develop strategies to restore alveolarisation.
Statement of interest
None declared.
- Received February 14, 2008.
- Accepted July 23, 2008.
- © ERS Journals Ltd
References










