





Abstract
The transcription factor complex hypoxia-inducible factor (HIF)-1 controls the expression of most genes involved in adaptation to hypoxic conditions. HIF-1 is a heterodimer composed of oxygen-labile HIF-α and constitutively expressed HIF-β subunits. The oxygen-dependent regulation of HIF-α is a multistep process that includes degradation under normoxia but stabilisation, translocation into the nucleus and activation under hypoxic conditions.
The present paper summarises the contributions of optical methods to the understanding of oxygen-dependent regulation of the HIF-1 pathway. The tissue- and cell-specific distribution of HIF-α was visualised immunohistochemically and by immunofluorescence. Transcriptional activity of HIF-1 was monitored using green fluorescent protein as a reporter under control of hypoxia response elements in living cells, spheroids and tumour tissues in living mice. With cyan and yellow variants of green fluorescent protein fused to HIF subunits and regulatory proteins, subcellular distribution, migration and interaction were imaged in vivo by means of fluorescence recovery after photo-bleaching and fluorescence resonance energy transfer. Noninvasive imaging of these cellular and molecular processes by laser scanning microscopy complements ex vivo molecular biology assays and provides an additional spatial and temporal dimension to the understanding of the HIF-1 pathway.
- Fluorescence recovery after photo-bleaching
- fluorescence resonance energy transfer
- green fluorescent protein fusion proteins
- hypoxia-inducible factor-1
- immunohistology
- in vivo imaging
SERIES “HYPOXIA: ERS LUNG SCIENCE CONFERENCE”
Edited by N. Weissmann
Number 3 in this Series
In response to reduced oxygenation, mammalian cells activate hypoxia-inducible factor (HIF)-1, which regulates transcription of genes involved in angiogenesis, erythropoiesis, glycolysis, iron metabolism and cell survival 1. In addition to its role in oxygen homeostasis, in vivo studies have revealed a specific role of HIF-1 during ischaemia, inflammation and various cancers, as well as during development 1.
HIF-1 is a heterodimeric basic helix–loop–helix/PER/ARNT/SIM (PAS) domain transcription factor, composed of a stable and constitutively expressed β-subunit (HIF-1β, also known as aryl hydrocarbon receptor nuclear translocator (ARNT)) and an inducible α-subunit (HIF-1α), the levels of which are controlled by cellular oxygen concentration (fig. 1⇓). In addition to the ubiquitous HIF-1α, the HIF-α family contains two other members, HIF-2α (also referred to as endothelial PAS domain protein-1) and HIF-3α, both of which have a more restricted tissue expression 2, 3. Whereas HIF-2α is directly involved in hypoxic gene regulation, most notably in endothelial cells, HIF-3α might be a negative regulator of hypoxia-inducible gene expression 4.

Structure of hypoxia-inducible factor (HIF)-1 subunits. The human HIF-1α subunit (826 amino acids) and the constitutively expressed HIF-1β subunit (789 amino acids) of the HIF-1 complex belong to the group of basic helix–loop–helix (bHLH)/PER/ARNT/SIM (PAS) proteins. bHLH/PAS domains mediate DNA binding and heterodimerisation of the subunits. Transactivation of the HIF-1 complex is mediated by the N-terminal transactivation domain (N-TAD) and the C-terminal transactivation domain (C-TAD) containing asparaginyl residue 803 (N). Degradation of HIF-1α under normoxic conditions is dependent on the oxygen-dependent degradation domain (ODDD). P: proline residues 402 and 564; NLS: nuclear localisation signal.
HIF-1α is rapidly degraded under normoxic conditions, when it is hydroxylated at two proline residues within the oxygen-dependent degradation domain by prolyl hydroxylase domain (PHD) proteins 1–3 (fig. 2⇓) 5. This post-translational modification serves as a recognition motif for subsequent ubiquitination by the E3 ubiquitin ligase activity of the von Hippel–Lindau tumour suppressor protein (pVHL), thus targeting HIF-1α for degradation by the proteasome. Hypoxic conditions inhibit prolyl hydroxylation, leading to stabilisation of HIF-1α 6, 7.

Hypoxia-inducible factor (HIF)-1-mediated oxygen sensing of transcription. The schematic illustrates how the α-subunit of the heterodimeric transcription factor HIF-1 conveys hypoxic conditions to transcriptional control. a) Under hypoxic conditions, HIF-1α dimerises with HIF-1β and binds to HIF-responsive target genes, such as erythropoietin or vascular endothelial growth factor. After recruitment of the transcriptional coactivator CREB (cAMP response element binding)-binding protein (CBP)/p300 to HIF-1α and additional binding of gene- or tissue-specific transcriptional cofactors, HIF-1-dependent transcription is induced. b) Under normoxic conditions, HIF-1α is absent because it is inactivated and immediately degraded by the proteasomal machinery. The oxygen sensors translating the oxygen concentration into changes in HIF-1α are oxygen-dependent hydroxylases: factor inhibiting HIF-1 (FIH-1) and prolyl hydroxylase domain-containing proteins 1–3 (PHDs). In the presence of oxygen, FIH-1 hydroxylates asparaginyl residue 803 (N) within the C-terminal transactivation domain of HIF-1α, preventing the recruitment of CBP/p300 to the HIF-1 complex, which leads to reduction of transcriptional activity. PHDs hydroxylate two prolyl (P) residues (P402 and P564) within the oxygen-dependent degradation domain of HIF-1α, allowing binding of the von Hippel–Lindau tumour suppressor protein (pVHL). pVHL functions as a recognition element for the multiprotein E3 ubiquitin ligase complex, which targets HIF-1α, via polyubiquitination, for proteasomal degradation. Whereas dimerisation and transactivation of the HIF-1 complex takes place in the cell nuclei, it is still under discussion whether the oxygen sensing process of FIH-1/PHDs, the binding of pVHL and the proteasomal degradation can take place in both cytoplasm and nucleus. OH: hydroxyl group.
Stabilised HIF-1α translocates into the nucleus, where it heterodimerises with HIF-1β (fig. 2⇑). The HIF-1 complex binds to DNA regulatory sequences called hypoxia response elements (HREs), which are present in the promoter or enhancer regions of HIF-1 target genes 8. After recruiting transcriptional coactivators, such as CREB (cAMP response element binding)-binding protein (CBP)/p300, the target gene expression is induced 9. The interaction of HIF-1α with CBP/p300 is also oxygen controlled via hydroxylation (fig. 2⇑). An asparagyl-hydroxylase called factor inhibiting HIF-1 modifies the C-terminal transactivation domain (C-TAD) of HIF-1α and inhibits its association with CBP/p300 when oxygen is available 10. Thus, hypoxia, apart from stabilising HIF-1α, can also lead to activation of transcriptional activity. Furthermore, movement of HIF-1α between cell compartments appears to be dependent on the oxygen concentration 11. The present review focuses on the contributions of optical methods to the improvement of the understanding of oxygen-dependent regulation of the HIF-1 pathway. The tissue- and cell-specific distribution of the oxygen-controlled HIF-α subunits was visualised by immunohistochemistry and immunofluorescence. Transcriptional activity of HIF-1 was monitored using green fluorescent protein (GFP) as a reporter under control of HREs in living cells, spheroids and living mice. Using the cyan and yellow variants (CFP and YFP, respectively) of GFP to generate fusion proteins with HIF subunits and regulatory proteins, the subcellular distribution, migration and interaction were imaged in vivo by means of fluorescence recovery after photo-bleaching (FRAP) and fluorescence resonance energy transfer (FRET). Noninvasive imaging of these cellular and molecular processes by two-photon laser microscopy complements ex vivo molecular biology assays and provides an additional spatial and temporal dimension to the understanding of the cellular hypoxic response.
TISSUE- AND CELL-SPECIFIC EXPRESSION OF THE HIF-α SUBUNITS
In order to better understand the mechanisms and sites of HIF-α regulation, the distribution patterns of HIF-α in normal tissue, and changes that occurred during physiological development, malignant transformation or under ischaemia have been examined by immunohistochemistry.
The spatial and temporal distribution of HIF-1α in vivo has been determined in healthy mice in response to a reduced inspired oxygen partial pressure. Immunohistochemical examination of brain, kidney, liver, heart and skeletal muscle has revealed that HIF-1α protein was present in the nucleus, even under normoxic conditions. Nuclear and cytoplasmic staining of HIF-1α has been found to increase in response to systemic hypoxia (6% oxygen), except in the lung 12. These findings suggest that physiological oxygen concentrations in normal tissue, ranging 2–5% oxygen 13, allow levels of HIF-1α that may help to maintain homeostasis of cellular activities, e.g. by controlling steady-state expression of genes required to provide cellular energy.
In addition to hypoxia, other signals generated during cellular differentiation and in response to growth factors may regulate HIF-1α expression in normoxic organs within distinct cell types. Strong accumulation of HIF-1α protein was detected in specific cells, such as in neurons of the hippocampus, the dentate gyrus in the brain and in hepatocytes in the liver 12. In contrast, HIF-1α was undetectable in mouse pulmonary cells even under hypoxia (6% oxygen) 12. However, HIF-1α expression was induced when ferret lungs were ventilated with <1% oxygen for 4 h. Upon re-oxygenation, HIF-1α was rapidly degraded, with a half-life of <1 min 14. Within the hypoxic ferret lung, HIF-1α was expressed in most cell types, including smooth muscle, alveolar cells and vascular epithelium 14.
Alternatively, cells that do not contain HIF-1α may preferentially express HIF-2α to adapt to hypoxia. Although HIF-2α was not detectable under normoxic conditions, systemic normobaric hypoxia (8% oxygen) and anaemic hypoxia (0.1% carbon monoxide) induced HIF-2α in all organs of rats investigated, including brain, heart, lung, kidney, liver, pancreas and intestine 15. The time course and amplitude of induction varied between organs, apart from the lung, where HIF-2α protein was only very weakly increased.
Immunohistochemistry revealed nuclear accumulation in distinct cell populations in each tissue, which were always nonparenchymal in some organs (kidney, pancreas and brain), predominantly parenchymal in others (liver and intestine) or equally distributed (myocardium) 15. In the lung, however, immunohistochemistry on tissue from rats exposed to 8% oxygen showed strong and frequent nuclear staining of type II pneumocytes, while anaemic hypoxia only weakly increased HIF-2α abundance. In contrast, the endothelial cells of pulmonary arteries showed strong nuclear signals in response to both forms of hypoxia 15. Thus, HIF-2α may play an important role in the transcriptional response to hypoxia in vivo that is complementary, rather than redundant, to HIF-1α.
The expression patterns and the extent to which HIF-1α and -2α exert similar or distinct functions in vivo have been investigated by comparative immunohistochemistry of different cell types from kidneys and heart in response to hypoxia or ischaemia 16–18. To study the expression of HIF-1α versus -2α in kidneys, rats were exposed to systemic hypoxia, anaemia induced by bleeding, functional anaemia (0.1% carbon monoxide), renal ischaemia or cobalt(II) chloride (CoCl2; known to mimic hypoxia). These treatments led to marked nuclear accumulation of HIF-1α and -2α that was selective with respect to cell type, kidney zone and experimental conditions 16. HIF-1α was mainly induced in tubular cells, whereas HIF-2α was expressed in endothelial cells of glomeruli and in peritubular endothelial cells and fibroblasts. Despite the well-known heterogeneity of renal oxygen tensions (<0.1–7% oxygen), no signals from HIF-α units were observed in the kidneys of normal rats 16. In contrast, HIF-α proteins were expressed in developing human and rat kidneys during glomerulogenesis and nephrogenesis. HIF-1α seems to be primarily involved in tubulogenesis and HIF-2α in renal vasculogenesis; both isoforms are found during glomerulogenesis 18. Similarly, during first trimester development of the human lung, which takes place in the relatively hypoxic environment of the uterus, HIF-1α was found to be mainly restricted to the branching epithelium, whereas HIF-2α was also present in the vascular structures of the lung parenchyma 19. The different spatial expression of HIF-α species may reflect their different roles in pulmonary development. HIF-1α appears to be more important for organogenesis, whereas the function of HIF-2α seems to be the fine-tuning of pulmonary vascularisation and remodelling 19. These data support the notion that HIF-1α and -2α already exert distinct functions during early development of organs, as well as their function in response to hypoxic or ischaemic conditions in adult tissues.
In normal human tissue very little expression of HIF-1α or -2α has been found. Significant HIF-2α expression was found to be limited to Kupffer cells in the liver and bone marrow macrophages. This HIF-2α staining was predominantly cytoplasmic, with a small amount of nuclear localisation in some macrophages 20. Weak HIF-1α expression was also found in foetal hepatocytes, proliferating B-cells in tonsils and spleen, and seminiferous tubules of the testis, and was correlated with cell proliferation 21.
In contrast, HIF-1α and -2α proteins were detected in most types of human tumours and their metastases, including glial, breast, ovarian, gastric, colon, hepatocellular, pancreatic, prostate, bladder and renal carcinomas 20–23. Within tumours, the extent, intensity, intracellular localisation and distribution of HIF-1α and -2α staining was heterogeneous 20, 21. HIF-1α staining was primarily nuclear, but some cytoplasmic staining of HIF-1α was also detected in colon, breast, pancreas and prostate adenocarcinomas. However, HIF-1α was not observed in tumour-adjacent stroma tissue 21. In renal cell carcinoma, staining for HIF-1α was cytoplasmic and nuclear, whereas HIF-2α was exclusively nuclear. In a hepatocellular carcinoma, nuclear and cytoplasmic staining for both proteins was observed 20. In the perinecrotic regions of tumours, the HIF-1α protein staining tended to be stronger than that of HIF-2α 20. HIF-1α-positive cells were also prominent at tumour margins and surrounding areas of neovascularisation 21. Variable amounts of nuclear expression of HIF-1α and -2α, or incompletely overlapping patterns of the two HIF-α subunits, suggest that hypoxia-independent mechanisms in the tumour micro-environment contribute to the upregulation of HIF-α proteins in cancers.
MONITORING TRANSCRIPTIONAL ACTIVITY OF HIF-1
Although immunohistochemical analysis can be useful in determining whether HIF-α proteins are present at higher levels in cancer cells compared with surrounding normal tissue, it does not reveal whether the HIF-1 complex is transcriptionally active. To investigate the transcriptional activity of HIF-1 in vivo, several HREs are usually fused to GFP or its variants. In response to hypoxia or CoCl2, induced HIF-1-dependent GFP expression was observed in several cell lines both by flow cytometry analysis and fluorescence microscopy 24–26.
To monitor and assess the level of endogenous HIF-1 activity during tumour development and growth, Serganova et al. 24 have developed a retroviral vector for stable transduction of tumour cells, bearing a dual reporter gene cassette with an HIF-1-inducible 8HRE-thymidine kinase/GFP fusion and a constitutively expressed reporter gene, Discosoma red fluorescent protein 2/xanthine phosphoribosyltransferase. This dual reporter system allowed imaging of HIF-activity in spheroids or in xenografts in living mice, both by confocal laser microscopy as well as by noninvasive in vivo positron emission tomography (PET) of a radiolabelled probe 27. In multicellular spheroids grown from transduced C6 rat glioma cells, HIF-1 activity was detectable within the central region when spheroids exceeded 350 µm in diameter and had a hypoxic core zone. These findings are in agreement with those for immunostained multicellular prostate tumour spheroids (350–400 µm in diameter) analysed by quantitative laser scanning microscopy, which showed an increase in HIF-1α expression in the more central parts, correlating with reduced pericellular oxygen levels as measured by microelectrodes 28. In contrast, Indovina et al. 26 reported that three-dimensional cell organisation leads to almost immediate HIF-1 activation even in small spheroids (100 µm in diameter) grown from osteosarcoma cells stably transfected with an HRE-enhanced (E)GFP reporter vector, when analysed in vivo with two-photon laser microscopy. Since 100 µm is still within the diffusion limit for oxygen, the observed HIF activity in small spheroids was most likely not due to hypoxia alone, but may have been caused by paracrine factors produced during early three-dimensional cell growth 26.
In addition, HIF-1 activity has been shown to be heterogeneous and dependent on tumour size in xenografts transfected with HIF-1-specific GFP reporter vectors. With increased diameter (>3 mm) of C6 rat glioma tumours, a marked increase in HIF-1 transcriptional activity was observed in the core regions of tumours, as visualised by micro-PET and confirmed by fluorescence microscopy of excised tumours. Ischaemia–reperfusion injury in small tumour xenografts caused a rapid induction of HIF-1 transcriptional activity that persisted for longer (24 h) than in normal tissues, revealing that, besides hypoxia, oncogenic signalling may play a role in controlling HIF-1 activity 24. In order to monitor HIF-1 activity in real time in tumour xenografts, Liu et al. 25 chose a 5HRE promoter fused to destabilised EGFP with a half-life reduced to <2 h (d2EGFP). HIF-1 activity, monitored by EGFP fluorescence intensity, correlated with hypoxic treatment, and re-oxygenation resulted in a rapid decrease in HIF-1 activity. Within solid tumours of living mice, HIF-1 activity was heterogeneous, and immunohistochemical analysis confirmed that d2EGFP-expressing regions reflecting HIF-1 activity were almost identical to areas staining for the hypoxia marker pimonidazole 25. On closer inspection, however, d2EGFP-positive cells appeared to be more abundant than pimonidazole-stained cells. This might either reflect a lower threshold for HIF-1 activation compared with the formation of pimonidazole adducts or be due to nonhypoxic activation of HIF-1 that does not increase pimonidazole staining 25.
SUBCELLULAR DISTRIBUTION OF HIF SUBUNITS AND INTERACTION WITH REGULATORY FACTORS
It has been widely shown via immunofluorescence that HIF-1α accumulates in nuclei of many cell lines in response to hypoxia, hypoxia-mimicking agents, inflammatory cytokines, nitric oxide and hormones 29–34. The nuclear accumulation of HIF-1α is a consequence of either reduced degradation or enhanced synthesis of HIF-1α. Using GFP fusion protein constructs and deletion variants, Kallio et al. 11 identified a nuclear localisation signal motif in the C-terminus of HIF-1α that mediated the translocation into the nucleus. Nuclear translocation itself seems to be constitutive, independent of oxygen availability or pVHL status in cells infected with an adenovirus expressing GFP fused to the HIF-1α subunit 35. During re-oxygenation, HIF-1α is relocated to the cytoplasm in order to be degraded by the proteasome in many cell lines 11, 35, 36. Export of HIF-1α upon re-oxygenation is not mediated by chromosome maintenance region (CRM)1/Exportin1, since the nuclear export was unaffected by the drug leptomycin B 35 and a typical nuclear export signal has not been identified within HIF-1α. However, Groulx and Lee 35 provided evidence that pVHL plays a dual role, being involved in both degradation of HIF-1α and export to the cytoplasm. Groulx and Lee 35 showed that interaction of HIF-1α and pVHL occurs in the nucleus, where HIF-1α is ubiquitinated before it is exported through a non-CRM1-dependent mechanism to the cytoplasm. According to this model, HIF-1α becomes completely located in the nucleus under hypoxia, because it does not interact with pVHL and, therefore, cannot be exported to the cytoplasm 35.
However, it has been previously reported that trapping of HIF-1α either in the nucleus or in the cytoplasm does not prevent HIF-1α degradation 37, implying that HIF-1α degradation can occur in both compartments. Indeed, proteasome complexes and pVHL are present in both the cytoplasm and nucleus 38–40. Furthermore, localisation studies have shown that the oxygen sensors initiating HIF-1α degradation, namely the PHD proteins, are also found in both compartments 41–43. Consistent with these findings, Zheng et al. 44 recently showed that compartmentalisation of HIF-1α degradation and subcellular distribution are regulated in a cell type-specific manner. Fragments of the N-terminal transactivation domain (N-TAD) of HIF-1α that were able to inhibit pVHL-mediated degradation when fused with GFP were targeted to different cell compartments, allowing identification of two possible cell types 44. In highly proliferating or transformed cells (e.g. HepG2 hepatoma cells), HIF-1α degradation occurs mainly in the cytoplasm. Under hypoxia, HIF-1α translocates to the nucleus and, upon re-oxygenation, the protein needs to be relocated to the cytoplasm to be efficiently degraded. In this cell type, retention of HIF-1α in the nucleus can protect it from degradation. In contrast, in primary endothelial cells (e.g. from mouse brain), HIF-1α can be degraded with equal efficiency in the nucleus and cytoplasm. In this cell type, HIF-1α is present in both compartments under conditions of normoxia, hypoxia or re-oxygenation. Furthermore, Mylonis et al. 45 suggested that the activity level of the mitogen-activated protein kinase (MAPK) pathway contributes to the cell type-specific subcellular distribution of HIF-1α. Amino acid residues were identified within the inhibitory domain of HIF-1α that, when phosphorylated, enhanced nuclear accumulation of GFP-HIF-1α. Since MAPK pathway activity is highly connected to the proliferation state of the cell, the distribution of HIF-1α could be differentially regulated in highly transformed, rapidly proliferating cells and primary, slow-growing cells.
In contrast to HIF-1α, the β-subunit of the HIF-1 complex is found exclusively in the nuclear compartment and is unaffected by oxygen concentration. Furthermore, HIF-1α accumulation and nuclear translocation occur independently of HIF-1β, as shown by immunofluorescence in HIF-1β-mutant mouse hepatoma cells and embryonic stem cells 36. However, HIF-1β has been shown to be indispensable for HIF-1 DNA binding and transactivation 46.
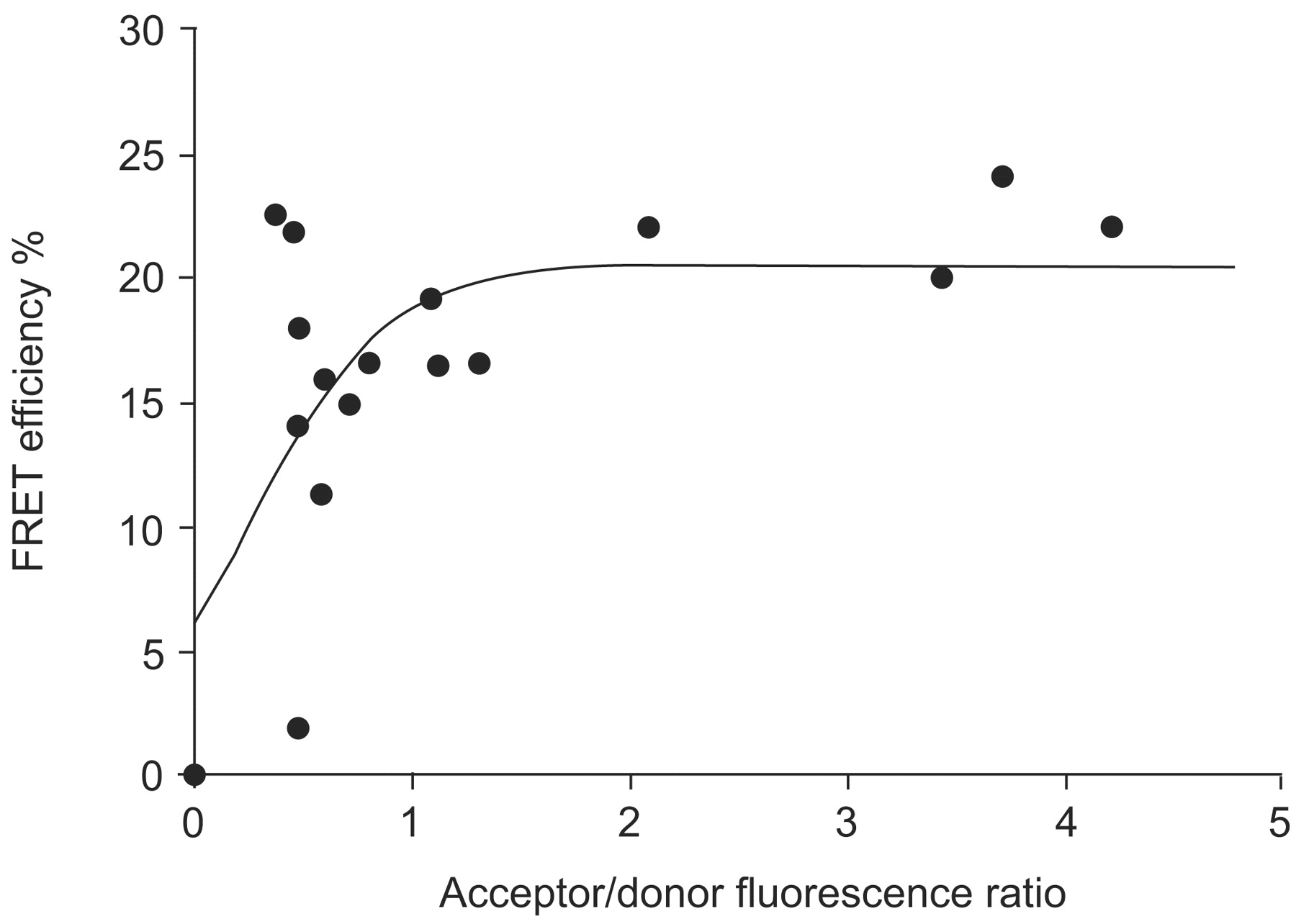
Within the hypoxic nucleus, endogenous HIF-1α and HIF-1β are colocalised in speckle-like structures, as observed by immunofluorescence combined with three-dimensional visualisation studies 47. These speckle-like structures were also observed when ECFP-HIF-1α and EYFP-HIF-1β fusion proteins were coexpressed, but not when each construct was transfected alone, implying that these structures are the sites of HIF-1 complexes 48. Using FRAP, it has been observed that EYFP-HIF-1β moves faster than ECFP-HIF-1α (50% recovery of fluorescence after 31 s compared with 65 s, respectively) when cells were cotransfected with both fusion proteins 48. This indicates that the subunits can move separately and, therefore, do not always form HIF-1 complexes inside the nucleus. To analyse the interaction of HIF-1α and HIF-1β in the DNA-bound HIF-1 complex in living cells, ECFP or EYFP was fused to the N-terminus or C-terminus of the full-length HIF-1 subunits and subjected to FRET analysis. The highest FRET efficiency and, thus, closest association of the fusion proteins was observed when ECFP and EYFP were located at the N-terminus of the HIF-1α and HIF-1β subunits, respectively (fig. 3⇓). The fluorophore-labelled N-termini of the HIF-1 subunits were colocalised as close as 6.2 nm and the C-termini as close as 6.7 nm, and even the N- and C-termini of the HIF-1 complex were not further apart than 7.4 nm 48, indicating that the HIF-1 complex is more compact than previously thought. Furthermore, the subnuclear distribution of HIF-1α/β heterodimerisation was heterogeneous when imaged by sensitised FRET (fig. 4⇓). Different FRET efficiencies within the cell nucleus may result from local variation in the mean distance between the ECFP- and EYFP-labelled HIF-1 subunits. Since experimental X-ray crystallography data on the three-dimensional structure of the HIF-1 complex are not yet available, due to the low solubility of the subunits 49, FRET experiments are the method of choice to gain more insights into the three-dimensional organisation of the HIF-1 complex and its nuclear distribution in living cells under different oxygen concentrations.

In vivo analysis of hypoxia-inducible factor (HIF)-1α and HIF-1β dimerisation using fluorescence resonance energy transfer (FRET). Enhanced cyan fluorescent protein (ECFP)-HIF-1α and enhanced yellow fluorescent protein (EYFP)-HIF-1β were coexpressed in U2OS osteosarcoma cell nuclei and imaged by two-photon laser scanning microscopy, while the cells were kept under hypoxia. According to the theory of FRET, energy from the donor molecule (ECFP-HIF-1α) is transferred to the acceptor (EYFP-HIF-1β) if the two molecules come closer than 10 nm. FRET efficiencies from cell nuclei were calculated using the acceptor bleaching method. The mean FRET efficiency of ECFP-HIF-1α and EYFP-HIF-1β interaction depends on their relative expression within the respective cell nucleus. Therefore, the distance between the molecules was determined (6.2 nm) when a plateau was reached 48.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Subnuclear distribution of hypoxia-inducible factor (HIF)-1α and HIF-1β dimerisation, analysed by fluorescence resonance energy transfer (FRET). U2OS osteosarcoma cells were transfected with enhanced cyan fluorescent protein (ECFP)-HIF-1α and enhanced yellow fluorescent protein (EYFP)-HIF-1β. Under hypoxia, both fusion proteins were expressed in cell nuclei and FRET was imaged by laser scanning microscopy. The distribution of FRET efficiencies within a single cell nucleus was estimated using the sensitised FRET method 50 and visualised in false colour mode, as indicated by the colour bar. Subnuclear variations in FRET efficiency most likely reflect local variation in the mean distance between the ECFP- and EYFP-labelled HIF-1 subunits. Scale bar = 2 μm.
In addition, HIF-1-mediated transcription requires the recruitment of coactivators, such as CBP/p300 and factors belonging to the steroid receptor coactivator (SRC)/p160 family, to build a transcriptionally active HIF-1/coactivator complex 9. Ruas et al. 51 showed that colocalisation of CFP-mouse-HIF-1α and YFP-CBP in nuclear accumulation foci was dependent on the integrity of the C- and N-TADs of HIF-1α, which correlated with HIF-1-mediated activation of transcription. Further colocalisation studies of CFP or YFP fusion proteins of HIF-1α, HIF-1β, CBP and SRC-1 suggest that CBP, besides interacting with HIF-1α, can also interact with HIF-1β, thereby stabilising the formation of the HIF-1α/β heterodimer 52. In addition, CBP was found to mediate the interaction between SRC-1 and HIF-1α, indicating that CBP is the mediator of HIF-1/coactivator formation 9.
To activate transcription in response to hypoxia in a tissue-specific manner, further cofactors have to be recruited to HIF-1 or the HIF-1/coactivator complex, e.g. hepatocyte nuclear factor-4 to activate transcription of erythropoietin in the liver 53. The challenge for the near future will be to analyse the dynamics and architecture of the assembly of the HIF-1/coactivator/cofactor multiprotein complexes in vivo, by combining FRAP and FRET methods.
CONCLUSIONS
Immunohistology and immunofluorescence revealed that the oxygen-labile α-subunits of HIF-1 and -2 are hardly detectable in normal tissues of human adults, although the oxygen concentration within the tissue ranges 3–5%. However, accumulation of HIF-α is observed during early development of organs, in response to ischaemia and in tumour tissue. HIF-1α and -2α staining is heterogeneous and only partially overlapping, suggesting that the α-subunits are not redundant and play different roles. By expressing GFP under control of an HRE in tumour cells, multicellular spheroids or xenographs of living mice, HIF-1 transcriptional activity has been shown to be increased in response to hypoxia or ischaemia. Under hypoxic conditions, HIF-1α is shown to be localised in the nucleus of most cells, although in other cells HIF-1α is also found in the cytoplasm. By expressing different GFP fusion proteins of HIF-1α in living cells, the subcellular localisation was found to be cell specific and regulated by the degradation pathway of HIF-1α.
Within the nucleus, HIF-1α and HIF-1β were localised in speckle-like structures, where interaction of the subunits takes place. Heterodimerisation of ECFP-HIF-1α und EYFP-HIF-1β was analysed in living cells using FRAP and FRET methods. Distances between HIF-1α and -1β fusion proteins were calculated as 6.2–7.4 nm. Furthermore, HIF-1-mediated transcription requires the recruitment of several transcriptional coactivators and tissue-specific cofactors to the HIF-1 complex.
The challenge for the near future will be to analyse the dynamics and architecture of the assembly of the multiprotein complexes involved in the transduction of the hypoxic signal in living cells, by combining fluorescence recovery after photo-bleaching and fluorescence resonance energy transfer with noninvasive methods such as two-photon laser microscopy.
Support statement
This work received financial support from the Deutsche Forschungsgemeinschaft (grants FA225/18-2 and FA225/19-2) and from the European Commission under FP6 (contract no. LSHM-CT-2005-018725, PULMOTENSION). J. Fandrey received travel reimbursement from the European Respiratory Society for attending the Fifth Lung Science Conference in Taormina, Italy.
Statement of interest
A statement of interest for J. Fandrey can be found at www.erj.ersjournals.com/misc/statements.shtml
Footnotes
-
Previous articles in this series: No. 1: Wagner PD. The biology of oxygen. Eur Respir J 2008; 31: 887–890. No. 2: Zhou G, Dada LA, Sznajder JI. Regulation of alveolar epithelial function by hypoxia. Eur Respir J 2008; 31: 1107–1113.
- Received January 28, 2008.
- Accepted January 31, 2008.
- © ERS Journals Ltd
References










