





Abstract
Although children commonly travel to high altitudes, their respiratory adaptation to hypoxia remains elusive. Therefore, in the present study respiratory inductive plethysmography, pulse oximetry (Sp,O2) and end-tidal CO2 tension (PET,CO2) were recorded in 20 pre-pubertal children (aged 9–12 yrs) and their fathers during 1 night in Zurich (490 m) and 2 nights at the Swiss Jungfrau-Joch research station (3,450 m) following ascent by train within <3 h.
In children, mean±sd nocturnal Sp,O2 fell from 98±1% at 490 m to 85±4 and 86±4% at 3,450 m (nights 1 and 2, respectively); PET,CO2decreased significantly from 37±6 to 32±3 and 33±4 mmHg (3,450 versus 490 m). In adults, changes in nocturnal Sp,O2 and PET,CO2at 3,450 m were similar to those in children.
Children spent less time in periodic breathing at 3,450 m during night 1 and 2 (8±11 and 9±13%, respectively) than adults (34±24 and 22±17%, respectively), and their apnoea threshold for CO2 was lower compared with adults (27±2 and 30±2 mmHg, respectively, both nights). Sp,O2, PET,CO2 and time in periodic breathing at altitude were not correlated between children and their fathers.
In conclusion, children revealed a similarly reduced nocturnal O2 saturation and associated hyperventilation at high altitude as adults but their breathing pattern was more stable, possibly related to a lower apnoea threshold for CO2.
Many families with children travel to high mountain areas for leisure activities, to pursue a parental occupation and for other reasons 1. However, little is known about children's respiratory adaptation to hypobaric hypoxia and their tolerance of high altitude. In adults, hypobaric hypoxia increases ventilation and induces respiratory alkalosis 2–4. This attenuates hypoxaemia but may also destabilise ventilation due to hypocapnia that triggers apnoea 5. Periodic breathing characterised by crescendo–decrescendo ventilation with hyperpnoea alternating with hypopnoea/apnoea is typically observed in adults at high altitude. Periodic breathing causes frequent arousals from sleep 6, 7 resulting in poor sleep quality, one of the manifestations of acute mountain sickness 8, 9. There is a considerable inter-subject variability in periodic breathing due to individual differences in autonomic control of ventilation 10. According to feedback control theory, susceptibility to periodic breathing depends on various factors, such as on controller gain, gas exchange, the circulation time and response lags involved in the chemoreflex feedback 11. Consistent with this theory, periodic breathing has been induced in animals by inserting a circulatory delay system between the lungs and the brain 12 or by augmenting controller gain by an increase in the gain of a servo-respirator during mechanical ventilation 13. Furthermore, ventilatory instability and periodic breathing have been shown to depend on the proximity of the eupnoeic carbon dioxide tension (PCO2) to the apnoeic and hypopnoeic threshold PCO2, as well as on the ventilatory sensitivity to CO2 below eupnoea 14. These factors may differ between children and adults and may affect ventilation, oxygenation and thereby well-being and performance at high altitude 15–17. As respiratory adaptation to acute altitude exposure has not been studied in detail in children, the current authors performed unobtrusive breathing pattern recordings by calibrated respiratory inductive plethysmography and actimetry in pre-pubertal children and their fathers. Studies were performed at low altitude in Zurich (490 m) and during 2 consecutive nights at the Jungfrau-Joch high-altitude research station (3,450 m), Switzerland. Some of the results of these studies have been reported previously as a case report 18.
METHODS
Subjects
In total, 20 pre-pubertal children (four young females) at Tanner stage I 19, 20, i.e. without clinical signs of puberty, aged mean±sd (range) 11±1 (9–12) yrs, and with a body mass index of (BMI) 16.2±2.0 kg·m−2, were recruited from lowland residencies (<1,000 m) in Switzerland. In addition, 20 males (19 were biological fathers of participating children) aged 44±4 (36–57) yrs, and with a BMI of 23±1.6 kg·m−2 were recruited from lowland residencies (<1,000 m) in Switzerland. Participants were healthy, did not suffer from any sleep or breathing disorder, and had not stayed at altitudes >2,000 m in the 2 months prior to the study. No drugs other than paracetamol were allowed to treat high altitude-related headaches. All participants gave informed consent and the Institutional Ethics Committee of the University of Zurich (Zurich, Switzerland) approved the study.
Measurements
A medical history and physical examination were obtained. Acute mountain sickness (AMS) was assessed using the environmental symptoms questionnaire (ESQ); a cerebral score with ≥0.7 points was considered to indicate AMS 21. Participants were assisted in filling out the questionnaires by an investigator.
Nocturnal polygraphic studies included: calibrated respiratory inductive plethysmography (SomnostarPT; Sensormedics, Yorba Linda, CA, USA), pulse oximetry, ECG, and capnography of expired air to estimate end-tidal CO2 tension (PET,CO2) as a surrogate of arterial PCO2 (Pa,CO2; CapnoSleep; Weinmann, Hamburg, Germany). The technical pulse oximeter response time from sensor disconnection to a drop in O2 saturation was 8.5 s. The capnograph response time from the beginning of the rise in CO2 concentration to 90% of maximal amplitude after switching from room air to a 5% CO2 mixture was 0.6 s. The respiratory inductive plethysmograph was first calibrated using the Qualitative Diagnostic Calibration method 22, and then against the integrated output of a flow meter during 10–20 breaths 23. Validation of the calibration in the morning after sleep studies revealed a deviation of tidal volume (VT) by inductance plethysmography from corresponding flow meter values of ≤20% in all instances. Wrist actigraphy was performed as an indirect measure of sleep and wakefulness (Actiwatch; Cambridge Neurotechnology, Cambridge, UK) 8, 24.
Protocol
Father and child pairs underwent baseline examinations in Zurich (490 m). They ascended to the Jungfrau-Joch research station (3,450 m) 2–4 weeks later by a <3-h train ride and stayed there for 48 h. Nocturnal polygraphic recordings were performed during 1 night at 490 m and during the 2 nights at 3,450 m. The nocturnal rest period lasted ∼8 h, from 22:00–06:00. Clinical examination and assessment of AMS were performed in the evenings and mornings.
Data analysis and statistics
Polygraphic recordings were analysed from lights off in the evening to lights on in the morning. This period was defined as time in bed (TIB). Breathing pattern variables derived from respiratory inductive plethysmography were measured breath-by-breath and mean values were computed for TIB 23. Periodic breathing was defined as a pattern of waxing and waning of ventilation with periods of hyperventilation alternating with central apnoeas/hypopnoeas for at least three successive cycles, with hypopnoeas showing a >50% amplitude reduction of the inductive plethysmographic VT signal compared with the preceding 2-min baseline of stable breathing for >5 s (fig. 1⇓) 5, 8, 25. The number of periodic breathing cycles, their length (the cycle time), and the fraction of TIB spent with periodic breathing were determined. Occasional apnoeas/hypopnoeas that were not part of periodic breathing were defined as a reduction of the inductive plethysmographic VT signal to <50% of the preceding 2 min baseline of stable breathing during ≥10 s in adults and during ≥5 s in children 26, 27. Obstructive apnoeas/hypopnoeas were differentiated from central events by ribcage-abdominal asynchrony 23 and by continued chest wall excursions in the absence of PET,CO2 deflections. The apnoea/hypopnoea index (AHI) was defined as the number of apnoeas/hypopnoeas (including periodic breathing cycles) per hour TIB. O2 desaturations ≥4% per hour of TIB were determined as the O2 desaturation index. Sighs were defined as large breaths with VTs greater than twice the previous stable VT amplitude 28. The CO2 apnoea threshold was defined as the value of PET,CO2 of the last breath before an apnoea occurred 29. The CO2 reserve (ΔPET,CO2) was calculated as the difference in PET,CO2during eupnoea and the CO2 apnoea threshold. The lung-to-finger circulation time was measured as the time from the end of an apnoea to the subsequent nadir of arterial O2 saturation as an estimate of the circulatory delay from the lungs to the peripheral chemoreceptors 29. To evaluate whether there were differences in circulation time between children and adults independent of body size, the circulation time was also expressed as time in seconds divided by height. Oximeter probes were placed on the finger rather than on the ear lobe as this was found to be less disturbing to the children. Mean CO2 apnoea threshold and circulation time from three consecutive apnoea/hyperpnoea cycles were measured every 15 min if applicable. Activity and rest periods were derived from actigraphy with dedicated software. The acceleration level below which sleep is assumed was graphically assigned 24. Time with acceleration below threshold was defined as rest time, and estimated sleep efficiency as rest time in per cent of TIB.
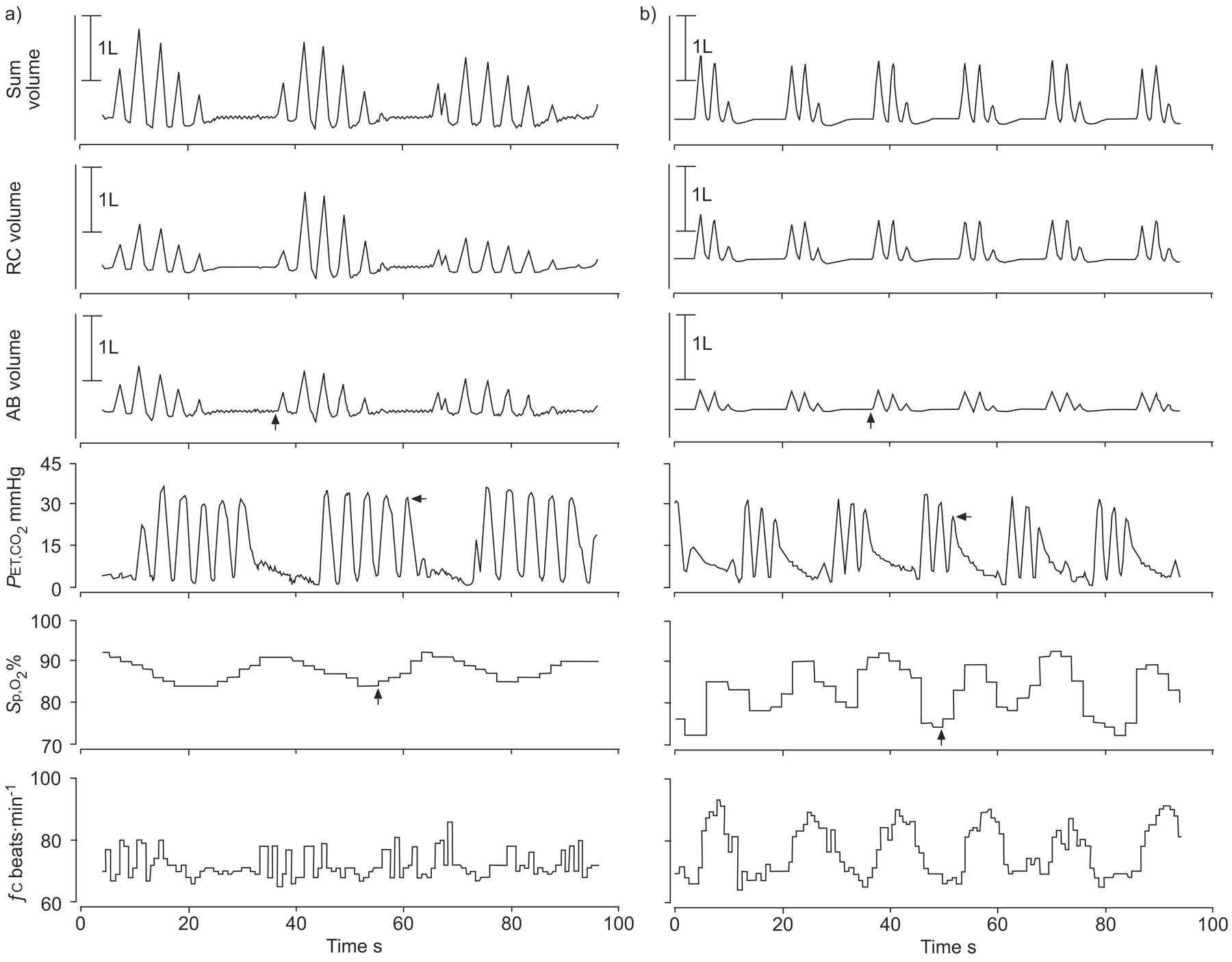
Nocturnal polygraphic recording obtained at 3,450 m in a) a 43-yr-old father and b) his 11-yr-old son. Inductive plethysmographic signals reflecting the volume of the ribcage (RC), the abdomen (AB) and their sum, the lung volume. The end-tidal carbon dioxide tension (PET,CO2), arterial oxygen saturation measured by pulse oximetry (Sp,O2) and cardiac frequency (fc) measured by ECG are also shown. There is a periodic breathing pattern associated with fluctuations in Sp,O2and fc. This was extraordinarily pronounced in this child. The apnoea threshold for carbon dioxide is lower in the boy (b; 25 mmHg, horizontal arrow) than in his father (a; 30 mmHg, horizontal arrow). The circulation time measured as the time from the end of an apnoea to the corresponding nadir in arterial oxygen saturation (vertical arrows) is shorter in the boy (b; 12 s,) than for the father (a; 19 s).
Results are expressed as mean±sd. Measurements at different altitudes and between children and fathers were compared by ANOVA followed by Newman–Keuls tests if appropriate. A probability of p<0.05 was assumed to be significant.
RESULTS
In total, 17 children and 19 of the adults completed the entire protocol. One child returned home with his father after the first night at 3,450 m because of homesickness; one child was withdrawn from the study as she developed severe AMS 18; and in one child, the cardiorespiratory sleep study on the second night at 3,450 m could not be analysed because of technical failure.
Results of polygraphic studies are summarised in table 1⇓. In children and adults mean nocturnal O2 saturation was significantly reduced by 11% to 13% during the first and second night at 3,450 m when compared to baseline at 490 m. This was associated with a significant increase in minute ventilation (V′E) that was similar in children and adults when expressed in per cent of the corresponding baseline value at 490 m, although absolute values in L·min−1 were higher in adults. Consistent with an increased respiratory centre drive that induced hyperventilation, mean inspiratory flow (VT/inspiratory time) at 3,450 m was increased by 22% to 45% above baseline values at 490 m, while PET,CO2 was decreased. These changes were similar in children and adults (table 1⇓). Children achieved their increased V′E mainly by increasing the breath rate, whereas adults increased both the breath rate and the VT. Compared with adults, cardiac frequency (fC) in children was already higher at baseline and increased even more at high altitude. Actigraphical recordings revealed that rest time and estimated sleep efficiency decreased significantly in the first night at high altitude in children and adults (table 1⇓).
Nocturnal polygraphic recordings
The analysis of transient respiratory events is summarised in table 2⇓. In children and adults the observed apnoeas and hypopnoeas were nearly exclusively of the central type (>99%) and occurred as part of a periodic breathing pattern. Therefore, central and obstructive apnoeas/hypopnoeas are not reported separately. At 490 m, apnoeas/hypopnoeas were rarely detected. In contrast, at 3,450 m, periodic breathing with central apnoeas/hypopnoeas was common in children and adults. However, children had less apnoea/hypopnoea at 3,450 m. During the first night they spent less than a quarter of the time with periodic breathing compared with adults (table 2⇓, fig. 2⇓). On the second night at 3,450 m, adults had a slight reduction in the AHI compared with the first night but their AHI was still higher than that of children who spent less than half of the time in periodic breathing compared with adults. Children had less cyclical O2 desaturations than adults, but their swings in O2 saturation tended to be more pronounced than in adults, although these differences were not statistically significant (table 2⇓).

The percentage of the night-time spent with periodic breathing was classified into three groups: <10%, 10–30%, >30%. Most of the children (□) spent <10% of the night-time with periodic breathing. In contrast, almost all adults (▓) spent >10% of the night-time in periodic breathing.
Nocturnal periodic breathing and transient respiratory events
To evaluate potential mechanisms responsible for the greater stability of ventilation in children, their apnoea threshold for CO2 was compared with that of adults (figs 1⇑ and 3⇓). This analysis revealed a lower apnoea threshold in children (table 2⇑). Since the mean PET,CO2was similar in children and adults, this suggested a greater CO2 reserve in children during both nights at 3,540 m. Moreover, children had a significantly shorter lung-to-finger circulation time than adults (table 2⇑), and the cycle time in children was also shorter than the corresponding value in adults.

Time series of inductive plethysmographic recordings obtained at 3,450 m reflecting the volume of the ribcage (RC), abdominal volume (AB), and their sum, the lung volume, along with end-tidal carbon dioxide tension (PET,CO2), arterial oxygen saturation measured by pulse oximetry (Sp,O2) and cardiac frequency (fC). a) A short sequence of periodic breathing from a child begins with a sigh as the initial disturbance. The breathing pattern normalises after only four cycles of periodic breathing consistent with a stable respiratory control system. b) A recording obtained in an adult reveals irregular, large breaths that trigger periodic breathing with progressively increasing fluctuations in amplitude, suggesting instability of the respiratory control system.
Figure 4⇓ further illustrates factors responsible for the greater stability of ventilation in children compared with adults. The relationship between alveolar ventilation (V′A) and alveolar PCO2 (PA,CO2), i.e. the isometabolic line, is defined as 30:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Diagrammatic representation of the relationship between alveolar ventilation (V′A) and alveolar CO2 tension (PA,CO2) showing the factors responsible for the greater ventilatory stability in children compared with adults. The line connecting the eupnoeic PA,CO2 in children with the apnoeic threshold (▵: – – – –) is less sleep than in adults (•: – – – –), and the increase in V′A required to drive PA,CO2 below the apnoeic threshold (arrows) is greater in children (19%) than in adults (7%). The apnoeic threshold is marked at V′A = 0. #: eupnoeic level of PA,CO2.
V′A = K×V′CO2/ PA,CO2 (1)
where K is the constant and V′CO2 is the CO2 production. V′A is expressed in per cent of the value during eupnoea since its absolute value is not known and varies between children and adults. Although V′CO2 and the constant K have different values for children and adults, this has no effect on the metabolic line if V′A is expressed in per cent V′A at eupnoea and K×V′CO2 remains constant within individuals during stable resting conditions. During the first night at 3,540 m, the eupnoeic PET,CO2 (the surrogate for PA,CO2) at 100% V′A was 32 mmHg in both children and adults (table 1⇑). The apnoeic threshold PET,CO2 was 27 and 30 mmHg in children and adults, respectively (table 2⇑). The lines connecting the eupnoeic PA,CO2at 100% V′A on the metabolic line with the apnoeic threshold PA,CO2at V′A = 0 reflect the ventilatory responsiveness for PCO2 below eupnoea (ΔV′A/ΔPA,CO2 = 100% V′A eupnoea/CO2 reserve). The mean value of ΔV′A/ΔPA,CO2 was 2.5 times greater in adults than in children (50 and 20% V′A eupnoea per mmHg PA,CO2, respectively; fig. 4⇑). The increase in V′A required to drive PA,CO2from the eupnoeic level to the apnoeic threshold is much smaller in adults than in children (7 and 19%, respectively), which contributes to ventilatory instability in adults.
No symptoms or signs of high-altitude pulmonary oedema, ataxia or impaired consciousness that would have suggested impending high-altitude cerebral oedema in any of the participants (apart from the child excluded because of severe AMS) were found. Nevertheless, 10 (50%) out of the 20 children and six (30%) out of the 20 adults (p = 0.1) fulfilled the criteria for AMS according to the ESQ criteria (AMS-C ≥0.7) at least once during their stay at 3,450 m. AMS-C scores of children were not statistically different from the corresponding values in adults (table 3⇓). There was no significant correlation for both nights at high altitude between AMS-C scores and V′E (children: R = 0.24; adults: R = 0.03), O2 saturation (children: R = 0.05; adults: R = 0.14), PET,CO2(children: R = 0.06; adults: R = 0.13), and per cent time with periodic breathing (children: R = 0.14; adults: R = 0.08; p = nonsignificant (ns) for all analyses). Multiple regression analysis revealed no association of V′E, O2 saturation, PET,CO2 and per cent time with periodic breathing on the first and second night at 3,450 m with AMS-C scores in the following morning in children (R2 = 0.09, p = ns) or adults (R2 = 0.07, p = ns).
Symptoms of acute mountain sickness(AMS)
To evaluate a potential genetically determined component of ventilatory adaptation to hypoxia, changes in V′E, O2 saturation, PET,CO2and per cent time in periodic breathing from baseline at 490 m to the first and second night at 3,450 m were correlated among each of the 19 pairs of children and fathers. Multiple regression analysis did not reveal any statistically significant relationship of physiological variables within families (data not shown).
DISCUSSION
This is the first study to provide detailed data on nocturnal breathing pattern and ventilation in children acutely exposed to high altitude. Using calibrated respiratory inductive plethysmography and other unobtrusive monitoring techniques during 2 successive nights at 3,450 m, it was found that children had a similar degree of hypoxaemia and a proportional increase in mean ventilation at altitude as their fathers who accompanied them. In contrast, periodic breathing was much less pronounced in children than adults. The greater stability of breathing in children was related to a lower apnoea threshold for CO2, a larger CO2 reserve and a shorter circulation time compared with adults. Half of the children and 30% of the adults suffered from AMS during their stay at 3,450 m. Therefore, the greater stability of ventilation in children did not translate into a clinically superior tolerance of high altitude.
Acute exposure to hypobaric hypoxia at 3,450 m resulted in a similar degree of hypoxaemia in children and adults with a mean arterial O2 saturation of 84–86% (table 1⇑), which is comparable to the mean nocturnal O2 saturation of 86% previously observed in healthy adults at 3,605 m 8, but lower than the value of 91% recorded at 3,109 m in children aged 3–36 months 31. According to the expected ventilatory stimulation at altitude 2, V′E and mean inspiratory flow, which reflect respiratory centre drive 32, were proportionally increased by 22% to 42% in children and adults, while PET,CO2, the study surrogate of Pa,CO2, was decreased to 33 mmHg, indicating a similar degree of hypoxic-induced hyperventilation in children and adults (table 1⇑).
Although altitude exposure resulted in high counts of central apnoeas/hypopnoeas, which occurred as part of a periodic breathing pattern in children and adults, this was significantly less pronounced in children (table 2⇑). The shorter cycle length of periodic breathing additionally contributed to a much lower fraction of the night-time spent with periodic breathing, i.e. <10%, compared with 22–34% in adults, which is similar to the corresponding value of 25% previously observed in adults at 3,650 m 8. To evaluate potential mechanisms responsible for the greater stability of ventilation in children, the current authors applied the concepts described by Dempsey 30, Nakayama et al. 14 and Xie et al. 33, as illustrated in figure 4⇑. The present authors measured the apnoea threshold for CO2 and found it to be 3 mmHg lower in children than adults, i.e. 27 versus 30 mmHg in both nights at 3,450 m (table 2⇑, figs 1⇑ and 4⇑). As the mean nocturnal PET,CO2was not significantly different between children and adults, this indicated that the CO2 reserve, i.e. the difference between baseline PCO2 and the apnoea threshold, was more reduced in adults. This explains their greater susceptibility to ventilatory instability. These findings principally agree with those of Xie and co-workers 29, 33 who observed a reduced CO2 reserve during hypoxia compared with normoxia in healthy adults hyperventilated by pressure support ventilation until central apnoea occurred. Although the present estimation of the apnoea threshold and the CO2 reserve cannot be directly compared with previous studies that used face masks, mechanical ventilation and sedation 29, 33, the current authors believe that the present results, obtained in a more natural setting with unobtrusive techniques, reflect relevant physiological mechanisms. In adults, periodic breathing was further promoted by an only modestly increased heart rate at altitude associated with a long circulation time (table 2⇑), which additionally impaired a stable control of ventilation 11 and contributed to the longer hyperpnoea duration and cycle time of periodic breathing compared with children. These findings are similar to the differences among patients with central sleep apnoea associated with heart failure compared to those with normal cardiac function 34. Thus, a higher heart and breath rate and a shorter circulation time, even when corrected for height (table 2⇑), allows children a more dynamic cardio-respiratory adaptation to high altitude compared with adults, and in combination with a greater CO2 reserve, reduces their propensity to periodic breathing. These concepts are consistent with a greater response of heart and breath rate and ventilation during a hypoxic challenge, as well as with a lower apnoea threshold for CO2 in children compared with adults, as reported by Marcus et al. 15.
Children tended to have more pronounced swings in O2 saturation during periodic breathing than adults (9 versus 7–8%, table 2⇑), although these differences were not statistically significant. As the minimal O2 saturation was 80–82% (table 2⇑) and the mean saturation 84–85% (table 1⇑), this indicated that the O2 saturation oscillated around the mean value, dropping from baseline during desaturations and overshooting during resaturations (figs 1⇑ and 3⇑). Hypoxic ventilatory stimulation during rapid and pronounced drops in O2 saturation in children might have contributed to their shorter apnoea duration (table 2⇑). Since adults did not desaturate more than children despite longer apnoeas they might have had larger O2 stores in relation to O2 consumption.
Actigraphic recordings suggested similar total sleep duration for children and adults. However, the current authors could not assess potential differences in sleep–wakefulness transitions and sleep-stage distribution that might have also influenced the breathing pattern at high altitude. Previous studies have shown that periodic breathing in hypoxia occurred less frequently in rapid eye movement compared with nonrapid eye movement sleep 5, 35, and children are known to spend a higher proportion of their total sleep time in rapid eye movement sleep. Differences in the ventilatory response to hypoxia and hypercapnia in children and adults may have further influenced the prevalence of periodic breathing 15.
Children sighed significantly more frequently during the night than adults, and their number of sighs increased with ascent from low to high altitude (table 2⇑). Although sighs have been described during sleep in infants 36 and in adults 28, their physiological significance is not well understood. Some data in infants suggest a role of sighs in restoration of lung mechanics 37 and in resetting the neuro-respiratory control system 36 or as a trigger of arousals during sleep 38. Sighs in pre-pubertal children have not been investigated in detail. As an increase in sigh frequency at altitude was found in children but not in adults (table 2⇑), sighs may represent a specific response of children to hypobaric hypoxia. Although the transient hypocapnia and neuromechanical inhibition (lung stretch reflex) 39 following sighs may trigger central apnoea, the prevalence of periodic breathing in children was significantly lower than in adults despite their higher number of sighs. This further corroborates the greater stability of their respiratory control system (fig. 2⇑).
Half of the children and 30% of adults experienced significant symptoms of AMS (defined by ≥0.7 points in the ESQ cerebral score) during their stay at 3,450 m (table 3⇑). For comparisons, all six children aged 6 months to 4 yrs and five out of 10 children aged 13–18 yrs had significant symptoms of AMS (defined by ≥3 points in the Lake Louise questionnaire) after one night at 3,500 m (very young children were evaluated with a modified questionnaire) 16. Since no statistical correlation was found among AMS scores and measures of periodic breathing in the current study, an interaction between periodic breathing and severity of AMS symptoms seems unlikely or weak. This is in agreement with the current authors' previous observations in adults ascending to Capanna Regina Margherita (4,559 m) 8.
The present authors did not identify any significant correlation among corresponding breathing pattern characteristics in children and their fathers, but as the study may not have been appropriately powered and did not include mothers, it might have failed to detect potential genetically determined properties of ventilatory control.
In summary, the present analysis of nocturnal breathing patterns in children and adults during acute high-altitude exposure revealed a similar degree of hypoxaemia and hyperventilation but significantly less periodic breathing in children compared with adults. This suggests a more stable control of ventilation in children than in adults at high altitude, which is related to their lower apnoea threshold for CO2, a larger CO2 reserve and a shorter circulation time.
Statement of interest
None declared.
Footnotes
-
Earn CME accreditation by answering questions about this article. You will find these at the back of the printed copy of this issue or online at www.erj.ersjournals.com/current.shtml
- Received September 10, 2007.
- Accepted February 4, 2008.
- © ERS Journals Ltd
References










