





Abstract
Epithelial cell injury and apoptosis are recognised as early features in idiopathic pulmonary fibrosis and bleomycin-induced fibrosis in mice. Telomerase is a known apoptosis-alleviating factor. The role of telomerase was studied during bleomycin-induced lung epithelial cell (LEC) apoptosis in vitro in a mouse LEC line, and in vivo in LECs isolated from bleomycin-treated mice.
The current authors evaluated changes in murine telomerase reverse transcriptase (mTERT) mRNA levels and changes in telomerase activity with the TRAPeze Detection Kit, telomeric length with the TeloTTAGGG Telomere Length Kit, and LEC apoptosis with FACScan and 4,6-diamino-2-phenylindole dihydrochloride stain.
There was a significant elevation in mTERT mRNA and a transient 41% increase in telomerase activity 24 h after in vitro bleomycin treatment. At 72 h, telomerase activity had fallen to 26% below levels in untreated cells. Reduction of telomerase activity over time, or by direct inhibition, significantly elevated LEC apoptosis. No change in average telomeric length was noted. In vivo, telomerase activity of LECs from bleomycin-treated mice increased at 7 and 14 days.
In conclusion, telomerase activity may play a protective role against robust bleomycin-induced lung epithelial cell apoptosis. Moreover, stabilising telomerase activity may decrease epithelial cell apoptosis and the resulting lung fibrosis.
Bleomycin-induced lung injury in mice is a well-established model of lung fibrosis 1. Following in vivo instillation of bleomycin to the trachea, apoptosis is induced in lung epithelial cells, which lose their potential to reconstitute a normal alveolar surface 2. When this occurs, normal lung epithelial cells are replaced with fibroblasts, leading to fibrosis 2–4. Inhibition of apoptosis could diminish epithelial cell replacement by fibroblasts, and thus reduce lung fibrosis.
Telomere length is maintained by telomerase, a ribonucleoprotein RNA-dependent DNA polymerase complex. Telomerase consists of an RNA template and a catalytic protein, telomerase reverse transcriptase (TERT) 5. Maintenance of telomerase activity is essential for the survival of proliferating cells 6–8. Knockout mice lacking the gene for TERT express higher levels of apoptosis and show liver fibrosis, both of which can be repaired by the introduction of telomerase 9.
The current authors have recently confirmed that bleomycin causes apoptosis of lung epithelial cells in vitro in a time- and dose-dependent manner 10, and also reported the specific induction of epithelial cell apoptosis by myofibroblasts from fibrotic lungs via the Fas/Fas ligand pathway 11. The aim of the present study was to evaluate telomerase activity in bleomycin-treated lung epithelial cells (LECs) and the resulting effect on apoptosis in bleomycin-treated mouse lung epithelial (MLE) cells. It was hypothesised that bleomycin injury would result in a reduction of telomerase activity in lung epithelial cells, in contrast to in fibroblasts, and thus an increase in LEC apoptosis.
It was found that bleomycin causes an initial increase, and then a reduction, in telomerase activity controlled, at least in part, at the mRNA level. When telomerase activity is diminished, significant apoptosis of epithelial cells is initiated. With further disruption of telomerase activity, apoptosis occurs at significantly higher levels.
In order to establish the relevance of these findings to the pathological process of lung fibrosis, the current authors evaluated telomerase activity in vivo in an experimental model of murine bleomycin-induced fibrosis. An early reduction of telomerase activity in lung epithelial cells isolated from bleomycin-treated mice was found. This was followed by a pattern similar to that encountered in vitro, with significant elevation of telomerase activity in the surviving cells, followed eventually by a reduction back to baseline levels.
MATERIALS AND METHODS
Cell line and culture
Experiments were conducted with the murine type II, simian vacuolating virus-40-transformed LEC line (ATCC, MLE-15). This extensively used cell line 11–13 was maintained in HITES (Ham's F12, insulin, transferin, β-estradiol and sodium selenite) medium, supplemented with 2% foetal bovine serum (Biological Industries, Beit Haemek, Israel).
Animals
Male, 11–12-week-old, C57B1/6 mice (Harlan Sprague–Dawley, Indianapolis, IN, USA), each weighing 25–35 g, were used. The local committee for animal experiments (Hadassah – Hebrew University Medical Center, Jerusalem, Israel) approved all animal care and experimental protocols.
Reagents
The reagents used were: propidium iodide (PI) stock solution, 1 mg·mL−1 (Calbiochem, La Jolla, CA, USA) in PBS; fluorescein isothiocyanate (FITC)-conjugated Annexin V (BD Pharmingen, San Diego, CA, USA); bleomycin (ASTA Medica, Frankfurt am Main, Germany); tetra(N-methyl-4-pyridyl)-porphyrin chloride (TMPyP4), a G-quadruplex-intercalating porphyrin telomerase inhibitor (Calbiochem, Cambridge, MA, USA); 4,6-diamino-2-phenylindole dihydrochloride (DAPI) stain (Sigma-Aldrich, St Louis, MO, USA); pentobarbitol 6% (Shoresh Pharm, Jerusalem, Israel); dispase II (Roche Diagnostics GmbH, Mannhein, Germany); Dulbecco/Vogt modified Eagle's minimal essential medium (DMEM; Sigma-Aldrich); DNase (Sigma-Aldrich); 4% paraformaldehyde (Sigma-Aldrich); and saponin buffer (Sigma-Aldrich).
Exposure of MLE line cells to bleomycin
MLE cells suspended in HITES medium were incubated with or without 0.1 unit·mL−1 bleomycin. This bleomycin dose was selected based on previous kinetic studies in the current authors’ laboratory, because at 24 h it induces significant, but not overwhelming, apoptosis 10. After incubation, trypsin (Biological Industries) was added for adherent cell removal and the mixture was centrifuged (253×g; 10 min). The pellet was resuspended for further evaluation and viable cells were counted using trypan blue (Sigma-Aldrich).
Evaluation of apoptosis
DAPI staining was used to assess the proportion of apoptotic cells, as previously described 12, 14. After elution of the media, MLE cells were washed with PBS (5 min; room temperature). Cold methanol (2 mL) was added to each plate, cells were reincubated (30 min; -20°C) and then rewashed, and sediment was placed on coverslips. DAPI stain (100 μL of 1 μg·mL−1) was added to the coverslips, which were then incubated in the dark (15 min; room temperature), washed twice with PBS and dried. The coverslips were placed on slides that had been prepared in advance with 20 μL of mounting solution. Slides were evaluated with a fluorescent microscope (Axiovert 200; Carl Zeiss AG, Oberkochen, Germany) and data were analysed (Image-Pro Plus 4.1; MediaCybernetics, Silver Spring, MD, USA). At least 100 cells were randomly selected and counted from each slide, and the average percentage of apoptotic cells from all slides in each group was reported.
Flow cytometry analysis with FACScan (Becton Dickinson, Mountain View, CA, USA) was used in one of two ways as a second method to evaluate apoptosis, as follows.
1) Cells (106·mL−1) were incubated (15 min; room temperature) with 5 μL·mL−1 of FITC-conjugated Annexin V, collected and pelleted. PI (1 μg·mL−1) was added to the cells (5 min; room temperature). Flow cytometry analysis (FL1/FITC-Annexin V plotted against FL2/PI-positive cells) was performed with FACScan, and data were analysed with CellQuest-ProTM software (Becton Dickinson). PI-positive cells were excluded from the presented data.
2) Due to the autofluorescence of TMPyP4, Annexin V measurement was not possible when TMPyP4 was used. In this situation, cells (106·mL−1) were evaluated for size and granularity (forward and side scatter) using FACScan. The population of apoptotic cells was characterised by their smaller size, implying apoptotic shrinkage, as well as their higher granularity, representing apoptotic cytoplasmic condensation, as previously reported 15, 16.
RNA isolation
Total cellular RNA was isolated from cell pellets using TRI Reagent (Sigma-Aldrich) supplemented by 1 μg·mL−1 of glycogen, according to the manufacturer’s protocol. To assess RNA integrity and verify the absence of a high molecular weight band representing contamination with genomic DNA, an aliquot of each sample was analysed by 1% agarose gel electrophoresis. RNA was also analysed by spectrophotometer for assessment of integrity, protein contamination (ratio of absorbance at 260/280 nm >1.8) and quantification.
Semiquantitative RT-PCR of mTERT
RNA was reverse-transcribed to cDNA using an avian myeloblastosis virus reverse-transcription (RT)-based protocol with random primers, as well as poly(dT) (Reverse Transcription System; Promega, Madison, WI, USA). From each sample, 1 μg was uniformly used for RT. Taq DNA polymerase (0.25 mL; Roche Diagnostics GmbH) was added to each tube following hot-start incubation (5 min; 95°C). This was followed by 40 cycles for murine TERT (mTERT) and 28 cycles for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) denaturation (15 s; 96°C), annealing (20 s; 65°C), extension (1 min; 72°C) and a final extension step (5 min). A total of 2 μL of forward and reverse primer sequences (Sigma-Aldrich) were used as follows. Forward mTERT: 5′-GGGAGATGGCCAAGAGCGTCTAAA-3′; reverse mTERT: 5′-CGGTGGGCTGGTGTTCAAGG-3′; forward GAPDH: 5′-GTTGCCATCAATGACCCCTTC-3′; and reverse GAPDH: 5′-CATGTGGGCCATGAGGTCCAC-3′. The number of cycles was predetermined to be the greatest number in which amplification was within the linear range. PCR products were analysed by electrophoresis on a 1% agarose gel stained with ethidium bromide (Sigma-Aldrich). The intensity of the fluorescent signal emitted by the PCR products was determined by densitometry using a tabletop scanner (Multi-AnalystTM PC Version 1.1; Bio-Rad Laboratories, Hercules, CA, USA). Data were analysed with the Fluor-S-MultiImager (Bio-Rad Laboratories). Each cDNA was amplified in triplicate, corrected to the level of GAPDH mRNA, and the median value used. Amplification was repeated with a smaller quantity of substrate if the densitometer signal was beyond the predetermined linear range.
Detection of telomerase activity
Telomerase activity was detected using the TRAPeze Telomerase Detection Kit (Intergen, Purchase, NY, USA), a PCR-based telomeric repeat amplification protocol (TRAP) method, as previously described 17. Briefly, cell extracts were resuspended in CHAPS lysis buffer and combined with the reaction mix in RNase-free PCR tubes, adding a varied number of telomeric repeats onto the substrate oligonucleotide according to telomerase activity. Positive and negative controls were examined accordingly. Protein levels were evaluated using the Bradford method, and equal amounts of protein were evaluated. PCR amplification was then performed. Loading dye was added to each reaction tube, and the mixture was loaded onto a 12.5% nondenaturising SDS-PAGE gel. Following electrophoresis, the gel was stained with ethidium bromide, and detection was determined by densitometry using a tabletop scanner (Bio-Rad Laboratories). The total and relative quantities of telomerase product for each sample were calculated using the Fluor-S-MultiImager.
Measurement of terminal restriction fragment (telomeric) length
Telomeric length was determined as previously described 17, 18 using the TeloTTAGGG Telomere Length Assay Kit (Roche Diagnostics GmbH). Briefly, genomic DNA was isolated and digested with Hinf1/Rsa, DNA fragments were separated by electrophoresis on agarose gel, and Southern blot transfer was performed. The membrane was hybridised with a telomere-specific digoxigenin (DIG)-labelled probe, incubated with anti-DIG alkaline phosphatase, and assessed with chemiluminescence detection (Kodak X-OMAT 2000 Processor; Kodak, Rochester, NY, USA). Average telomeric length was determined using Telometric 1.2 (Fox Chase Cancer Center, Philadelphia, PA, USA).
Telomerase inhibition
Telomerase activity was inhibited in MLE cells using 1.5 mM of TMPyP4, and inhibition was confirmed using the TRAPeze Telomerase Detection Kit.
Intratracheal instillation of animals
Intratracheal instillation of bleomycin or saline to mice was performed as has been previously described 1, 19. Briefly, mice were anaesthetised intraperitoneally. The trachea was exposed and a metal cannula was carefully inserted into the trachea. Bleomycin (0.08 mg) in 0.1 mL of 0.9% saline, or saline alone was injected.
LEC isolation for ex vivo culture
LECs were isolated as has been previously reported 11. Briefly, animals were sacrificed by aortic transection while under pentobarbitol 6% anaesthesia. The lungs were filled with 3 mL dispase II and allowed to collapse. Low-melt agarose (0.5 mL) was then infused, and lungs were covered with ice for 2 min. The lungs were excised from the animals and incubated in 1 mL dispase II in a 12-mL polypropylene culture tube (45 min; room temperature). The lungs were transferred to 7 mL DMEM with 0.01% type II DNase in a 60-mm petri dish. Digested tissue was gently swirled for 5–10 min. The resulting suspension was filtered through 100-μm and 40-μm Falcon strainers, and then through a 22-μm nylon mesh, centrifuged (142×g; 12 min; room temperature) and resuspended in culture medium. The cells were placed on culture plates previously coated with mouse anti-CD45 and mouse anti-CD16/32 monoclonal antibodies (IQ Products, Groningen, the Netherlands). Cells were incubated (2 h; 37°C), transferred to new uncoated culture plates (45 min; 37°C) and collected by centrifugation.
Evaluation of isolated LECs for purity by flow cytometry
In order to assess the purity of isolated epithelial cells, both extracellular and intracellular markers were evaluated.
Extracellular staining
LECs were fixed with 4% paraformaldehyde (10 min; room temperature) and then washed with PBS. LECs were then incubated (30 min; room temperature) with 10 μL/100 μL of one of four antibodies: 1) FITC-conjugated lymphocyte marker (anti-CD3, IQ Products); 2) FITC-conjugated leukocyte marker (anti-CD31; BD Pharmingen); 3) FITC-conjugated macrophage marker (F4/80; Serotec, Raleigh, NC, USA); and 4) type I epithelial cell marker (AQ5, non-FITC conjugated, with addition of secondary antibody FITC-conjugated anti-goat; Jackson Immunoresearch Laboratories, West Grove, PA, USA). The cells were washed twice in saponin buffer and analysed by flow cytometry.
Intracellular staining
Cells were washed with 0.1% saponin buffer and then incubated (30 min; room temperature) with 1 μL/100 μL of one of the following antibodies: 1) type II epithelial markers (anti-surfactant protein (SP)B or anti-SPC; Chemicon, Temecula, CA, USA); or 2) myofibroblast marker (anti-α-smooth muscle actin FITC conjugate; Chemicon). For SPB and SPC staining, 1 μg/100 μL of FITC-conjugated anti-rabbit antibodies (Jackson Immunoresearch Laboratories) was added and cells were reincubated (30 min; room temperature). The cells were then washed twice in saponin buffer and analysed by flow cytometry.
Following this two-step evaluation, ∼90% of the isolated cells were found to be type I or II lung epithelial cells.
Statistical analysis
Analysis of variance was performed with the Kruskal–Wallis test for nonparametric data. When Kruskal–Wallis tests of comparability were statistically significant, Mann–Whitney comparisons with Holms sequential Bonferroni corrected p-values were performed. In order to determine whether relative telomerase activity was consistently greater or less than baseline, the data were dichotomised. One sample Chi-squared test was performed, comparing the observed distribution with an expected random distribution. A p-value <0.05 was considered significant.
RESULTS
Bleomycin modifies telomerase activity in vitro
In order to study the effects of bleomycin on telomerase activity of LECs, MLE cells were exposed to bleomycin for 24, 48 or 72 h. Cells were collected and resuspended. Viable cells were counted using trypan blue in order to exclude necrotic cells. A similar number of viable cells were taken for evaluation of telomerase activity from each group of bleomycin-treated and untreated cells. Telomerase activity increased in bleomycin-treated cells, compared with untreated control cells, by a mean of 41 and 12% at 24 and 48 h respectively. The mean activity then decreased to 26% below control levels at 72 h (fig. 1⇓). At 96 h, bleomycin caused excessive cell death, precluding measurement in four out of six experiments. However, in two successful experiments the mean reduction of telomerase activity at 96 h in bleomycin-treated compared with control cells was 43% (data not shown). Figure 1⇓ shows the level of telomerase activity in bleomycin-treated compared with untreated control cells at each time point. A regression analysis of the percentage change versus time of bleomycin exposure yielded the following linear equation: ratio = 1.771−0.307×exposure days (r2 = 0.83; p = 0.023). The level of telomerase activity in bleomycin-treated cells compared with untreated control cells was significantly higher at 24 h (p = 0.014) and lower at 72 h (p = 0.014).

Bleomycin modifies telomerase activity. Telomerase activity was assessed in bleomycin-treated and untreated mouse lung epithelial cells using the TRAPeze Telomerase Detection Kit. The percentage change of telomerase activity in bleomycin-treated cells, compared with untreated control cells, at 24, 48 and 72 h is presented. Each point represents an independent comparison between bleomycin-treated and control cells. *: p<0.05.
Bleomycin increases expression of mTERT mRNA
The transcriptional regulation of the catalytic component of telomerase, mTERT, is recognised as one of the major rate-limiting steps in the induction of telomerase activity 5. In order to assess the regulation level of telomerase activity in bleomycin-treated MLE cells, mTERT mRNA levels were measured. Cells were exposed to bleomycin for 24, 48 or 72 h. mTERT mRNA in bleomycin-treated MLE cells was measured by semiquantitative RT-PCR, and compared with the median level of untreated control cells at each time point. Bleomycin induced a substantial increment of mTERT mRNA at 24 h. There was some elevation at 48 h, although considerably less significant. The level of mTERT mRNA in bleomycin-treated MLE cells returned nearly to control levels at 72 h (fig. 2⇓).

Bleomycin increases expression of murine telomerase reverse transcriptase (mTERT) mRNA. The level of mTERT mRNA was assessed in mouse lung epithelial cells using semiquantitative reverse transcription-PCR. The ratio between the level of mTERT mRNA in bleomycin-treated cells and the median level of mTERT mRNA in untreated control cells, at each time point, both adjusted to glyceraldehyde-3-phosphate dehydrogenase, is presented. Each point represents an independent experiment. ……: point of equality between the two groups.
Bleomycin does not change average telomeric length
Telomerase inhibition can induce apoptosis in several ways, but telomere shortening is generally accepted to be the primary mechanism 7. In order to assess bleomycin-induced changes in telomeric length, the average terminal restriction fragment length was measured using the TeloTTAGGG Telomere Length Assay Kit (Roche Diagnostics GmbH). In three independent experiments, there was no difference between average telomeric length in MLE cells exposed to bleomycin compared with controls at 24 or 72 h. The results shown in figure 3⇓ are representative of four experiments.

Bleomycin does not change telomeric length. A representative experiment showing the results of average telomeric length measurements at 24 and 72 h. No difference was seen between the calculated average telomeric length in bleomycin-treated mouse lung epithelial cells (Bleo) and untreated control cells. M: size marker.
Elevation in telomerase activity delays robust apoptosis
The current authors performed kinetic evaluations of telomerase activity levels and apoptosis rates in bleomycin-treated MLE cells at 6–48 h. Telomerase activity was assessed with the TRAPeze Telomerase Detection Kit, and the percentage of apoptotic cells was estimated using double staining with Annexin V and PI. A representative experiment is shown in figure 4⇓. There was an initial elevation of relative telomerase activity in bleomycin-treated cells, followed by a decrease in activity that was associated with a significant increase in apoptosis.
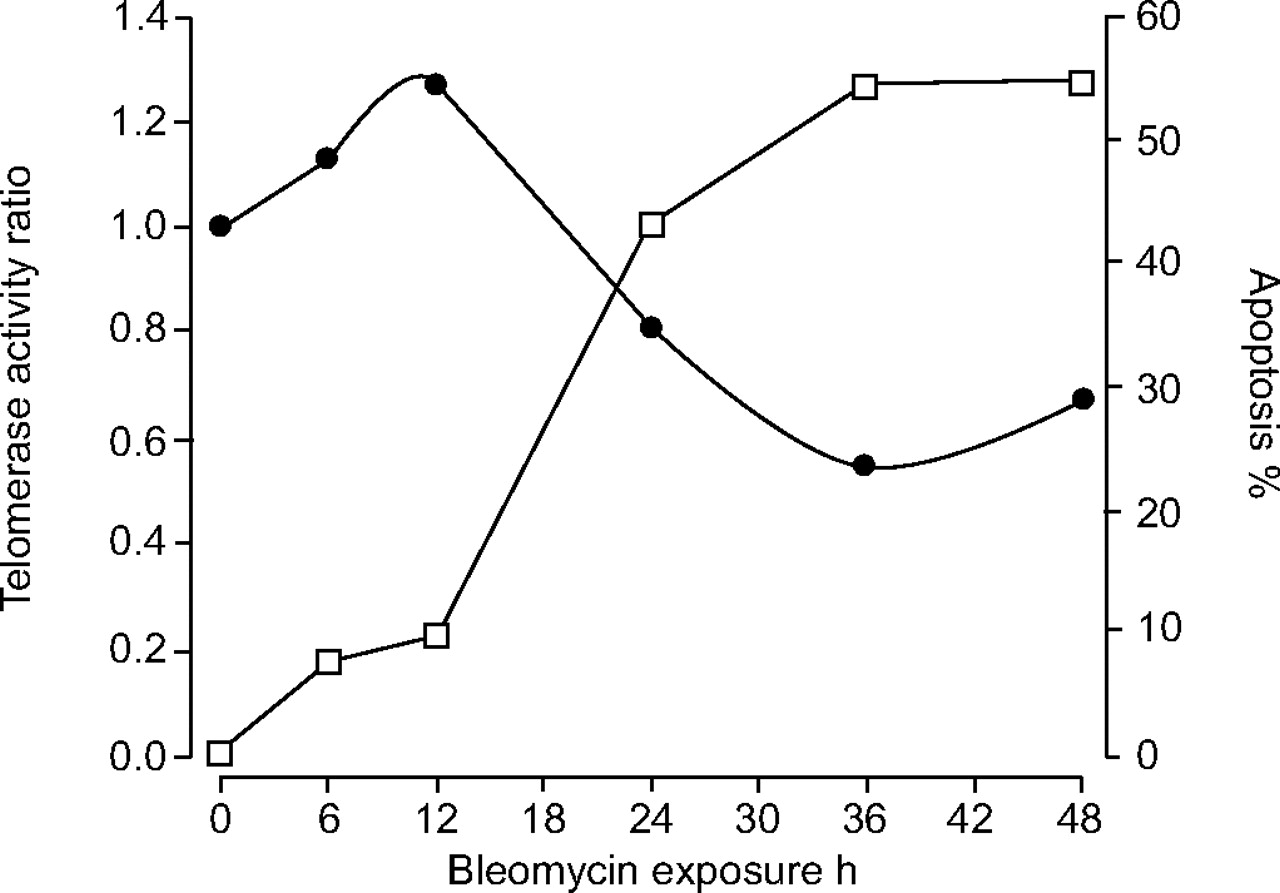
Elevation in telomerase activity delays robust apoptosis. Telomerase activity at 6–48 h, assessed with the TRAPeze Telomerase Detection Kit, is expressed as the ratio of bleomycin-treated to untreated control mouse lung epithelial cells (•). The percentage of apoptosis in bleomycin-treated cells was evaluated with Annexin V/propidium iodide staining (□).
Inhibition of telomerase activity by TMPyP4 increases epithelial cell apoptosis
TMPyP4 is a known G-quadriplex inhibitor of telomerase activity 20. TMPyP4 exposure alone caused apoptosis in MLE cells at rates similar to bleomycin exposure (data not shown). In order to establish the relationship between changes in telomerase activity and apoptosis, bleomycin-treated MLE cells were exposed to TMPyP4 for 24 h. TMPyP4 was confirmed to inhibit the bleomycin-induced elevation in telomerase activity using the TRAPeze Telomerase Detection Kit (data not shown). Apoptosis, evaluated using the DAPI stain, was higher in bleomycin-treated MLE than untreated control cells, and increased appreciably in bleomycin-treated MLE cells that were exposed to the telomerase inhibitor TMPyP4 (figs 5a–c⇓). For quantification, the proportion of apoptotic cells was calculated in 200 random cells on every DAPI-stained slide in four independent experiments. The mean proportion of MLE apoptotic cells was 81.3% in bleomycin and TMPyP4-treated cells compared with 43.4% in cells treated with bleomycin alone (p<0.05; fig. 5g⇓).
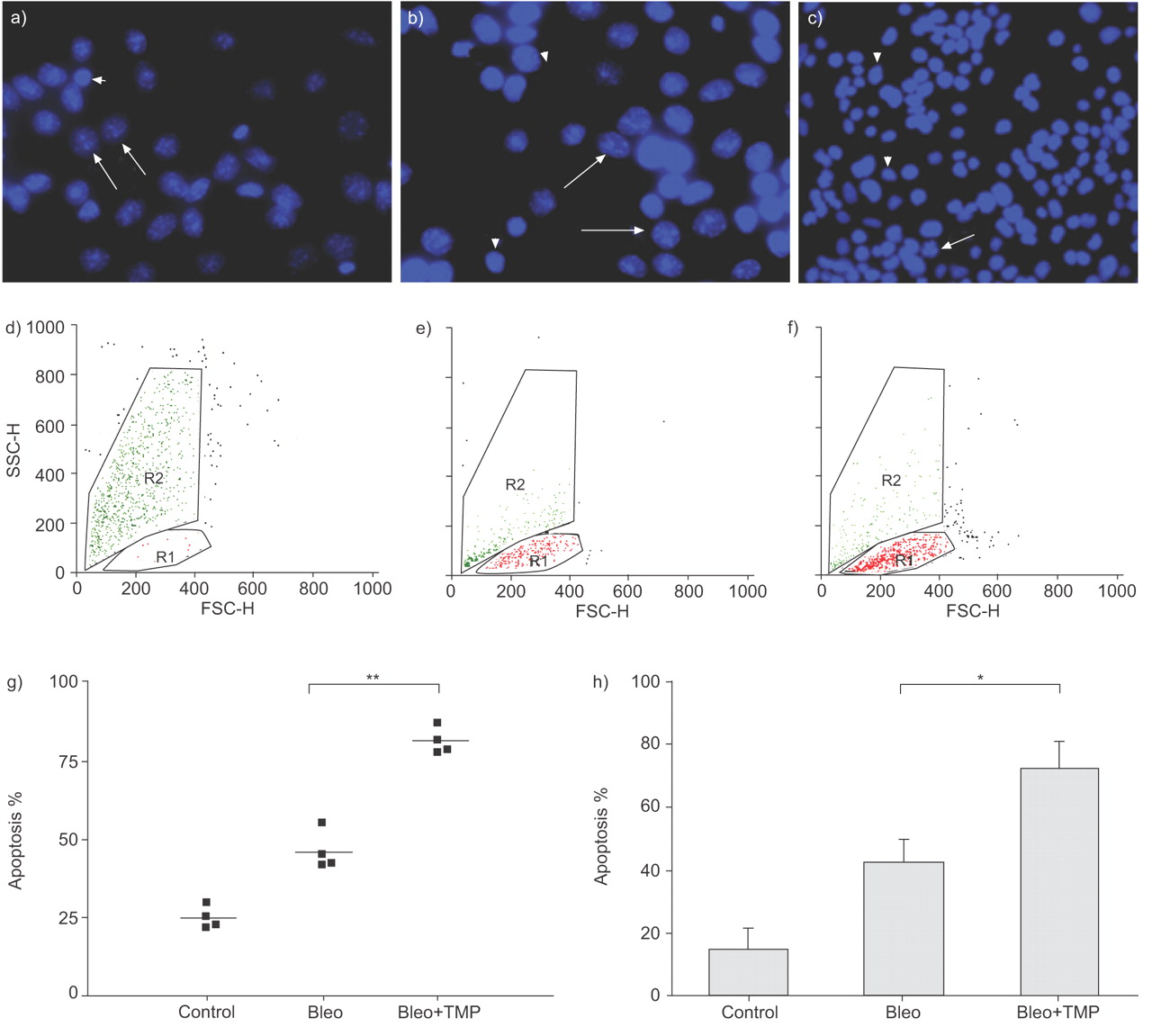
Inhibition of telomerase activity increases bleomycin-induced apoptosis. Using 4,6-diamino-2-phenylindole dihydrochloride (DAPI) stain for the identification of apoptotic cells, a) presents the increased percent of apoptotic cells in bleomycin- and tetra(N-methyl-4-pyridyl)-porphyrin chloride-treated mouse lung epithelial (MLE) cells (Bleo+TMP) at 24 h, compared with, in b), cells treated solely with bleomycin (Bleo). c) Minimal apoptosis is shown in untreated control cells. The speckled cells are apoptotic (arrows), while the smooth cells are nonapoptotic (arrowheads). Also shown are representative FACS analyses of apoptosis at 24 h in the Bleo+TMP (d), Bleo (e) and control (f) cells. Apoptotic cells were identified by size and granularity (forward (FSC-H) and side (SSC-H) scatter height). R1: region of nonapoptotic cells; R2: region of apoptotic cells. g) The proportion of apoptotic MLE cells each represented by a point, at 24 h in four independent experiments using DAPI stain. Mean values are presented by solid lines. h) Mean±sd values representing the proportion of apoptotic MLE cells in seven independent experiments using FACS analysis. *: p<0.05; **: p<0.01.
Apoptosis was further evaluated using flow cytometry analysis of size and granularity as previously described 15, 21. Cells exposed to both bleomycin and telomerase inhibition had a significantly higher proportion of apoptotic cells when compared with treatment with bleomycin alone (fig. 5h⇑). Representative dot plots are presented in figures 5d–f⇑.
Bleomycin modifies telomerase activity in LECs in vivo
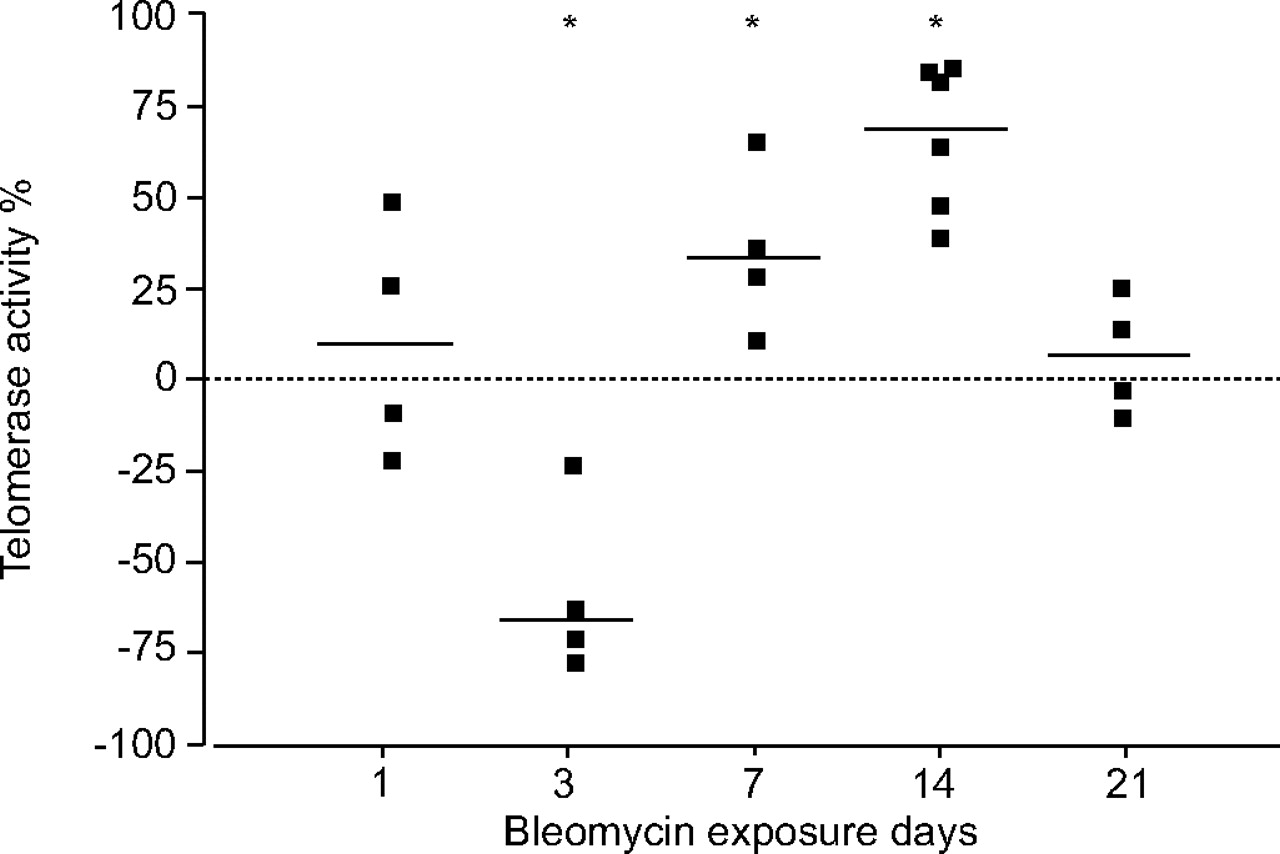
Epithelial cells were isolated from mouse lungs 1, 3, 7, 14 and 21 days after intratracheal instillation of bleomycin or saline. The development of fibrosis was confirmed in bleomycin-treated mice, as previously reported 1. Changes in telomerase activity of LEC isolated from bleomycin-treated compared with saline-treated mice are presented in figure 6⇓. Telomerase activity decreased at day 3 by a mean of 59% (p = 0.046). At day 7, there was a mean elevation of 36% in telomerase activity in surviving LEC (p = 0.046), with a further increase at day 14 by a mean of 68% (p = 0.014). No significant change in telomerase activity in bleomycin-treated compared with control mice was found at day 21, when the fibrotic process subsided (fig. 6⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bleomycin modifies telomerase activity in lung epithelial cells (LECs) in vivo. Telomerase activity was assessed in LECs isolated from bleomycin- and saline-treated (control) mice using the TRAPeze Telomerase Detection Kit. The percentage change in telomerase activity of LECs isolated from bleomycin-treated in relation to saline-treated control mice at 1, 3, 7, 14 and 21 days post intratracheal instillation is presented. Each point represents an independent value. *: p<0.05.
DISCUSSION
In the present study, the role of telomerase during bleomycin-induced apoptosis was studied in LECs in vitro and LECs isolated from mice with bleomycin-induced lung fibrosis in vivo.
It was found that bleomycin causes an initial elevation of telomerase activity, followed by a reduction, in MLE cells in vitro. Initial bleomycin-induced apoptosis has occurred at 12 h, as telomerase activity increases. Subsequent reduction of telomerase activity over time results in substantially increased apoptosis (fig. 1⇑).
In order to establish the relevance of these findings to the evolution of lung fibrosis, telomerase activity was evaluated in vivo in LECs isolated from bleomycin-treated mice. At day 7, and even more so at day 14, LECs isolated from bleomycin-treated mice showed a significant elevation of telomerase activity compared with LECs from saline-treated control mice. Increased telomerase activity is possibly a mechanism that enables these cells to outlive the attack of neighbouring inflammatory and other effector cells. When the fibrotic process subsided, at day 21, telomerase activity dropped down to the levels found in control mice.
Mounting evidence indicates that telomerase activity protects cells from apoptosis via multiple mechanisms 7, 8, 22, 23. In the current study, the anti-apoptotic effects of telomerase was shown in murine lung epithelial cells, where a transient, bleomycin-induced elevation of telomerase activity contributed to a significant delay in bleomycin-induced apoptosis (fig. 4⇑). Klapper et al. 24 reported a similar transient elevation in telomerase activity induced in human promyelocytic leukaemia HL60 cells by etoposide, another DNA-damaging agent.
Early apoptosis is mediated by the process of DNA damage 10, 25, and telomerase activity does not protect from apoptosis at these early time points. Telomerase activity increases only after apoptosis has initiated (fig. 4⇑). It is possible that apoptosis is the trigger for the induction of telomerase as a defence mechanism from bleomycin-induced robust apoptosis. Therefore, the initial increase in bleomycin-induced telomerase activity, possibly mediated by apoptosis, may protect MLE cells from further apoptosis. Indeed, when the bleomycin-induced increase in telomerase activity diminishes over time (fig. 1⇑), the level of bleomycin-induced apoptosis increases significantly. The fact that robust apoptosis is induced by specific inhibition of telomerase with TMPyP4 (fig. 5⇑) strengthens the hypothesis that this response is not merely correlative.
The current authors have previously shown that bleomycin causes significant DNA damage in MLE cells 10. Masutomi et al. 26 showed that suppression of human TERT abrogates the cellular repair response to DNA double-strand breaks. It is therefore reasonable to assume that telomerase activity is elevated as part of the cellular reaction to that damage. The present work is further supported by the results of Rubio et al. 27, who showed that telomerase expression and elevation of its activity increased the resistance of cell lines to bleomycin. Taken together, these results suggest that resistance to bleomycin injury could be related, at least partly, to the inhibition of apoptosis induced by the telomerase complex. The current authors’ data analysis, using PI staining followed by flow cytometry, excludes necrotic and dying cells, as opposed to apoptotic cells. This process argues against the possibility that changes in telomerase activity are merely a toxic effect of bleomycin. Elevations in telomerase activity and mRNA support the concept that this reaction is part of an active phenomenon.
Telomere shortening is the main mechanism for apoptosis induction following telomerase inhibition 7. Massive telomere loss has been shown to occur early in DNA-damage-induced apoptosis 28. However, there is accumulating evidence to suggest that telomerase also plays an important role in protecting karyotypic stability, independent of its part in maintaining telomeric length 22. Rubio et al. 27 reported that resistance to injury was related to telomerase activity only in cell lines with short telomeres, suggesting that telomere shortening is the cause of apoptosis. In contrast, the current authors found no change in the average length of telomeres in bleomycin-treated MLE cells for up to 72 h, a time by which significant apoptosis has typically occurred. This finding agrees with the accepted convention that telomere shortening is a slow-acting mechanism for apoptosis induction 5. It is possible that a limited and more specific telomeric loss (i.e. in a specific chromosome) is responsible for the induction of robust LEC apoptosis. However, it is clear that general massive telomere loss is probably not primarily responsible for apoptosis induction in these cells. Two rapid mechanisms of telomerase activity have been described and could account for the protection from apoptosis without notable change in telomeric length. These are telomere capping 7, 29 and regulation of gene expression by telomerase 23. Induction of telomerase activity has been shown to significantly influence the expression of several oncogenes 30, and the absence of telomerase could activate genes involved in apoptosis 7.
The transcriptional regulation of the catalytic component of telomerase expression is recognised as one of the major rate-limiting steps in the induction of telomerase activity 5. The current authors have found a rapid, substantial elevation in mTERT mRNA as early as 24 h after treatment with bleomycin. The bleomycin-induced elevation in telomerase activity is therefore regulated, at least in part, by this transcription.
Measurement of telomere elongation was the method used to assess telomerase activity. These measurements revealed a modest increase in telomerase activity at 24 h (fig. 1⇑) versus a substantial elevation of mTERT mRNA (fig. 2⇑). The lack of correlation between these measurements supports the concept that telomerase protected LECs from apoptosis in a manner unrelated to telomere elongation and telomeric loss.
Increasing evidence suggests that lung epithelial cell apoptosis is an important contributor to bleomycin-induced lung fibrosis 2, 31. One of the main theories for the pathophysiology of idiopathic pulmonary fibrosis (IPF) is that injury to the pulmonary parenchyma causes epithelial cell damage and apoptosis 32. Evidence of type II pneumocyte apoptosis in normal alveoli of IPF patients has been previously reported 33, and loss of alveolar epithelium, with failure of re-epithelisation, was proposed to stimulate persistence and progression of fibrosis 32.
The current authors have recently shown that myofibroblasts, which are known to accumulate abundantly in the lungs of mice with bleomycin-induced fibrosis 3, 34, are not only collagen producers, but also function as effector cells 11. It was also demonstrated that they induce apoptosis of lung epithelial cells in vivo, a possible mechanism for the prevention of lung tissue remodelling 11. Apoptosis thus plays a role in the induction of fibrosis.
In the present work, it was found that telomerase activity may have a role in bleomycin-induced lung fibrosis in vivo. Shortly after bleomycin injury, telomerase activity in LECs decreases. However, at days 7 and 14, telomerase activity increases, enabling epithelial cell regeneration, while epithelial cells without elevated telomerase activity may have already undergone apoptosis. Since elevated telomerase activity has been repeatedly shown to assist cell survival 7, 8, 22, 23, it is to be expected that, during intensive fibrosis (on days 7 and 14), surviving epithelial cells have levels of telomerase activity that are higher than baseline (fig. 6⇑). The current authors suggest that this elevation in telomerase activity is a cellular defence mechanism against active myofibroblast-induced LEC apoptosis.
In the past, the current authors have shown that the fibrotic process in bleomycin-treated C57Bl/6J mice subsides at day 21 1. It is therefore not surprising that, at this time point, when the lung regeneration process has advanced and elevation of telomerase activity confers no survival advantage, it returns to normal levels in the remaining epithelial cells.
The impact of bleomycin-induced injury on telomerase activity is different in lung fibroblasts and epithelial cells. Nozaki et al. 4 noted a constant elevation in telomerase activity in fibroblasts following bleomycin instillation with extended life span. In contrast, the present study shows that epithelial cells have only a transient elevation of telomerase activity, followed by a significant reduction, resulting in robust apoptosis.
The most important question arising from the present work is the relationship between changes in telomerase activity and human lung fibrosis. Mutations in the telomerase complex are linked to dyskeratosis congenita, a rare, inherited syndrome of bone marrow failure 35. Pulmonary fibrosis is a recognised manifestation of this syndrome 36. Furthermore, strong support for the relationship between an altered telomerase complex and IPF has been very recently published, demonstrating that mutations in the genes encoding telomerase can appear in familial IPF 37. The current authors’ work on epithelial cells, together with other research carried out on fibroblasts 3, 8, suggests the possibility of a relationship between telomerase mutations and lung fibrosis. Therefore, changes in telomerase activity could alter the differential reaction of LECs and fibroblasts to lung injury.
The present results demonstrate that changes in telomerase activity, regulated at least in part by transcription, play a role in LEC apoptosis by a mechanism that is independent of telomere loss. The elevation of telomerase activity in LECs in vivo may represent an active defence process in surviving LECs against neighbouring myofibroblasts that may cause apoptosis of epithelial cells as part of the fibrotic process. Those epithelial cells that activate a defence mechanism, including, but probably not limited to, elevation of telomerase activity, could escape apoptosis, survive the injury and help reduce the extent of fibrosis.
In conclusion, a novel possibility is presented for regulation of DNA-damage-induced lung injury through modifying telomerase activity. Understanding the role of telomerase in the apoptosis of epithelial cells is an important step in the evaluation of lung injury and possibly of fibrosis related to DNA damage. Therefore, stabilising telomerase activity in lung epithelial cells may decrease epithelial cell apoptosis and eventual lung fibrosis. This hypothesis is strongly supported by the recent reports that mutations in genes encoding telomerase have been found in familial idiopathic pulmonary fibrosis 37.
Acknowledgments
The authors wish to thank S. Fraifeld (Hadassah – Hebrew University Medical Center) for her editorial assistance in the preparation of this manuscript.
- Received January 24, 2007.
- Accepted April 30, 2007.
- © ERS Journals Ltd
References










