





Abstract
Galectins-1 and -3 regulate epithelial proliferation/apoptosis and neutrophil activation, and are implicated in lung cancer and asthma. The role of galectins in chronic obstructive pulmonary disease (COPD), characterised by epithelial changes and neutrophil infiltration, remains unknown.
In the present study, galectin-1 and -3 expression was assessed by immunohistology in the bronchial epithelium of lung specimens from eight severe COPD patients and compared with nine nonsmokers and six smokers without COPD. Findings were related to epithelial proliferation (Ki-67), tissue inflammation and lung function.
Epithelial galectin-3 immunostaining was increased only in the small airways of COPD patients when compared with nonsmokers and smokers. In contrast, galectin-1 was only significantly increased in the small airways of the group of smokers. Ki-67+ epithelial cells and neutrophils were increased in the small airways of COPD patients when compared with smokers. Furthermore, intra-epithelial neutrophils correlated in the small airways with Ki-67+ epithelial cells and with the forced expiratory volume in one second/forced vital capacity ratio. However, no correlation was observed with galectin expression.
The present study supports the hypothesis that distal airways represent an important site for detecting changes in chronic obstructive pulmonary disease. In patients with severe disease, an increased galectin-3 expression and neutrophil accumulation in the small airway epithelium was demonstrated, correlating with epithelial proliferation and airway obstruction.
Chronic obstructive pulmonary disease (COPD) is a frequent disorder characterised mostly by irreversible airway obstruction that develops in susceptible smokers 1. <1?show=[to]?>During the course of the disease, COPD airways are infiltrated by neutrophils, macrophages and T-cells (especially CD8+) through the upregulation of adhesion molecules, such as intercellular adhesion molecule-1 or CXC chemokine receptor 3 2, 3. However, the pathways involved in the persistence of recruited leukocytes, especially neutrophils, in the bronchi of COPD patients remain to be identified. Furthermore, there are differences in the leukocyte phenotypes present in COPD airways with regard to disease severity and micro-localisation within bronchial tissues 4.
In addition to inflammatory changes, COPD airways also undergo structural remodelling 5. This includes: 1) increased number and size of mucous acini in submucosal glands; 2) hyperplasia and metaplasia of surface mucus-secreting goblet cells in large and small airways, respectively; and 3) squamous metaplasia 5. The mechanisms leading to the aforementioned changes are unknown. Additionally, the bronchial epithelium function is affected in COPD patients, with upregulated expression of cytokines/chemokines 6, such as interleukin (IL)-8 or macrophage inflammatory protein-1α and -1β, and downregulation of proteins, such as the polymeric immunoglobulin (Ig) receptor 7, probably following degradation by neutrophil enzymes 8.
Galectins are galactose-binding proteins that are involved in lung physiology 9–12. Galectin-3 can enhance the proliferation of respiratory epithelial cells by limiting adhesion to the basement membrane through competition with integrin receptors for the binding of laminin in the extracellular matrix 13. Galectins-1 and -3 are also involved in apoptosis regulation via binding to the cell surface or cytoplasmic ligands, such as glycans of the fibronectin receptor, CD7 or intracellular proteins, such as B-cell leukaemia/lymphoma 2 gene product 10, 14. The nuclear and/or cytoplasmic pattern of galectin-3 expression could play a role during malignant transformation, as suggested in breast cancer 15. In addition, galectin-3 is known to: 1) trigger the release of oxygen metabolites and elastase by neutrophils and macrophages; 2) enhance macrophage phagocytosis; and 3) stimulate natural killer cell activity and IL-1 production 14, 16. It also represents an adhesion molecule independent from selectins and β2-integrins that underlie neutrophil recruitment to the lung during streptococcal pneumonia 17, 18. In contrast, galectin-1 inhibits tumour necrosis factor-α production by monocytes/macrophages 14, suggesting that galectin-1 and -3 have balanced, opposite effects on cell apoptosis/proliferation and phagocyte activation. It is noteworthy that these two lectins are abundantly expressed in the lung 11, 18.
The role of galectin-3 has been evaluated in lung cancer and asthma, whereas its role in COPD remains unknown. Therefore, the expression of galectins-1 and -3 in the bronchial epithelium of both large and small airways was evaluated by immunohistochemistry from eight patients with severe COPD undergoing lung transplantation. Lung specimens from nine nonsmokers and six smokers without COPD were used as controls. Findings on galectin expression were related to clinical parameters of airflow limitation, epithelial proliferation (Ki-67 expression) and tissue accumulation of neutrophils, macrophages and T-cells in the epithelium and lamina propria, in both large and small airways.
MATERIALS AND METHODS
Patients
COPD patients were recruited from a population undergoing lung transplantation, as previously described elsewhere 7. Eight very severe COPD patients classified as stage IV, according to the Global Initiative for Chronic Obstructive Lung Disease classification 1, were included. The mean (range) age was 52 (42–60) yrs. The COPD patients were ex-smokers, had symptoms of chronic bronchitis and all but one were treated by inhaled corticosteroids. Controls consisted of six patients, aged 60 (42–74) yrs, recruited from a population of smokers without COPD and nine nonsmokers (mean age 62 yrs) undergoing lung surgery for a solitary peripheral lung tumour. All patients were clinically stable with no evidence of lung infection at the time of surgery. Pulmonary function tests were performed 37±26 (mean±sd) days before surgery. For each patient, random tissue blocks (three to 11 per patient) were sampled in central and peripheral areas from each lobe of one or both lungs (single or bipulmonary transplantation), fixed under constant inflation pressure (30 cmH2O) in 4% formaldehyde and processed for immunohistochemistry. Patient characteristics are presented in table 1⇓.
Patient characteristics
Immunohistochemistry
Tissue sections were processed for galectin immunostaining. Endogenous peroxidase activity was inhibited by incubation in 0.03% v/v H2O2 and sections were blocked by normal horse serum and avidin/biotin blocking kit (Dako, Glostrup, Denmark). Sections were then treated for 30 min with 1:100 rabbit polyclonal anti-human galectin-1 or -3 antibodies, which had been rigorously tested for specificity by ELISA and Western blotting 12, 19, 20. Control sections were incubated with normal rabbit serum (1:100). Biotinylated horse anti-rabbit IgG (1:100) was used as secondary antibody and sections were incubated with avidin-biotin-horseradish peroxidase (ABC standard kit of Vectorstain; Dako). After revelation with 3,3'-diaminobenzidine tetrahydrochloride and 0.03% H2O2, sections were counterstained with Toluidine blue.
Leukocytes were detected using specific antibodies to neutrophil elastase (neutrophils), CD68 (monocytes/macrophages) and CD3 (T-lymphocytes). For macrophages and T-cells, antigen retrieval was performed using microwave treatment (five cycles of 3 min, 350 Watts) in 0.01 M citrate buffer, pH 5.7. Endogenous peroxidase activity was inhibited by incubation in 0.03% H2O2 and sections were blocked with 1% bovine serum albumin. Sections were then incubated with mouse monoclonal antibodies to neutrophil elastase (mIgG1κ, clone NP57; Dako) or CD68 (mIgG3κ, clone PG-M1; Dako) or with polyclonal rabbit antibody to CD3 (Dako). Control sections were treated as appropriate with normal rabbit serum or mouse IgG isotype. Secondary antibodies were biotinylated goat anti-rabbit IgG or goat anti-mouse IgG (Dako). The reaction was amplified by incubation with streptavidin-peroxidase conjugate and revealed using diaminobenzidine and H2O2. Sections were counterstained with Mayer's hemalum and mounted with coverslips in Eukitt medium (Vel, Leuven, Belgium).
Ki-67 antigen staining was carried out using MIB-1 monoclonal antibody (Dako) and the anti-mouse detection method described previously.
Computer-assisted quantification of immunostaining
Quantification of immunostaining was performed following the same methodology as previously published 7. Galectin and Ki-67 stainings were studied in 10 randomly selected areas of well-preserved bronchial epithelium, defined by the presence of basal and columnar cells without detachment from the basement membrane, on each section (three to 11 per subject), in both large and small airways. Leukocyte infiltration was evaluated in 10 randomly selected areas of epithelium, 10 areas of lamina propria and 10 areas of submucosal glands on each section. Large airways were defined as cartilaginous bronchi with submucosal glands and small airways as membranous bronchioles, without cartilage or glands and with an internal diameter <2 mm, as previously described elsewhere 21. Two to nine different airways per slide and a total of 10 to 21 large airways and nine to 28 small airways per subject were evaluated. A total of 30, 34 and 45 tissue sections were analysed in the nonsmokers, smokers and COPD groups, respectively.
Computer-assisted quantification of the staining in the selected areas was carried out using a SAMBA 2005 system (Alcatel TITN, Grenoble, France) equipped with a colour camera and an optical microscope (Olympus BX 50; Olympus Optica Company Ltd, Tokyo, Japan), using a final magnification of 400×. Results were expressed as the mean optical density (measured in arbitrary units), representing the mean intensity of staining in the considered area, and as the labelling index (measured as per cent), representing the percentage of surface staining with respect to the total area selected, therefore, reflecting relative numbers of positive cells.
Statistical methods
The staining indices in the different sections from each patient were subjected to statistical analysis. Two types of comparisons were performed. First, a mean value was calculated for each patient from the staining indices obtained in the different sections and comparisons between groups (COPD versus controls) were performed using the Mann–Whitney U-test. Secondly, within-group comparisons were performed on paired data (large versus small airways) obtained in the same section (where both large and small airways were present) using the Wilcoxon matched-pairs test. Correlations were tested using the nonparametric Kendall test or Spearman's method as appropriate. A p-value <0.05 was considered statistically significant. Results were expressed as medians and ranges.
RESULTS
Airway expression of galectin-1 and -3 in COPD and control patients
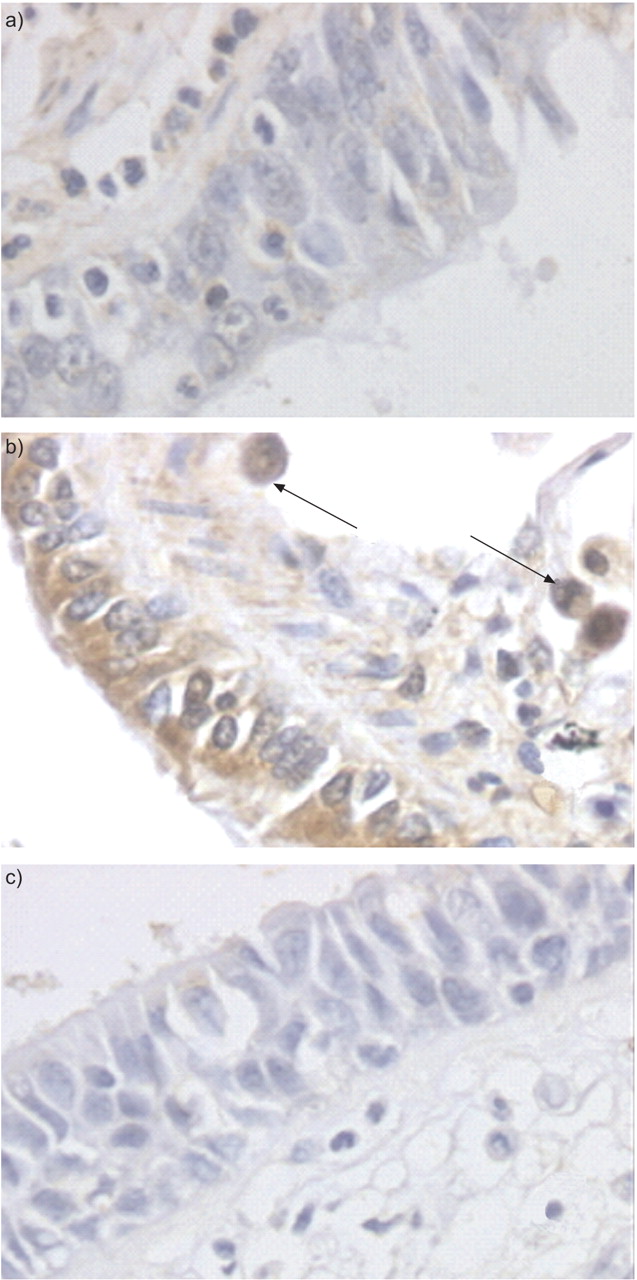
Galectin-1 expression was detected in the bronchial epithelium from control and COPD patients (fig. 1a⇓), as well as in the lamina propria notably in endothelial cells (data not shown). Galectin-3 immunostaining was observed in bronchial epithelial cells, especially in the cytoplasm of ciliated and sero-glandular cells (fig. 1b⇓), whereas staining was also observed in macrophages (fig. 1b⇓).

a) Galectin-1 and b) galectin-3 staining in the small airways from a chronic obstructive pulmonary disease patient and c) control staining with normal rabbit serum. Arrows represent galectin-3 expression observed in alveolar macrophages.
While no change in galectin expression was observed in the large airways, the bronchial epithelium of smokers exhibited increased galectin-1 expression in the small airways, as compared with nonsmokers (approximate three-fold increase; p<0.001; fig. 2⇓). In COPD, galectin-1 expression also tended to increase (p = 0.06, as compared with nonsmokers), but to a much lower extent than in smokers (fig. 2⇓).

Staining indices of galectin-1 and -3 expression in the bronchial epithelium. Epithelial expression of a) galectin-1 and b) galectin-3 in large and small airways from chronic obstructive pulmonary disease (COPD) patients versus nonsmokers (Nonsm) and smokers (Sm) is shown in terms of mean optical density (MOD). The horizontal bars represent the medians. au: arbitrary units. **: p<0.01; #: p = 0.06 (Mann–Whitney U-test).
In contrast, expression of galectin-3 in the small airway epithelium, which tended to increase in smokers (p = 0.06), was strongly increased in COPD (approximate two-fold increase; p<0.001; fig. 2⇑).
Ki-67 expressing epithelial cells in COPD and control airways
Numbers of Ki-67 expressing epithelial cells were significantly increased in small airways of patients with severe COPD, as compared with smokers (p = 0.05; fig. 3⇓). No significant changes in Ki-67 staining were observed in COPD large airways (data not shown).

Epithelial proliferation (Ki-67 staining) in the bronchial mucosa of chronic obstructive pulmonary disease (COPD) and control patients. Relative numbers (represented by the labelling index (LI)) of Ki-67 expressing epithelial cells are shown in the small airways of patients with severe COPD, nonsmokers (Nonsm) and smokers (Sm). The horizontal bars represent the medians. #: p = 0.05 (Mann–Whitney U-test).
Neutrophil, macrophage and T-cell infiltration of COPD airways
Highly significant increases in neutrophil numbers were observed in severe COPD as compared with controls (p<0.001; table 2⇓). When compared with smokers, neutrophils were elevated in submucosal glands and in small, but not large, airways (table 2⇓). Thus, a trend for increased neutrophil numbers was observed within the epithelium of large airways from smokers, as compared with nonsmokers (table 2⇓).
Leukocytes immunostaining data
CD68+ monocytes/macrophages were also observed in COPD airways occasionally clustered in foci (data not shown), whereas staining values largely overlapped with those observed in controls (table 2⇑). However, relative numbers of macrophages were significantly increased in small airways of smokers and COPD, as compared with nonsmokers (table 2⇑).
Lymphoid foci were observed around COPD airways (data not shown), with increased T-cell numbers in the epithelium and lamina propria of small airways, as compared with smokers (p = 0.003 and p<0.001, respectively; table 2⇑).
Relationships between galectins, epithelial proliferation, airway inflammation and lung function
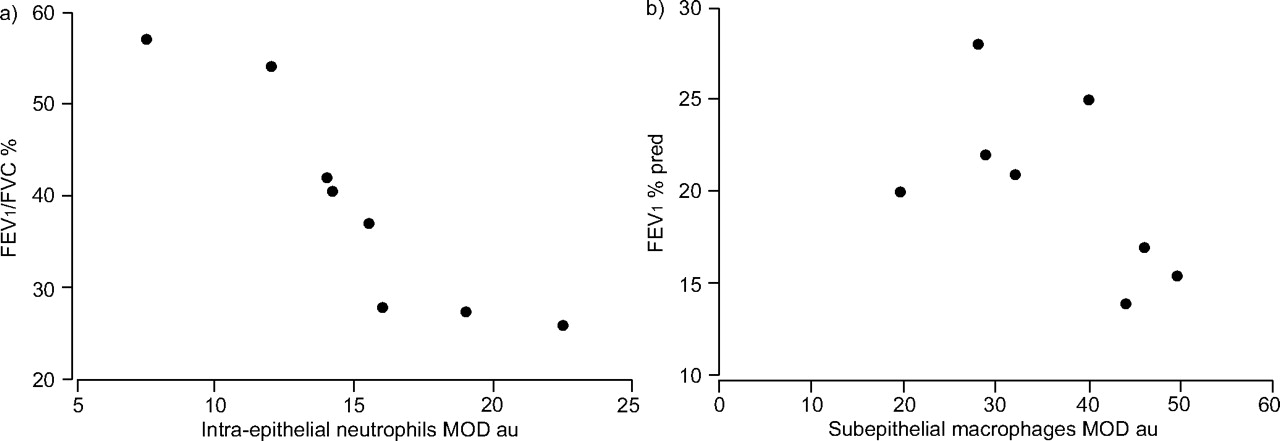
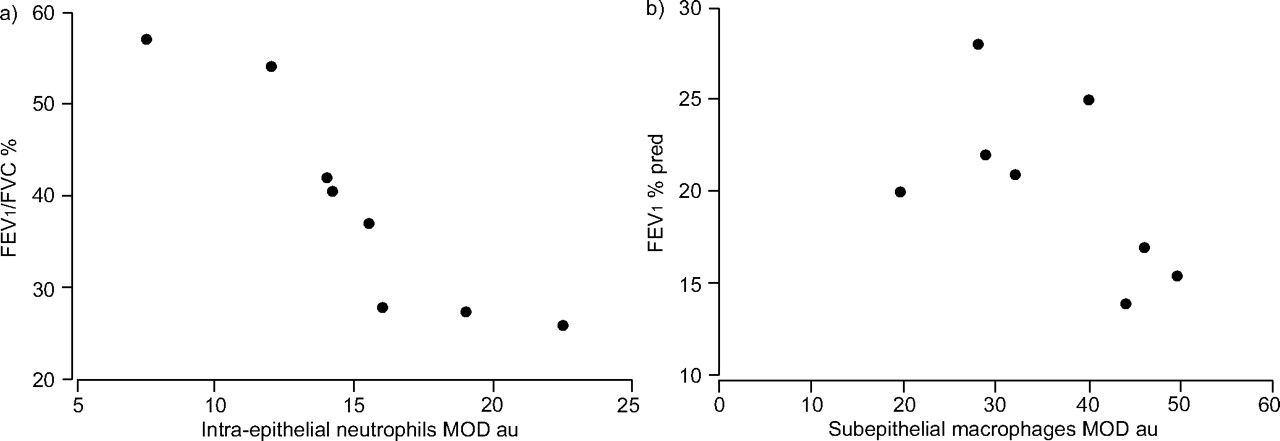
Epithelial galectin-3 expression in small airways did not correlate in COPD with epithelial proliferation or neutrophil infiltration. In the small airways, Ki-67+ cell numbers correlated with neutrophils (p = 0.03, r = 0.76; fig. 4⇓). Furthermore, intra-epithelial neutrophils correlated with airflow obstruction assessed by the forced expiratory volume in one second (FEV1)/forced vital capacity ratio (p = 0.001, K = -0.93) and subepithelial monocyte/macrophages correlated with FEV1 (fig. 5⇓).
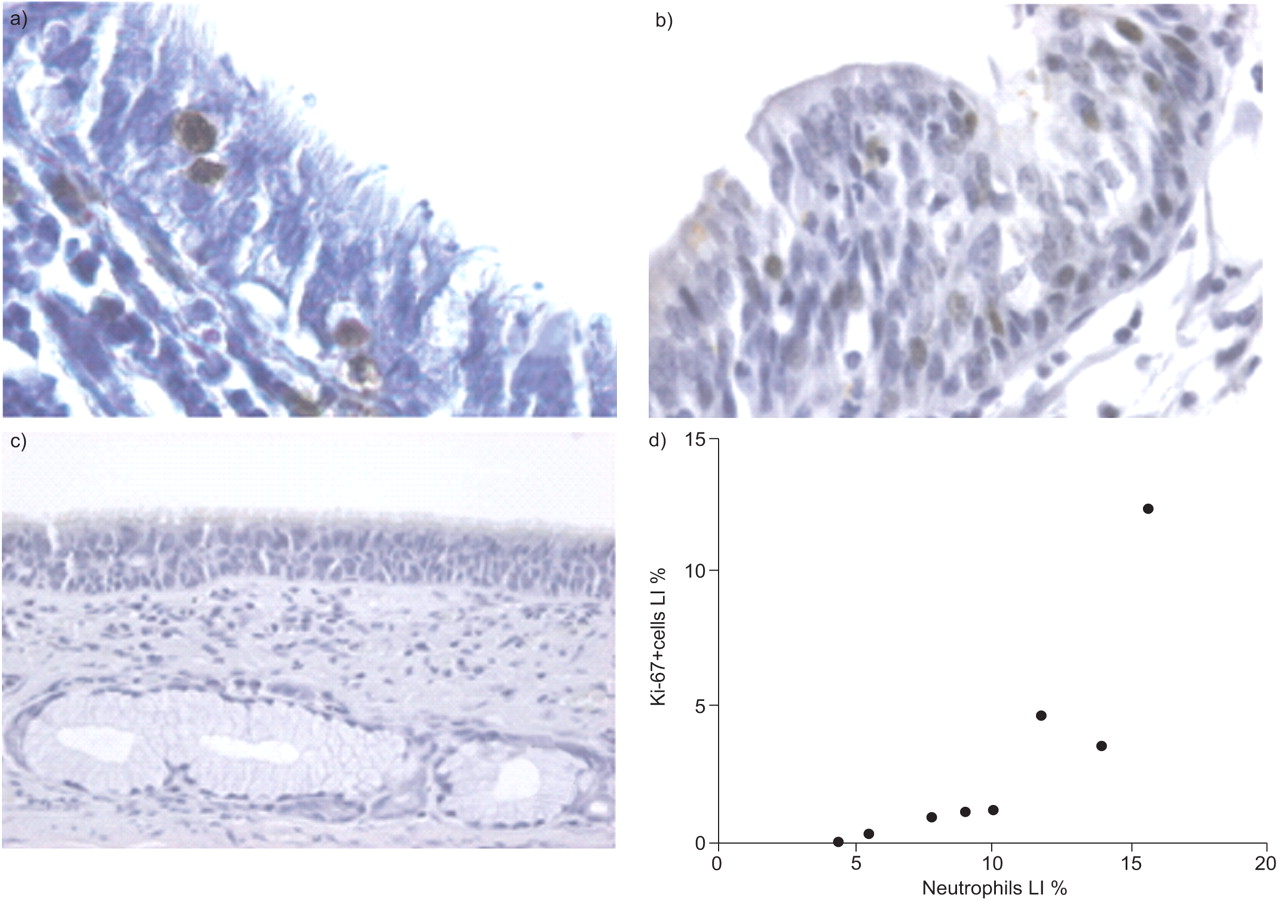
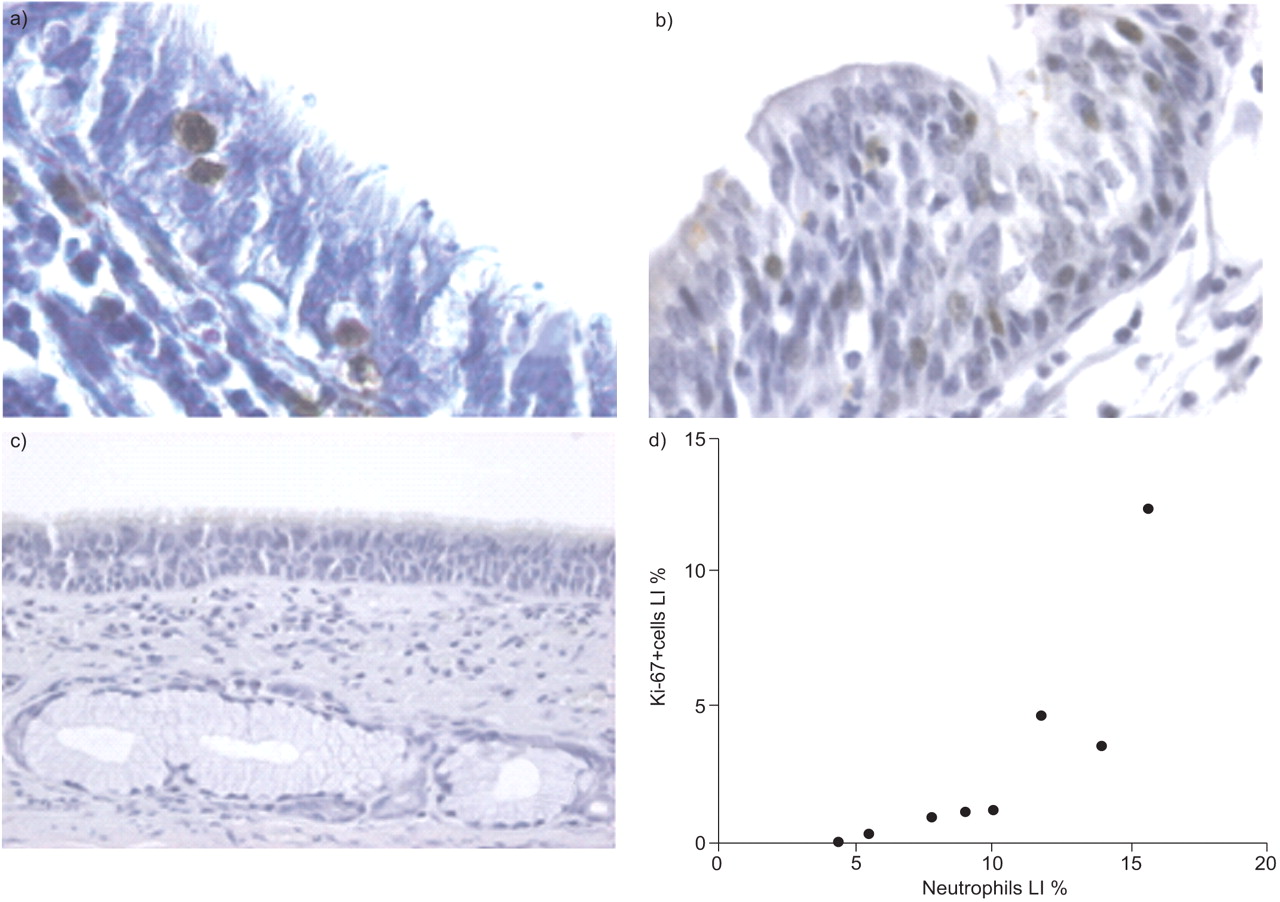
Relationship between a) neutrophils (brown-stained cells) and b) Ki-67+ epithelial cells (brown-stained cells) in small airways of severe chronic obstructive pulmonary disease (COPD) patients. c) A representative isotypic (mouse immunoglobulin G1κ) control. d) Neutrophils were plotted against Ki-67+ proliferating epithelial cells in the small airways of severe COPD patients according to the Spearman's method. LI: labelling index. p = 0.03, r = 0.76.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relationship between neutrophils, macrophages and lung function in the small airways of severe chronic obstructive pulmonary disease patients. Functional parameters of airway obstruction (forced expiratory volume in one second (FEV1) and FEV1/forced vital capacity (FVC) ratio) plotted against a) intra-epithelial neutrophils (p = 0.001, k = -0.93) and b) subepithelial macrophages (p = 0.03, k = -0.73) expressed as mean optical density (MOD) values. % pred: % predicted; au: arbitrary units. K (Kendall's tau) and p-values were calculated according to the Kendall test.
DISCUSSION
Galectins are secreted lectins that participate in the regulation of neutrophil recruitment and cell apoptosis/proliferation, notably during lung development and inflammatory responses 10–14. The present study showed for the first time that epithelial expression of galectin-3 is increased in the small airways of severe COPD patients. In contrast, increased galectin-1 in the small airways was observed in smokers without COPD. By assessing large numbers of tissue specimens containing various airway generations (whole explanted lungs and surgical specimens), which probably improved the power to detect between-group differences 22, it was also shown that in severe COPD neutrophils are preferentially micro-localised to the epithelium of the small airways, where they correlate with increased epithelial proliferation and airflow obstruction.
It has been shown that galectin-1 expression is induced in nasal polyps upon corticosteroid therapy and that this may limit tissue eosinophilia 23. Decreased galectin-1 and increased galectin-3 expression has been associated with defective apoptosis of synovial mononuclear cells in juvenile idiopathic polyarthritis 24. A recent study showed that galectin-3 is mitogenic for cardiac fibroblasts, induces collagen deposition and may lead to ventricular dysfunction 25. In the lung, epithelial expression of galectin-3 is increased in nonsmall lung cancer 26, 27, while in murine asthma models, gene therapy targeted to galectin-3 inhibits allergen-induced airway inflammation, including IL-5 expression, and epithelial mucous metaplasia 28–30. Secretion of galectin-3 is also upregulated in a rat model of radiation-induced lung fibrosis 31, 32.
Changes in galectin expression observed in the present study could be relevant to COPD pathogenesis, as galectins control apoptosis of epithelial and inflammatory cells as well as neutrophil and macrophage activation. Thus, galectin-3 participates in the recruitment of neutrophils and triggers their release of oxygen radicals and elastase 14, which are thought to mediate the lung destruction induced by cigarette smoking 33. However, the present study focused on the airways and not on the lung parenchyma, where an increased apoptotic rate of alveolar epithelial and endothelial cells could underlie the development of emphysema 34, including in end-stage COPD 35, 36. The altered galectin balance between galectin-1 and -3 in the small airways could favour the hyperplasia of the bronchial epithelium observed in COPD. Increased numbers of proliferating (expressing Ki-67) epithelial cells were observed in COPD small airways, as previously reported in squamous metaplastic areas of large bronchi 37. Elevated Ki-67 expression was also previously observed in the lung alveolar wall from severe COPD patients 38, whereas it remained unchanged from controls in another study 36. Interestingly, in the present study Ki-67+ epithelial cell numbers in small airways correlated with neutrophils, suggesting a link between inflammation and tissue remodelling. Accordingly, increased Ki-67+ epithelial cells following allergen challenge were related in asthma to airway eosinophilia 39. Alternatively, these correlations could be due to the presence of intra-epithelial inflammatory cells expressing Ki-67 antigen. Although this possibility cannot be excluded, Ki-67 staining could not be related to epithelial infiltration by neutrophils via examination of serial sections of COPD stained for Ki-67 and neutrophils (data not shown).
Smokers without COPD display increased galectin-1 expression in the bronchial epithelium of small airways in parallel to a trend for increase in galectin-3, suggesting that the impaired balance between galectin-1 and -3 favouring airway inflammation and remodelling is only observed in susceptible smokers who develop COPD. It is tempting to speculate that in nonsusceptible smokers galectin-1 could help to suppress signals of inflammatory and proliferative responses. However, the present study was not designed to address the mechanisms of these changes, which could relate to intrinsic changes of the COPD bronchial epithelium or to nonspecific factors associated with severe disease, such as hypoxia 40. Also a putative effect of the treatment of COPD cannot be ruled out, although it seems unlikely that inhaled corticosteroid treatment could account for the observed changes in galectin expression 23.
The present study did not document a correlation between epithelial galectin-3 expression and neutrophil accumulation in severe COPD patients. As previously demonstrated 41–45, neutrophil infiltration of the airways of severe COPD patients was observed. The present authors showed that neutrophils in severe COPD patients are more particularly localised in submucosal glands and the small airways, a latter feature in contrast with mild stage COPD 46–48. Smokers were characterised by neutrophil infiltration of proximal bronchi, particularly within the epithelium. As previously reported elsewhere 43, 45, macrophage numbers largely overlapped those observed in smokers, although in the present study a trend for increase was observed in the small airways.
Regarding T-cells, a nonsignificant difference between groups was observed in the large airways. This is in contrast with previous studies of mild-to-moderate COPD showing increased subepithelial CD3 42, 44, 45 and CD8 cells 44, 45 in large airways. Conversely, in a study in severe COPD by Di Stefano et al. 49, CD3 and CD8 cell numbers were decreased and inversely correlated with the degree of airflow limitation, suggesting that cigarette smoke-induced bronchial inflammation is downregulated as the disease progresses. Alternatively, severe COPD could represent a particular subgroup of COPD patients with a distinct immunological phenotype. However, the present findings confirm that T-cells infiltrate the small airways of severe COPD patients, as observed in COPD lung parenchyma 50 and around lung vessels 51, a fact that is in agreement with a previous study in severe COPD 52 and that had been reported previously in mild COPD cases 48. Furthermore, it is shown that a decrease in T-cell numbers characterises smokers without COPD as compared with nonsmokers. As previously suggested 45, this observation could relate to the selection of a subpopulation of smoking subjects with low CD3 cell counts who are more resistant to the effect of cigarette smoke.
Micro-localisation of neutrophils to smooth muscle has recently been reported in COPD 53 and related to local expression of CXC chemokines 54. Similarly, localisation of mast cells within peribronchial smooth muscle in asthma 55 has been associated with the release of CXCL10 by smooth muscle cells 56. This study shows that intra-epithelial, in contrast to subepithelial, micro-localisation of neutrophils in severe COPD correlates with airway obstruction. In addition to the production of galectin-3 reported in the present study, bronchial epithelial cells can express CXC chemokines and adhesion molecules that may facilitate the trafficking of neutrophils to the airway epithelium. Conversely, neutrophil-derived products, such as elastase, can contribute in COPD to locally activated epithelial cells and trigger transcriptional programmes leading to epithelial expression of pro-inflammatory mediators. The present authors observed that subepithelial macrophages also correlate with airflow limitation, suggesting that in COPD inflammatory cells have different functional implications within the various airway micro-compartments 57.
Although the small numbers of patients limit the conclusions, the present study provided further evidence of important changes occurring in the bronchial epithelium of patients with severe COPD. Upregulated galectin-3 expression, unbalanced by galectin-1, is observed in the small airways of smokers who have developed severe COPD, in contrast with smokers without COPD who tend to display the opposite expression profile. Although other factors appear involved, changes in galectin expression could contribute to epithelial hyperplasia and neutrophil accumulation in the airways of COPD patients. It is noteworthy that the emerging involvement of galectin-2 in T-cell regulation and the pathogenesis of myocardial infarction highlights the importance of further analysis of the galectin network in lung diseases 58–60.
The present study showed for the first time that in severe chronic obstructive pulmonary disease the small airway epithelium is a particular site of neutrophil infiltration, related to epithelial proliferation and airflow limitation. The findings support previous studies 61, which proposed an important role for small conducting airways in the pathophysiology of chronic obstructive pulmonary disease. It is suggested that chronic damage to the epithelium is associated in susceptible smokers with increased galectin-3 expression and that micro-localisation of neutrophils to the small airway epithelium is a particular and functionally relevant feature in severe chronic obstructive pulmonary disease.
Acknowledgments
The authors would like to thank P. Thurion (Laboratory of Histopathology, University of Louvain, Mont-Godinne, Belgium) for his excellent technical assistance and I. Salmon, C. Decaestecker and N. D'Haene (Laboratory of Histopathology, University of Brussels, Brussels, Belgium) for help with statistical analyses and staining quantification. The authors would also like to thank L. Wilcock (Imperial College London, London, UK) for language revision of the manuscript.
- Received June 25, 2005.
- Accepted January 3, 2007.
- © ERS Journals Ltd
References










