





Abstract
Patients with chronic obstructive pulmonary disease (COPD) show abnormal adaptations of skeletal muscle redox status after exercise training. Increased skeletal muscle oxidative stress in COPD patients may prompt mitochondrial dysfunction.
The present study explores the association between body composition and mitochondrial respiration in seven COPD patients with low body mass index (BMIL), eight COPD patients with normal body mass index (BMIN) and seven healthy controls. All of them underwent a vastus lateralis biopsy in which muscle structure, in vitro mitochondrial respiratory function, uncoupling protein 3 (UCP3) mRNA expression and glutathione levels in both isolated mitochondria and the whole muscle were determined.
Mitochondrial respiratory function (assessed by acceptor control ratio (ACR)) was impaired in BMIL (2.2±0.6) compared with both BMIN (5.3±1.3) and controls (8.2±1.3). ACR significantly correlated with arterial oxygen tensionand with muscle endurance but it showed a negative association with exercise-induced increase in blood lactate levels. UCP3 mRNA expression was reduced in BMIL patients.
In conclusion, chronic obstructive pulmonary disease patients with low body mass index show electron transport chain dysfunction, which may contribute to low muscle endurance in the current subgroup of patients.
Chronic obstructive pulmonary disease (COPD) 1 is associated with several extrapulmonary effects. Skeletal muscle dysfunction, one of the most extensively studied systemic effects, is a prominent contributor to exercise limitation 2. Limb muscle abnormalities exhibit a multifactorial nature, including physical inactivity, systemic inflammation/oxidative stress and cell hypoxia 3–5. Loss of muscle mass, which occurs in a subgroup of COPD patients 6, results in a deleterious impact on health status, increased use of healthcare resources and poor disease prognosis, which is partly independent of the degree of forced expiratory volume in one second (FEV1) impairment 7, 8. A better knowledge of the interplay among mechanisms determining limb muscle dysfunction and loss of fat-free mass (FFM) in COPD may enhance the understanding of the functional heterogeneities of the disease, which in turn may lead to a better management of patients.
In previous studies 9, 10, the present authors reported that although physical training improves muscle bioenergetics in COPD patients irrespective of the staging severity, abnormal adaptation to muscle oxidative stress is generated by repeated high-intensity training sessions. While in control subjects reduced glutathione (GSH) concentration is increased in skeletal muscle after 8-week endurance training, the opposite effect is observed in COPD patients, in whom increased oxidised glutathione (GSSG) levels were observed after training. These data suggest that antioxidant buffering did not fully adapt to the higher rate of reactive oxygen species (ROS) production, leaving the trained muscle more susceptible to oxidative stress. Altered adaptations of muscle redox status after 8-week endurance training were particularly evident in patients with low body mass index (BMIL) 9, 10. Interestingly, GSH and GSSG concentrations at baseline did not differ between COPD patients and control subjects 9. Nevertheless, since a fall in mitochondrial glutathione can lead to deleterious consequences for the cell, even in the presence of cytosol GSH 11, and because it represents 10–15% of the total GSH pool size, the regulation of mitochondrial GSH needs to be evaluated in isolated organelles.
It has been recently shown that uncoupling protein 3 (UCP3) levels are diminished in the skeletal muscle of patients with COPD 12, 13, which in turn return to normal levels after 8-weeks of endurance training 14. The exact physiological function of UCP3 is unknown. Although it is primarily linked to enhanced rest energy expenditure through mitochondrial respiratory chain uncoupling 15, it was hypothesised that the physiological role of UCP3 is to protect mitochondria against lipotoxicity in cases where fatty acid influx exceeds the capacity to oxidise them. Interestingly, activity of 3-hydroxyacyl-CoA dehydrogenase (HAD), an important enzyme involved in the oxidation of fatty acids, is reduced in the skeletal muscle of COPD patients 16. Furthermore, lipid peroxidation has been associated to mitochondrial dysfunction 17, 18. It is important to emphasise that in two of the aforementioned studies 12, 14 the FFM index (FFMi) was assessed and was found to be significantly lower in COPD patients compared with control subjects. With this in mind, it was of interest to analyse the relationship between UCP3 levels and body composition.
Unlike peripheral skeletal muscles, respiratory muscles show adaptive changes in response to the chronic mechanical load, i.e. increased mitochondrial density, increased oxidative capacity, increased density of interfibrillar capillaries and increased proportion of fibre type I 19, 20. Equally, mitochondrial electron transport chain function is enhanced in inspiratory muscles of patients with COPD 21. Therefore, different behaviour of the mitochondrial respiratory chain could be expected in peripheral and respiratory muscles.
It was hypothesised that mitochondrial function may be impaired in the peripheral muscles of COPD patients, especially in the subgroup of patients with BMIL. In the present study, the current authors assessed the ex vivo peripheral muscle mitochondrial function and its relationship with arterial oxygen tension (Pa,O2), whole muscle and mitochondrial GSH and GSSG concentrations and UCP3 regulation.
For this purpose, an open biopsy of the vastus lateralis was performed in eight COPD patients with normal body mass index (BMIN), seven COPD patients with BMIL (<21 kg·m−2) and seven healthy sedentary individuals. Muscle structure, in vitro muscle mitochondrial respiration, UCP3 mRNA expression and GSH and GSSG levels in both muscle homogenate and isolated mitochondria were examined 11.
METHODS
Study group
A total of 15 male, clinically stable COPD patients (table 1⇓) 22 took part in the study. All patients were currently receiving bronchodilators and inhaled corticosteroids. They were clinically stable at the time of the study, i.e. without episodes of exacerbation or oral steroid treatment in the previous 4 months. None presented significant comorbidities. Seven age-matched healthy sedentary subjects (all male) were included in the study as controls. Eight COPD patients were characterised as presenting BMIN (BMI ≥21 kg·m−2) and the remaining seven COPD patients were characterised as presenting BMIL (BMI <21 kg·m−2) 23. Selection procedures for inclusion in the study were: 1) clinical assessment; 2) pulmonary function testing (Master Screen; Jaeger, Würzburg, Germany) 24, 25; 3) chest radiograph film; and 4) general blood analysis. Characterisation of the subjects included: 1) body composition analysis by Bioimpedance (Quantum X; RJL Systems Instruments, Clinton Township, MI, USA); 2) incremental cycling exercise protocol (CardiO2 cycle; Medical Graphics Corporation, St. Paul, MN, USA) 23; 3) left quadriceps static (isometric) and dynamic (isokinetic) strength and endurance (Cybex 6000; Lumex Inc., Ronkokoma, NY, USA); and 4) muscle morphometry using immunohistochemical procedures 26. All participants were informed of any potential risks and discomfort associated with the experiment. Written informed consent was obtained in accordance with the Committee on Investigations Involving Human Subjects at the Hospital Clinic, Barcelona University (Barcelona, Spain), which approved the study.
Characteristics of the study groups
Muscle biopsy
An open muscle biopsy of the vastus lateralis (∼0.8 g) was performed and processed as follows: 1) ∼0.65 g of the muscle sample was added to Kreb’s buffer (pH 7.40) solution for immediate processing assays (homogenate and mitochondrial glutathione concentrations and mitochondrial respiration assays); 2) ∼0.15 g of the muscle sample was embedded in paraffin for immunohistochemical analysis; and 3) ∼0.1 g of the muscle sample was added to RNA stabilisation reagent (RNAlater®, Ambion Inc., Austin, TX, USA) and stored at -20°C for RNA extraction.
Lung function at rest, body composition and exercise testing
Lung function
Forced spirometry, lung volumes and the transfer factor of the lung for carbon monoxide (TL,CO) were measured (MasterScreen; Jaeger) and the results were expressed as a percentage of the reference values obtained in the present authors’ laboratory 24, 25. Pa,O2, carbon dioxide tension, pH and blood lactate were analysed on a blood gas analyser (Ciba Corning 800, Medfield, MA, USA).
Body composition
Body composition was estimated using single-frequency (50 kHz) bioelectrical impedance analysis (Quantum X; RJL Systems Instruments) while subjects were in a supine position. FFM was calculated from sex-specific regression equations 27. The FFMi was obtained by dividing FFM (in kg) by height (in m2).
Incremental exercise protocol
After the placement of an arterial catheter (Seldicath; Plastimed, Saint Leu la Forêt, France), subjects were installed on the cycle ergometer with the mouthpiece in place. Online calculations of whole body oxygen uptake (V′O2), carbon dioxide output, minute ventilation, respiratory exchange ratio and cardiac frequency were obtained. Arterial blood samples were taken every 3 min throughout the test for analysis of blood gases and lactate. Blood samples were kept on ice until analysis (Ciba Corning 800). After 3 min of unloaded pedalling, the work rate was increased by 5 or 10 W·min−1. All studies were performed using an electromagnetically braked cyclo-ergometer (CardiO2 cycle; Medical Graphics Corporation) with mechanical assistance to overcome the internal frictional resistance.
Muscle strength
Measurements of maximal voluntary contractions of the left quadriceps were performed during static (isometric) and dynamic (isokinetic) contractions against an isokinetic system (Cybex 6000; Lumex Inc.). After a previous muscle warm-up, the peak extension torque was evaluated at 60° of knee flexion and the best of five repetitions was used for the analysis. The isokinetic strength test consisted of five repetitions at the following different angular speeds: 60, 90, 120, 180 and 240°·s−1. The best peak torque and peak power were used for the analysis. An endurance test was also carried out in all patients. The test consisted of a set of 30 knee extensions at a speed of 90°·s−1. The total work realised during the repetitions was used to analyse quadricep endurance. To ensure the maximal muscle strength in each test, rest periods were introduced between them.
Muscle structure
The muscle was embedded in paraffin and 10-μm thick sections were cut, varying the inclination of the holder by 5° increments until the minimum fibre cross-sectional area (fCSA) was obtained, which was defined as truly transverse 28. Consecutive cross-sections were processed by immunohistochemical techniques using monoclonal antibodies directed against myosin heavy chain isoforms type I and type II (MHCs and MHCf clones; Biogenesis, Poole, UK). The fCSA, i.e. mean least diameter, and proportions of type I and II fibres were assessed using a light microscope (OLYMPUS Series BX50F3; Olympus Optical Co., Hamburg, Germany) coupled with an image-digitising camera (Pixera Studio, version 1.2; Pixera Corporation, Los Gatos, CA, USA) and a morphometry program 29. At least 100 fibres were measured from each biopsy 28. Fibre diameters 40–80 µm were considered normal 26, 30.
Mitochondrial respiration
Mitochondria were obtained by centrifugation through a Percoll density gradient 31. Enrichment and recovery of mitochondria were ascertained by the specific activity of succinic dehydrogenase. The rate of oxygen consumption was measured using a Clark-type oxygen electrode (Hansatech Instruments Ltd, King’s Lynn, UK) within 1 h of muscle biopsy 32. The respiratory function was analysed in a medium containing 225 mM sucrose, 5 mM MgCl2, 10 mM KH2PO4, 20 mM KCl, 10 mM Tris and 5 mM hydroxyethyl piperazine ethane sulphonic acid, which was maintained at 25°C and continuously stirred using an electromagnetic stirrer and a bar flea. The mitochondrial suspension was added to the reaction medium and adenosine diphosphate (ADP)-limited respiration (state 4) was initiated by the addition of succinic acid (13.5 mM) in the absence of ADP. State 3 respiration was determined upon the addition of ADP (0.63 mM), a phosphate acceptor. Oxidative phosphorylation efficiency was assessed by acceptor control ratio (ACR), dividing the slope of state 3 by the slope of state 4 11.
RNA isolation and UCP3 mRNA expression
Total RNA was extracted from muscle tissue samples treated with RNAlater® (Ambion Inc.) by using TRIzol Reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according to the manufacturer's protocol. The quality of RNA samples was evaluated by capillary electrophoresis using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). 1 μg of total RNA was reverse-transcribed to cDNA using the First Strand cDNA synthesis kit in a 20 μL reaction (Roche Diagnostics GmbH, Mannheim, Germany).
UCP3 mRNA expression was measured by real-time PCR (qPCR) using specific Assay-on-Demand Taqman Gene expression probes (Applied Biosystems, Foster City, CA, USA). The probe used for UCP3 spans the 1–2 exon junction amplifying both long and short variants of the gene. PCR reactions were performed with 1 μL of cDNA along with the Taqman PCR Universal Master Mix and the corresponding primers and probe. PCR reaction conditions were those recommended by the manufacturer. All sample and nontemplate control reactions were performed in duplicate in the ABI Prism 7900 Sequence Detection System (Applied Biosystems). The comparative threshold cycle (CT) method for relative quantification of gene expression 33 was used to calculate relative changes in mRNA abundance. The CT values were obtained for each target probe and normalised with the corresponding CT values for the internal control housekeeping gene β2-microglobulin. The mRNA quantity was expressed as arbitrary units (Au).
Assessment of muscle redox status
Fresh muscle samples were homogenised in a buffer containing 20 mM Tris, 0.25 M sucrose, 40 mM KCl, 2 mM ethylene glycol tetraacetic acid and 1 mg·mL−1 bovine serum albumin. A total volume of 500 μL was separated for glutathione concentration assessment and the rest of the sample was used for mitochondrial isolation according to Percoll centrifugation gradient 31.
Glutathione concentration assessment
The two molecular forms, GSH and GSSG, were obtained by high-performance liquid chromatography (HPLC) in both homogenate and mitochondrial fractions, as described previously in detail 34. HPLC apparatus equipped with a 3-aminopropyl column and an UV spectrophotometric detector (365 nm) was used.
Data analysis
Results are expressed as mean±sem. Multiple comparisons were performed using ANOVA. A Neuman–Keuls test was used as a post hoc test for contrast analysis. When normality test failed, a logarithmic transformation was applied to the data. Pearson (or Spearman for abnormally distributed variables) regression analysis was used to assess univariate correlations. A p-value <0.05 was considered statistically significant.
RESULTS
Anthropometric and functional characteristics of COPD patients (BMIN n = 8 and BMIL n = 7) and controls (n = 7) are indicated in table 1⇑. As expected, FFMi was significantly lower in BMIL (17.0±0.5 kg·m−2) than in both BMIN (21.4±0.9 kg·m−2) and controls (22.3±0.5 kg·m−2; p<0.001). None of the patients with BMIN showed abnormally low FFMi (<16 kg·m−2) 35. Patients with BMIN showed higher FEV1 and diffusing capacity of the lung for carbon monoxidethan those with reduced muscle mass. The distribution of patients, according to the Global Initiative for Chronic Obstructive Lung Disease 1 classification, was as follows: four in stage II (three BMIN and one BMIL), six in stage III (four BMIN and two BMIL) and five in stage IV (one BMIN and four BMIL). The two groups of patients showed a similar degree of mild arterial hypoxaemia, but exercise-induced hypoxaemia was slightly more pronounced in BMIL. Similarly, BMIL patients had lower exercise tolerance (V′O2 peak 11±1 mL·kg−1·min−1) than both BMIN (19±2 mL·kg−1·min−1) and controls (26±2 mL·kg−1·min−1; p<0.0001).
Quadricep muscle strength and endurance were also lower in the BMIL group compared with the other two groups (table 1⇑). Muscle fCSA was higher (24%) in controls than in COPD patients, with identical results observed between BMIN and BMIL groups. No statistically significant differences among groups were detected. Muscle strength normalised by FFM (strength/FFM ratio) showed no differences between all groups (data not shown). It is noteworthy that muscle endurance normalised by FFM (endurance/FFM ratio) was significantly lower in BMIL patients (16.7±3.5 J·kg−1) than in both BMIN patients (36.1±4.1 J·kg−1) and controls (35.1±3.6 J·kg−1; p<0.01).
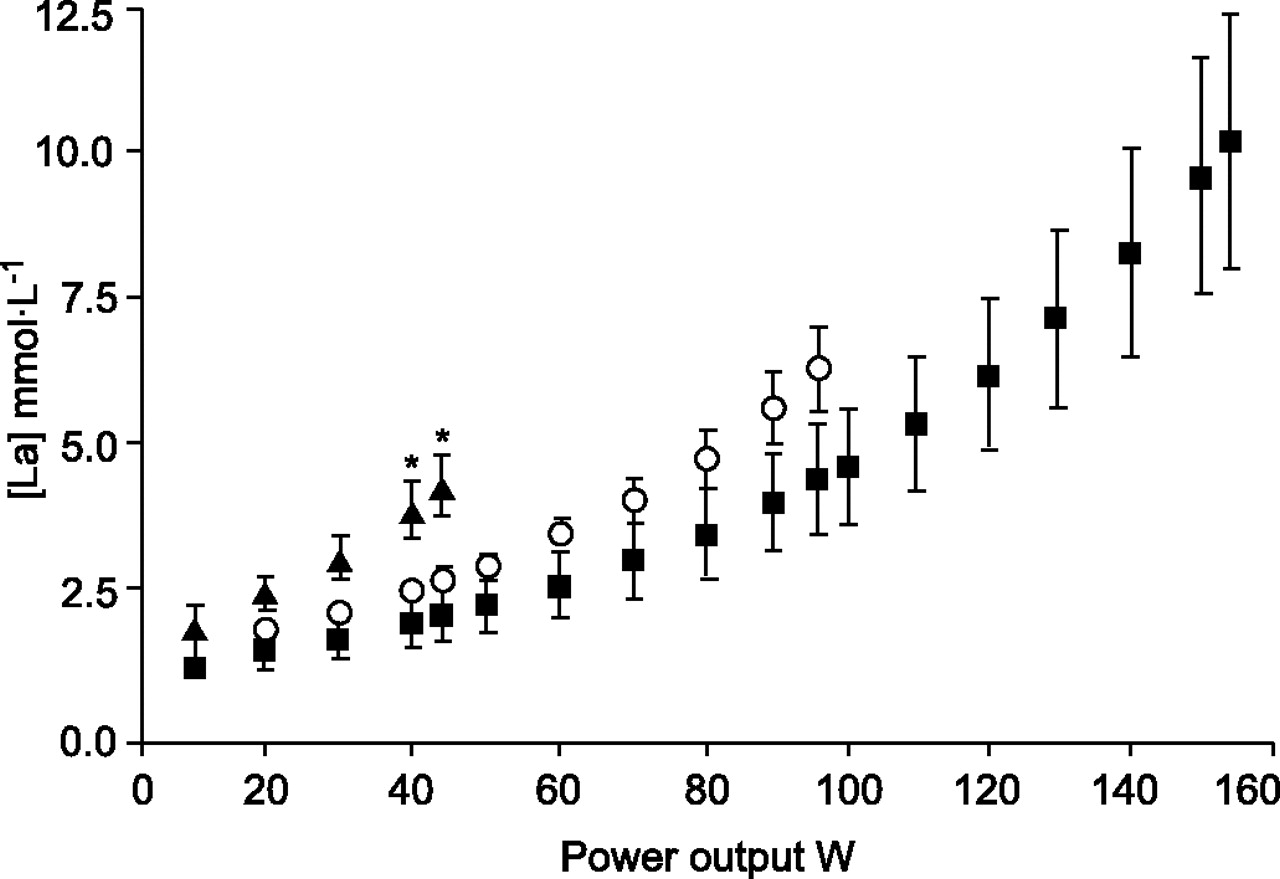
As shown in figure 1⇓, BMIL patients presented earlier arterial blood lactate release than both BMIN patients and controls. As expected, exercise-induced increases in blood lactate levels ([La]) showed a strong association with V′O2peak (r = 0.84, p<0.0001) and peak power output (r = 0.85, p<0.0001).

Mean arterial blood lactate levels ([La]) during incremental cycling exercise. The graph displays [La] in chronic obstructive pulmonary disease (COPD) patients with low body mass index (BMIL, ▴), COPD patients with normal body mass index (BMIN, ○) and healthy sedentary subjects (controls, ▪) during incremental cycling exercise. Early lactate release was seen in BMIL patients compared with both BMIN and controls. *: p<0.05.
Mitochondrial respiration
Individual data for ACR are illustrated in figure 2⇓. On average, ACR was significantly lower in BMIL patients (2.2±0.6) than in both BMIN patients (5.3±1.3) and in healthy sedentary subjects (8.2±1.3; p<0.01). State 3 of respiration using succinate-energised mitochondria was lower in BMIL patients (20.7±7.5 nmol·mg−1·min−1; p<0.01) compared with healthy controls (134.1±32.9 nmol·mg−1·min−1), but no statistical differences were seen when compared with BMIN patients (72.1±39.6 nmol·mg−1·min−1). No differences in state 4 of respiration were detected between the groups (BMIL 8.8±2.3 nmol·mg−1·min−1, BMIN 10.7±4.1 nmol·mg−1·min−1 and controls 17.4±3.4 nmol·mg−1·min−1). Mitochondrial metabolic state 3 significantly correlated with ACR (r = 0.68, p<0.001) and FFMi (r = 0.46, p<0.05).
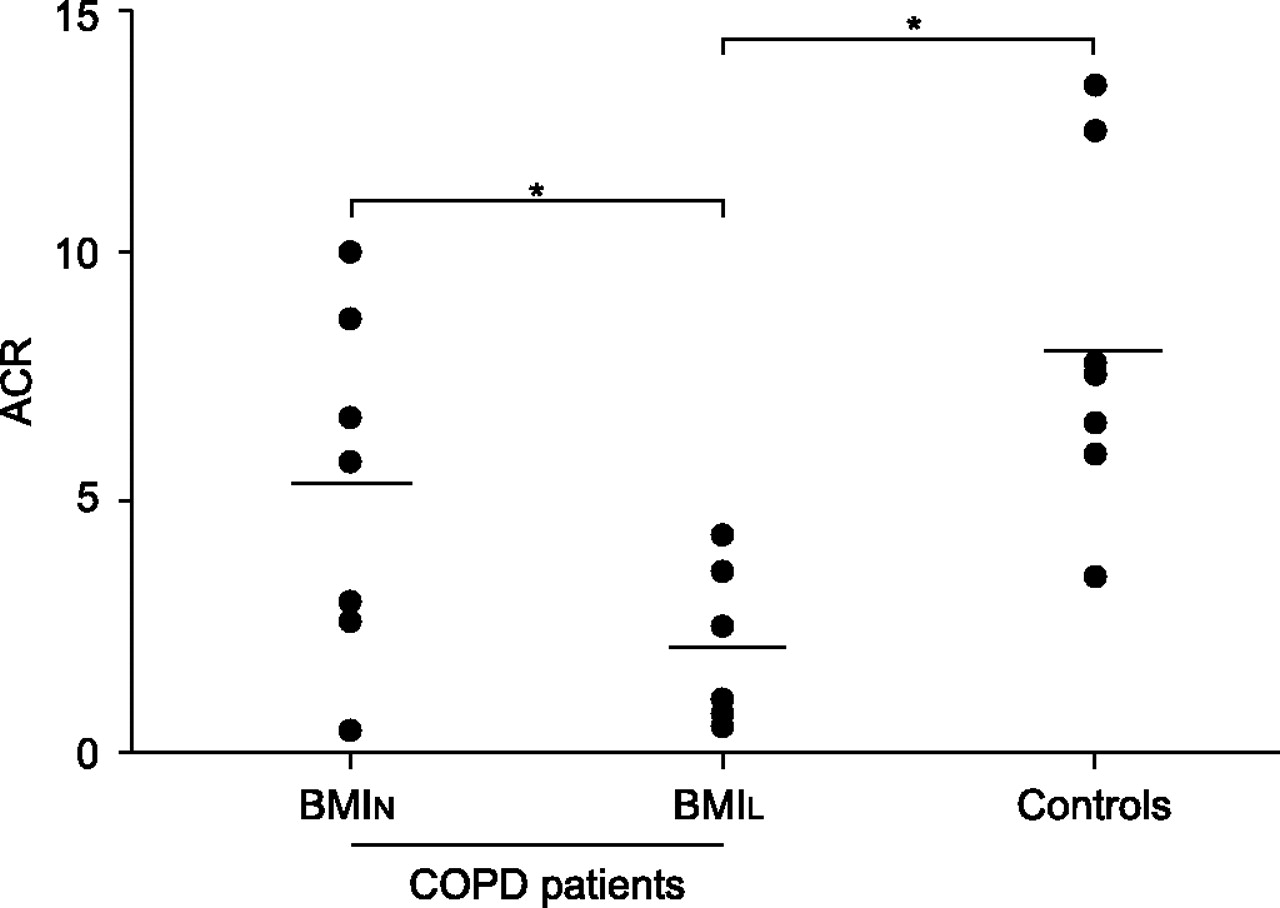
In vitro mitochondrial respiratory chain function. Individual (•) and mean group (––––) data for acceptor control ratio (ACR). Significant differences in ACR were shown between chronic obstructive pulmonary disease (COPD) patients with low body mass index (BMIL) and normal body mass index (BMIN) patients, and between BMIL patients and controls. Data from one COPD patient with BMIN are missing due to technical problems. *: p<0.05.
Pa,O2showed a significant correlation with ACR both at rest (r = 0.70, p = 0.0016) and at peak exercise (r = 0.63, p = 0.009; all subjects taken as a whole for calculations). Exercise-induced increase in [La] from rest to 45 W was negatively correlated with ACR (r = -0.60, p = 0.01). Furthermore, a positive correlation was shown between ACR and muscle endurance (r = 0.44, p<0.05).
Muscle UCP3 mRNA expression
Individual figures for qPCR UCP3 mRNA expression are displayed in figure 3⇓. On average, UCP3 mRNA expression was lower in BMIL patients (618.2±198.8 AU) compared with both BMIN patients (1,371±306.4 AU) and healthy controls (1,118.8±326.8 AU; p = 0.07). There was no association between UCP3 mRNA expression with any of the muscle measurements, either function or structure. However, a significant association was observed between UCP3 mRNA expression and FFMi (r = 0.58, p<0.01; fig. 4⇓).

Uncoupling protein 3 (UCP3) mRNA expression. Individual (•) and mean group (––––) data for UCP3 mRNA expression assessed by real-time PCR (qPCR) are presented. Low body mass index (BMIL) patients show lower values than both normal body mass index (BMIN) patients and controls. Data from one BMIN patient and one healthy subject are missing due to technical problems in qPCR assessment. *: p = 0.07.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relationship between uncoupling protein 3 (UCP3) mRNA expression and fat-free mass index (FFMi) for low body mass index (BMIL) patients (•), normal body mass index (BMIN) patients (○) and controls (□). A strong correlation is observed (r = 0.58, p<0.01), dashed lines represent 95% confidence interval of the regression. Data from one BMIN and one healthy subject are missing due to technical problems in UCP3 mRNA expression assessment by real-time polymerase chain reaction.
Glutathione assessment
No differences among groups were observed in muscle homogenate concentrations of GSH (BMIL 7.1±0.7, BMIN 8.9±2.3 and controls 5.3±1.2 nmol·mg−1) and GSSG. Similarly, no differences in mitochondrial GSH (BMIL 1.0±0.5, BMIN 1.5±0.4 and controls 1.0±0.1 nmol·mg−1) and GSSG were detected.
DISCUSSION
The current study provides original evidence of impaired mitochondrial oxidative phosphorylation, assessed as ACR, in succinate-energised mitochondria in COPD patients with reduced BMI (fig. 2⇑) this has not been observed in BMIN patients or in healthy subjects previously. Since no significant changes were detected in state 4 and state 3 was reduced, altered ACR in BMIL patients seems to be consistent with impaired electron flow from complex II–V and/or impaired adenosine triphosphate synthesis.
Although speculative, it can be hypothesised that altered oxidative phosphorylation in BMIL patients indicates an abnormal mitochondrial adaptation to long-term repeated episodes of cell hypoxia. The association between ACR and Pa,O2seems to support this contention. It is acknowledged, however, that causality cannot be established from the current study.
Relationships between poor arterial oxygenation and systemic manifestations of COPD, i.e. low FFMi and high tumour necrosis factor-α plasma levels, have been reported by other authors 36. Furthermore, abnormal mitochondrial respiratory chain function has been identified as a contributing mechanism to limb muscle dysfunction in patients with tissue hypoxia due to chronic peripheral arterial obstruction 37. In addition to potential perturbations of the main determinants of systemic oxygen flow, COPD patients are prone to present low muscle oxygen conductance 38 from capillaries to mitochondria and muscle oxygen supply and demand heterogeneities. These two factors may generate cell hypoxia during exercise, even in the absence of overt arterial hypoxaemia.
It is known that tissue hypoxia stabilises hypoxia-inducible factor 1-α 39, which mediates multiple cellular and systemic homeostatic responses to hypoxia 40 by actively downregulating mitochondrial oxygen consumption 41, 42.
Conversely, the lower UCP3 muscle levels in COPD patients may imply a lower ability to prevent fatty acids oxidation 43, which in turn might prompt mitochondrial abnormalities 17, 18.
Regardless of the potential mechanisms leading to a fall in oxidative phosphorylation in BMIL patients, this has deleterious consequences on adequate muscle performance, probably conditioning an early lactate release and interfering with the sustainability of exercise. The latter seems to be supported by the association between ACR and exercise-induced increase in [La] and muscle endurance. This is the first time, to the present authors’ knowledge, that peripheral muscle mitochondrial respiration dysfunction is evidenced in COPD patients with BMIL and, thus outlining a potential association between the latter and the early lactate release during exercise (fig. 1⇑) and the impairment in muscle endurance.
One intriguing finding of the present study was that functional abnormalities (e.g. ACR) were not accompanied by substantial alterations in muscle structure. It is understood that the analysis of muscle structure–function relationships requires further attention due to several confounding factors, i.e. physical de-conditioning and geographical and ethnic differences that may partly explain the heterogeneity of the results reported in the literature 44, 45.
Muscle UCP3 mRNA expression
Vastus lateralis of BMIL patients are characterised by reduced levels of UCP3 mRNA expression compared with both BMIN patients and healthy controls (fig. 3⇑). Reduced skeletal muscle UCP3 protein levels 12, 14 and mRNA expression 13 have been previously reported for COPD patients. It should be emphasised that in two of the aforementioned studies in which FFMi was measured, the values of this magnitude were significantly lower in COPD patients compared with controls, a fact that in the present study applies only in the case of patients with BMIL. Similarly, a significant correlation between UCP3 mRNA expression and FFMi has been found in the present study (fig. 4⇑).
Although the exact function of UCP3 is unknown, there is compelling evidence that UCP3 serves as a carrier to move fatty acid anions out of the mitochondrial matrix 46, 47, thereby protecting fatty acids from ROS-induced oxidative damage. Interestingly, the activity of HAD, an enzyme involved in the oxidation of fatty acids, is reduced in the skeletal muscle of patients with COPD 16. Gosker et al. 14 reported that exercise training restores UCP3 content in limb muscles of patients with COPD, especially in those that fail to increase HAD activity, a factor that supports the hypothesis that UCP3 may protect against fatty acid oxidation. In all, the lower UCP3 muscle levels in COPD patients may account for a reduced ability to prevent fatty acids oxidation 43.
Glutathione system
As described in previous reports 9, 10, no differences in glutathione concentration were observed in whole vastus lateralis homogenates between BMIN patients, BMIL patients and healthy controls at rest. Similarly, no differences in mitochondrial glutathione concentration were detected between groups. This is in contrast to other disorders 11, in which a decrease in mitochondrial glutathione concentration may occur even in the absence of differences in muscle homogenate, with deleterious consequences for the tissue. Thus, in the current study, analysis of the GSH system in isolated mitochondria did not generate information additional to the measurements performed in muscle homogenates. This may be a relevant methodological finding with the potential to simplify further studies in the field.
Limitations of the study
The rather invasive approach of the study accounts for the small sample size, which may constitute a limitation for some of the areas explored. Patients with COPD may present abnormal fat to lean body mass ratio, such that BMI did not properly indicate patients’ muscle mass. This fact does not seem to alter the interpretation of the current results, since none of the COPD patients with BMIN presented abnormally low FFMi. Furthermore, FFM and FFMi (p<0.01 and p<0.001, respectively) were significantly different when comparing BMIL with both BMIN and controls. Patients with BMIL showed a more advanced stage of the disease than those with BMIN. It is well known that BMIL and severe FEV1 impairment are not fully independent, and that their association is linked to poor disease prognosis.
Conversely, the UCP3 protein content was not analysed due to limitations in the availability of tissue sample. Nevertheless, the present UCP3 mRNA expression data are in agreement with previously published results 12, 13.
Conclusion
The current investigation clearly identifies mitochondrial dysfunction in chronic obstructive pulmonary disease patients with low muscle mass. This abnormality is associated with altered muscle performance in terms of early lactate release and reduced muscle endurance. A reduced expression of uncoupling protein 3 mRNA is also present in low body mass index patients, which may indicate compromised protection against lipotoxicity. Whether these phenomena occur only in a subset of susceptible advanced chronic obstructive pulmonary disease patients or if it is a general feature of the final stages of the disease needs to be elucidated. Further research is required in order to explore the underlying mechanisms that are likely to be shared by other chronic conditions.
Acknowledgments
The authors would like to thank F. Burgos, C. Gistau, J.L. Valera (servei de Pneumologia (ICT), Barcelona, Spain) and all the technical staff of the Lung Function Laboratory for their skillful support during the study. E. Gimeno, N. Sánchez, C. Hernando and I. Díaz from EUIF Blanquerna (Ramon Llull University, Barcelona, Spain) are acknowledged for their outstanding work supervising the training sessions. The authors would also like to thank A. Russell (Clinique Romande de Réadaption SUVA Care, Sion, Switzerland) for his advice on the UCP3 expression analysis, M.C. Carmona (University of Barcelona, Barcelona, Spain) for critical and helpful discussion and B. González for her excellent technical assistance.
- Received June 30, 2006.
- Accepted December 10, 2006.
- © ERS Journals Ltd
References










