





Abstract
Glucocorticoids stimulate foetal surfactant synthesis. Therefore, they are used in impending pre-term birth. One mechanism of action on surfactant synthesis is through the induction of neuregulin (NRG) secretion by foetal lung fibroblasts. The direct effects on signalling pathways, and specifically on erbB receptors in foetal type II cell surfactant synthesis, are less well understood.
The present authors studied the effect of known promoters of foetal surfactant synthesis (namely dexamethasone and mature (i.e. NRG-containing) fibroblast-conditioned medium (FCM)) on erbB receptor activation, protein content and dimerisation patterns in foetal mouse lung type II cells.
Dexamethasone inhibited surfactant synthesis in immature type II cells at day (d)16 of gestation, while the mature FCM had stimulatory effects. Both treatments directly stimulated surfactant synthesis in more mature (d17) cells. At this gestational day, dexamethasone had only a small effect on phosphorylation, but it stimulated the protein levels of all four erbB receptors. Dexamethasone effects were distinct from those of mature FCM, which stimulated both protein content and phosphorylation of all erbB receptors and of the signalling intermediate phospholipase Cγ. Dexamethasone modulated erbB receptor dimerisation patterns, such that erbB2 became the main dimerisation partner for erbB4.
In conclusion, dexamethasone signalling involves erbB receptors in foetal type II cells, in a manner similar to, but distinct from, neuregulin-containing fibroblast-conditioned medium signalling.
Antenatal surfactant production by foetal lung type II epithelial cells is crucial for the perinatal transition from the aquatic intrauterine environment to the gaseous atmosphere. Insufficient surfactant production, a condition present mainly in pre-term infants, leads to respiratory distress syndrome (RDS) 1. Antenatal glucocorticoids decrease the incidence of RDS by ∼50%, but they do not change the incidence of bronchopulmonary dysplasia (BPD), a chronic lung disease affecting surviving pre-term infants. Most BPD infants had RDS 2, 3. The pre-natal administration of glucocorticoids has both stimulatory and inhibitory effects on the developing lung 4.
Mesenchyme–epithelial interactions play an important role in foetal lung development by controlling the appropriate initiation of foetal surfactant production. This cell–cell communication process was described >25 yrs ago and is stimulated by glucocorticoids in the immature lung 5. However, there remains a lack of information on the exact signalling pathways and factors involved in both the fibroblast–type II cell communication process and how it is stimulated by glucocorticoids. A better understanding of these mechanisms is crucial for developing therapeutic strategies that will promote the optimal maturation of the foetal surfactant system while avoiding the deleterious side effects of glucocorticoid treatment.
The present authors recently reported that neuregulin (NRG), a ligand of the erbB3 and erbB4 receptors, is part of the fibroblast–type II cell signalling process that initiates type II cell maturation 6. Mature foetal lung fibroblasts in culture secrete NRG into the conditioned medium (fibroblast-conditioned medium (FCM)), and this NRG secretion is induced by glucocorticoids 6. Since the erbB receptors, especially erbB2, are involved in the stimulatory effect of dexamethasone on foetal type II cells, and NRG signals the endogenous onset of foetal surfactant synthesis in foetal type II cells, the present authors chose to study how dexamethasone and NRG-containing FCM affect erbB receptor signalling in foetal type II cells. It was hypothesised that even though glucocorticoids stimulate the endogenous maturation of the fibroblast, there are distinct and important differences between the endogenous and the glucocorticoid-induced effects on the mechanisms controlling development of foetal type II cell surfactant production. These differences were studied in primary cultures of foetal mouse type II cells at day (d)16 and d17 of gestation, and in an immortalised mouse lung type II cell line.
MATERIALS AND METHODS
Reagents were purchased as follows: [3H]-choline (specific activity 86 Ci·mmol-1) from Perkin Elmer Life Sciences (Boston, MA, USA); silica gel H chromatography sheets from Analtech (Newark, DE, USA); protein-A-Sepharose CL-4B from Sigma-Aldrich (St Louis, MO, USA); recombinant horseradish peroxidase-linked anti-phosphotyrosine antibody RC2O from Transduction Laboratories (Lexington, KY, USA); epidermal growth factor receptor (EGFR), erbB4 and phospholipase (PL)Cγ antibodies from Santa Cruz (Santa Cruz, CA, USA); and erbB2, erbB3, and monoclonal NRG1 antibodies (Ab-2 for the biological blockade of NRG1 bioactivity) from Neomarkers (Fremont, CA, USA). The recombinant epidermal growth factor (EGF)-like domain of NRG1β (a gift from K. Carraway 3rd; UC Davis, CA, USA) was expressed and purified as previously described 7, 8. Mouse lung epithelial (MLE) 12 cells were purchased from the American Type Culture Collection (Manassas, VA, USA).
Experimental design
Isolated foetal d16 and d17 mouse type II cells and MLE 12 cells were used to compare the effects of dexamethasone and mature, NRG-containing FCM (both of which stimulate surfactant synthesis) on erbB receptor protein content, activation (tyrosine phosphorylation), dimerisation patterns, and activation of the downstream signal transduction intermediate PLCγ.
Preparation of mature FCM from d18 foetal lung fibroblasts
The animal research protocol was approved by the institutional animal care and use committee. Animals were maintained in accordance with National Institutes of Health Guidelines and the Helsinki Conventions for the use and care of animals. Foetal lung fibroblast cultures were prepared as described previously 9. FCM was collected from confluent cells following a 24-h serum starvation and stored at 4°C until used. Only FCM from d18 female lungs was used, because of the well-described male delay in the timing of lung maturation, including the stimulation by FCM of type II cell surfactant synthesis 10, 11. FCM from d18 female lung fibroblasts is well-documented to stimulate type II cell surfactant synthesis 11 and to contain NRG as an active component 6.
Preparation of primary foetal type II epithelial cells
Type II cell isolation was continued after the first differential adherence for fibroblast cultures, as previously described 6. Purity and histochemical characterisation of the type II epithelial cell cultures isolated with this method were confirmed by methods of Post and Smith 12 and determined to be >92%, as previously published 13. This characterisation indicates that these cells retain morphological features of type II cells, including the presence of surfactant lamellar bodies. This indicates that these cells provide a good system for studies of biochemical events occurring during specific stages of lung development.
[3H]-choline incorporation into disaturated phosphatidylcholine
The stimulation of surfactant synthesis was determined by measuring the incorporation of [3H]-choline into disaturated phosphatidylcholine (DSPC) as previously described 6, 11. All experiments included FCM with known surfactant synthesis stimulatory activity as a positive control.
Immunoblotting of erbB receptors in foetal type II cells
Foetal d17 type II cells were grown in six-well plates in Dulbecco’s Modified Eagle Medium (DMEM) with 20% charcoal-stripped foetal calf serum (FCS) until 80–90% confluence. After a 24-h serum starvation, either FCM (with or without added NRG blocking antibody or a nonspecific immunoglobulin (Ig)G antibody, 10 μg·mL-1 each) or dexamethasone (10-8 M) was added for a further 24 h. Control cells were left in serum-free medium for the same time period. Cells were lysed and protein extracted. Aliquots of 20 μg of total protein per lane were separated by electrophoresis and detected by Western blot, as previously detailed 6. Blots were re-probed for the housekeeping gene β-actin as an internal standard to control for possible loading differences.
Co-immunoprecipitation of the erbB receptors in MLE 12 cells
MLE 12 cells, rather than primary foetal type II cells, were used for erbB receptor co-immunoprecipitation (IP) experiments because of the need for larger amounts of protein for co-IP work. The present authors have shown that the effects of foetal FCM and NRG on DSPC synthesis in MLE 12 cells are similar to those seen in primary cultures of d18 foetal type II cells 6, and others have shown that MLE 12 cells exhibit characteristics of foetal type II cells, including the expression of surfactant proteins SP-B and SP-C and formation of microvilli and multivesicular bodies 14. MLE 12 cells express all four erbB receptors, as do foetal type II cells; however, erbB3 shows a higher level of constitutive phosphorylation in MLE 12 cells than it does in cultured foetal d18 type II cells 6.
Immunoprecipitation experiments in MLE 12 cells were carried out using a co-IP protocol to study the patterns of erbB receptor heterodimerisation in response to dexamethasone and mature FCM. Cell proteins were immunoprecipitated under low-stringency conditions with antibodies to each erbB receptor in order to lean about receptor–receptor interactions. MLE 12 cells were plated at a density of 2×106 cells in 100 mm dishes and grown in DMEM with 10% FCS until 80–90% confluent. Cells were serum-starved in either DMEM (controls) or DMEM containing 10-8 M dexamethasone for 24 h. Cells were then: stimulated with either EGF (100 ng·mL-1), NRG (purified recombinant EGF-like domain of NRG1β 7, 8; 33 nM) or DMEM (controls) for 2 min at 37°C; washed three times with ice-cold PBS; and lysed in co-IP buffer (20 mM Tris (pH 7.4); 150 mM NaCl; 1 mM MgCl2; 0,1% Triton X-100; 10% glycerol; 1 mM Na3VO4; 1 mM NaF; 1 mM ZnCl2; 10 mM β-glycerolphosphate; 5 mM tetrasodium pyrophosphate; 1 mM phenylmethylsulphonylfluoride; and 0.4 μg·mL-1 each of aprotinin, leupeptin, and pepstatin), as described previously 15. For each IP, 500 μg of total protein was used. IP protocols were followed as published 6.
Data analysis
The effects of treatment on choline incorporation and the amount of erbB receptor protein (measured by densitometry and normalised to actin) were expressed as a percentage of their intra-experimental control values. FCM-blocking values were compared to their specific intra-experimental FCM-stimulated values. A two-tailed unpaired t-test, followed by a post hoc Bonferroni correction, where appropriate, was used to test for statistical significance. Data are presented as mean±sem, unless otherwise stated.
RESULTS
[3H]-choline incorporation into DSPC in primary foetal type II cells
In d16 immature foetal type II epithelial cells, dexamethasone treatment inhibited surfactant DSPC synthesis to 53±4.8% (n = 8; p = 0.0006) of the level seen in controls (100±8.1%; n = 7). Mature FCM, which the present authors have shown contains NRG 6, had the opposite effect. The mature FCM stimulated [3H]-choline incorporation into surfactant DSPC in these immature foetal d16 type II epithelial cells to 152±9.8% (n = 7; p = 0.0017) of that seen in control cells (fig. 1a⇓). To ensure that the stimulation of DSPC synthesis by the mature FCM was a result of the presence of NRG in the FCM 6, an inhibitory NRG antibody was added to the FCM. The presence of the NRG-inhibitory antibody blocked the FCM-induced stimulation of surfactant synthesis in the d16 type II cells (103±9.9%; n = 7; p = 0.0048). This result is similar to the specific inhibitory effect of this antibody in mature FCM added to mature d18 foetal lung type II cells, as described previously by the current authors 6.

Effect of mature fibroblast-conditioned medium (FCM) and dexamethasone on surfactant disaturated phosphatidylcholine (DSPC) synthesis in foetal type II epithelial cells at a) day 16 and b) day 17 of gestation. Data are presented as mean±sem. n = 3–6. □: control; ░: FCM; ▓: FCM + inhibitory antibody to neuregulin; ▒: dexamethasone. #: p<0.0125 after post hoc Bonferroni correction.
In d17 foetal type II epithelial cells, dexamethasone and mature FCM stimulated [3H]-choline incorporation to 136±19.9% (n = 6; p = 0.008) and 151±22.3% (n = 5; p = 0.009), respectively, as compared to experimental-specific controls (100±4.9%; n = 6; fig. 1b⇑). Again, the stimulatory effect of the mature FCM in d17 type II cells was blocked by the addition of inhibitory NRG1 antibody to the FCM (98±12.7%; n = 6), indicating the stimulatory effect of the FCM was related to the presence of NRG.
ErbB receptor protein expression and activation in primary foetal type II cells
Because both dexamethasone and mature FCM had similar stimulatory effects on DSPC synthesis in d17 type II cells, their effect on erbB receptor expression was investigated in these cells only. Mature FCM strongly stimulated endogenous tyrosine phosphorylation of erbB1 (EGFR), erbB2 and erbB4, without need for an additional stimulatory ligand (fig. 2a⇓, lane 2). To determine the specific role of NRG on this receptor-stimulatory activity, an inhibitory NRG1 antibody was added to the mature FCM. The antibody inhibited the stimulatory effect of the FCM on phosphorylation of these three erbB receptor proteins in the d17 type II cells (fig. 2⇓, lane 3). Dexamethasone, on the other hand, had no stimulatory effect on the endogenous phosphorylation of any of these proteins (fig. 2⇓, lane 5).

Effect of mature fibroblast-conditioned medium (FCM) and dexamethasone on expression and activation of erbB receptors and the signalling intermediate phospholipase (PL)Cγ in foetal type II epithelial cells at day (d)17 of gestation. a) Cell lysates were probed with an antibody reacting with phosphorylated tyrosine residues (αptyr) to evaluate activation of these proteins. b) Blots were stripped and re-probed with antibodies for the specific erbB receptors (αerbB1, αerbB2, αerbB3, αerbB4) and PLCγ (αPLCγ) proteins. All blots were re-probed with actin antibody as an internal standard. The erbB receptor sizes range from 170 kDa (erbB1) to 190 kDa (erbB4). PLCγ migrates at 150 kDa. Lane 1: lysates from untreated d17 type II cells (controls); lane 2: lysates from cells exposed to mature FCM for 24 h; lane 3: lysates from cells exposed to mature FCM plus an inhibitory antibody to neuregulin (NRG)1 (αNRG); lane 4: lysates from cells exposed to mature FCM plus a nonspecific immunoglobulin (Ig)G antibody (αIgG); lane 5: lysates from cells exposed to 10-8 M dexamethasone for 24 h; lane 6: protein standard.
Mature FCM stimulated the protein content of all erbB receptors (fig. 2b⇑, lane 2). Once again, the stimulatory effect of mature FCM was blocked by addition of inhibitory NRG1 antibody (fig. 2⇑, lane 3). This inhibition was not seen with the addition of a nonspecific IgG antibody (fig. 2⇑, lane 4). Exposure to dexamethasone for 24 h also stimulated the protein levels of all four receptors. This effect was most prominent for erbB2, erbB3 and erbB4 (fig. 2⇑, lane 5); it was less prominent for erbB1.
An erbB receptor-induced increase in protein content and phosphorylation of the intracellular signalling protein PLCγ after mature FCM treatment was also found. This increase was also blocked by the inhibitory NRG antibody, leading to the suggestion that one of the signalling pathways initiated by mature FCM in type II cell surfactant synthesis involves PLCγ. Interestingly, dexamethasone did not significantly increase the protein content or phosphorylation of PLCγ.
ErbB receptor protein expression and NRG-induced phosphorylation in MLE 12 cells
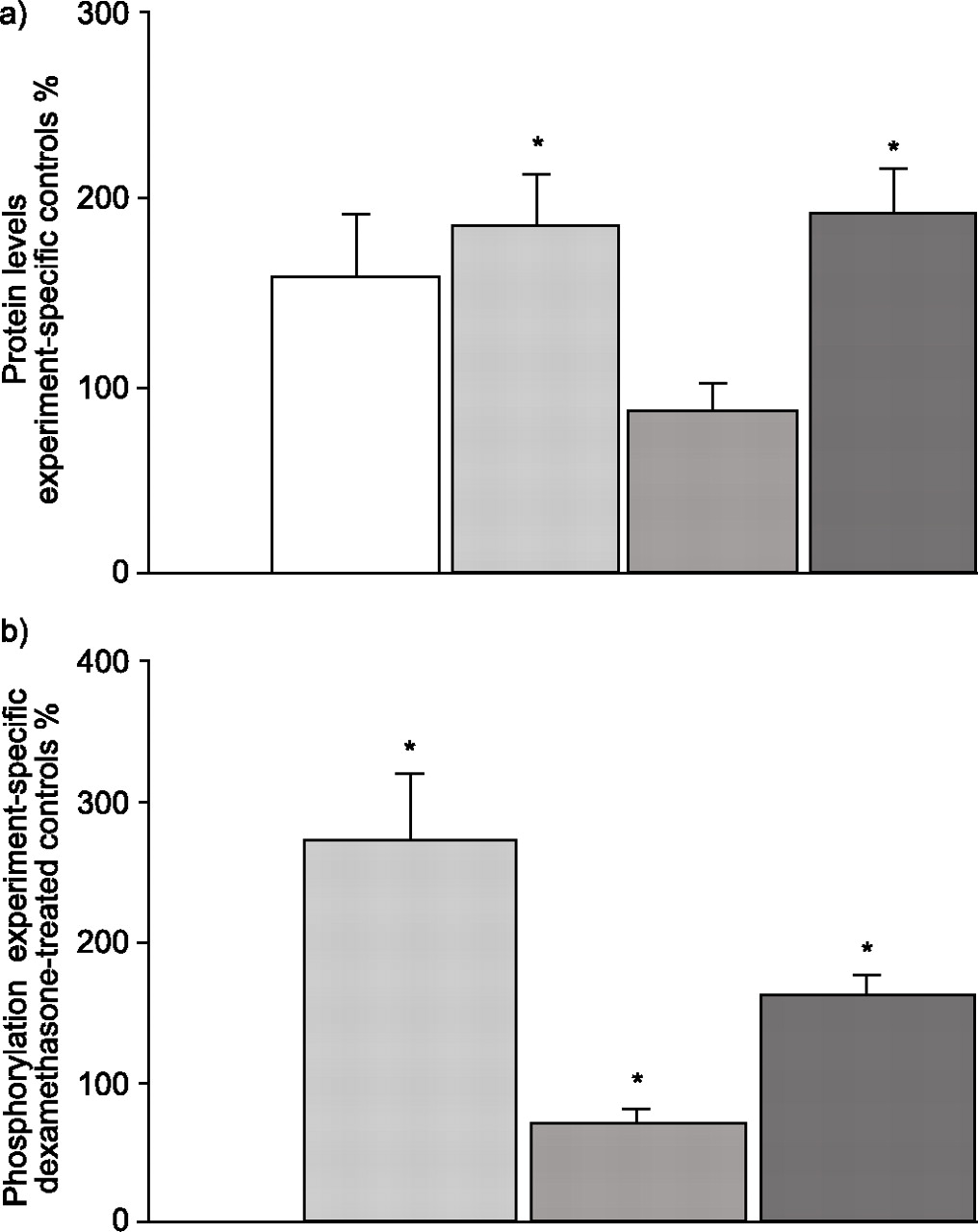
The foregoing studies showed effects of dexamethasone and NRG on erbB receptor protein content. To gain further insight into the effect on protein amount and on NRG-induced erbB receptor phosphorylation, immunoprecipitations for erbB receptors were performed, using MLE 12 cells. Exposure of the cells to dexamethasone for 24 h followed by immunoprecipitation with a receptor-specific antibody caused an increase in erbB1 protein content to 159±32% (n = 9), in erbB2 to 186±27% (n = 17; p = 0.006), and in erbB4 to 193±23% (n = 11; p = 0.002), compared with untreated control cells (fig. 3a⇓). ErbB3 protein content was decreased to 88±13% (n = 12; fig. 3a⇓). Dexamethasone exposure changed the NRG-induced activity of the erbB receptors, measured by receptor phosphorylation. NRG increased the phosphorylation of erbB2 and erbB4 to 272±47% (n = 8; p = 0.008) and 161±14% (n = 6; p = 0.009) respectively, over baseline (fig. 3b⇓). However, the phosphorylation of erbB3, which is highly constitutively phosphorylated in these cells 6, was reduced by NRG to 71±8% (n = 5; p = 0.024) of baseline (fig. 3b⇓).

The effect of dexamethasone on a) erbB receptor protein levels and b) erbB receptor activation in mouse lung epithelial 12 cells. a) ErbB receptor protein levels in dexamethasone-treated cells were compared to protein levels in experiment-specific nontreated control cells. n = 9–17. b) ErbB2, erbB3, and erbB4 receptor phosphorylation in dexamethasone-treated cells stimulated with neuregulin (NRG) was compared with phosphorylation in experiment-specific dexamethasone-treated control cells not stimulated with NRG. n = 5–8. The figure shows quantification of Western blots by densitometry, standardised to actin. Data are presented as mean±sem. □: erbB1; ░: erbB2; ▓: erbB3; ▒: erbB4. *: p<0.05.
ErbB receptor dimerisation patterns in MLE 12 cells
Since a specific treatment can cause diverse biological effects through formation of distinct erbB receptor dimers, the dimerisation pattern of erbB receptors induced by dexamethasone were studied. Immunoprecipitations were carried out for each individual erbB receptor under low-stringency conditions. Again, these studies were performed in the MLE 12 cell line. The present authors have shown in these cells that erbB1 and erbB4 are preferred dimerisation pairs (table 1⇓) 16. In the current study, dexamethasone treatment led to a change in the dimerisation patterns of erbB2 and erbB4 (fig. 4⇓) compared with the untreated control erbB co-IP profile (table 1⇓). Dexamethasone treatment induced co-precipitation of erbB2 with erbB4 protein, suggesting a preferred erbB2–erbB4 heterodimerisation (fig. 4b⇓). This was not found in the untreated MLE 12 cells, in which erbB2 preferentially dimerised with erbB3. Also, the heterodimerisation pattern of erbB4 was disturbed by glucocorticoid treatment (fig. 4d⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
The effect of dexamethasone treatment on erbB receptor heterodimerization in mouse lung epithelial 12 cells. Low-stringency co-immunoprecipitation protocols were applied to study erbB dimer formation after 24-h dexamethasone treatment. Lysates from dexamethasone-treated cells were stimulated with growth factor (epidermal growth factor (EGF) and/or neuregulin (NRG)) or Dulbecco’s Modified Eagle Medium (controls). Precipitates with a) anti-erbB1 (αerbB1), b) anti-erbB2 (αerbB2), c) anti-erbB3 (αerbB3) and d) anti-erbB4 (αerbB4) were first probed with an antibody reacting with phosphorylated tyrosine residues and then re-probed with antibodies to each of the four erbB receptors.
Summary of reported major dimerisation partners in mouse lung epithelial 12 cells under co-immunoprecipitation conditions 16
The preferred heterodimerisation of erbB2 with erbB4 following dexamethasone treatment was unaffected by a 2-min stimulation with EGF. However, a 2-min stimulation with NRG induced an increase in both erbB2 and erbB4 phosphorylation (fig. 4b⇑), indicating further promotion of erbB2–erbB4 heterodimerisation. Otherwise, the co-IP characteristics of erbB1 (fig. 4a⇑) and erbB3 (fig. 4c⇑) were not affected by dexamethasone. In sum, dexamethasone treatment changed the preferential dimerisation patterns from erbB1–erbB4 dimers to erbB2–erbB4 in MLE 12 cells.
DISCUSSION
Optimal strategies to decrease the incidence of BPD in pre-term infants would involve the induction of surfactant synthesis without inhibition of alveolar development. Having previously shown that erbB receptor activation, mediated by mature or dexamethasone-induced immature FCM, stimulates surfactant synthesis 6, the present authors wished to study whether direct dexamethasone treatment would alter erbB receptor activation distinct from mature FCM in foetal type II cells. The effect of exogenous dexamethasone was therefore compared with the endogenous stimulation of foetal surfactant synthesis by foetal lung fibroblasts. The present results show that conditioned medium from mature (d18) fibroblasts, which contains NRG, induces surfactant DSPC synthesis in immature foetal mouse (d16) type II cells. This expands on the earlier finding that NRG stimulates surfactant synthesis in mature (d18) foetal type II cells 6, implying that even at that early gestational age NRG is able to induce foetal surfactant synthesis in isolated type II cells. In these immature cells, dexamethasone had no stimulatory effect on foetal surfactant synthesis. However, at a later point in gestation (d17) dexamethasone exhibited the same stimulatory effect as did the NRG-containing FCM. This is also in agreement with the present authors’ previous published work in mature d18 type II epithelial cells 6 and demonstrates that in immature type II epithelial cells (d16) NRG is superior in its stimulatory effect on foetal surfactant DSPC synthesis.
In order to elucidate the differences in erbB receptor activation induced by dexamethasone and mature FCM, the present authors studied erbB receptor content, phosphorylation and heterodimer formation, all elements that determine receptor responses to ligands such as NRG. Dexamethasone increased the protein content of all four erbB receptors in d17 foetal type II cells, similar to NRG-containing FCM. Dexamethasone did not affect baseline erbB receptor phosphorylation, but produced an upregulation of NRG-induced erbB2 phosphorylation. The present authors have previously shown that pre-treatment of immature (d17) foetal lung fibroblasts with dexamethasone produces an FCM which induces erbB2 phosphorylation in type II epithelial cells. This effect was similar to the effect of mature NRG-containing d18 FCM, but was not seen after exposure to FCM from d17 foetal fibroblasts 6. ErbB2 lacks the ability to bind a ligand directly; it is activated indirectly in a ligand-dependent manner via the formation of heterodimers with the other three erbB receptors 17. When activated, erbB2 is a potent receptor strongly involved in cancer biology 17 and frequently involved in cross-talk with other signalling pathways, including those of cytokines 18. The present findings suggest that dexamethasone signalling affects erbB2 signalling in a similar manner to findings in ovarian carcinoma epithelial cells 19.
The complexity and diversity of the erbB receptor signalling network and the resulting variety of biological effects following erbB receptor activation are mediated through receptor activation by specific ligands and utilisation of specific receptor dimers. This allows the initiation of specific signalling cascades and is likely to account for the markedly different biological results erbB receptors can induce 17. The present results indicate that dexamethasone exposure disturbs the preferred dimerisation of erbB1/erbB4 previously described in type II epithelial cells 16, promoting the dimerisation of erbB2 with erbB4. It is known that different circumstances preferentially induce the formation and activation of different erbB receptor dimer pairs 20, 21 leading to distinct profiles of gene expression 15. The current data suggest that, although it exerts stimulatory effects on the surfactant system, dexamethasone might have effects on other processes in lung development, e.g. on alveolarisation, through upregulation of erbB2.
One of the major signalling pathways of activated erbB1 involves the activation of PLCγ, which is known to be associated with cell differentiation 22. PLCγ is upregulated in the foetal lung shortly before the onset of foetal surfactant synthesis 23. In contrast to FCM containing NRG, dexamethasone did not promote erbB receptor-induced PLCγ expression. This is in agreement with the changes in dimerisation pattern, where erbB1 appears to play a less prominent role after dexamethasone treatment. Because upregulation of PLCγ appears to be involved in the physiological induction of foetal surfactant synthesis, it might be speculated that differences in signalling between mature FCM and dexamethasone might explain the differences in the biological effects seen in these cells. The deleterious effects of dexamethasone on alveolarisation might involve changes in the erbB receptor signalling network involving erbB2 upregulation and erbB1 downregulation.
The current results are potentially of clinical importance, because glucocorticoids, as known stimulators of foetal lung maturation 24, are recommended in the clinical setting of pre-term labour to stimulate foetal lung maturation 25. Pre-natal glucocorticoids have a stimulatory effect on: structural development 26; the oxidative system by inducing antioxidative enzymes 27; vascular development by decreasing vascular permeability 28; and development of the foetal surfactant system 29–31. There is also a growing literature showing inhibitory effects of glucocorticoids on other aspects of lung development such as alveolarisation 4, 32. More detailed information on the signalling pathways affected by glucocorticoids is needed to define means to decrease the unwanted side-effects of glucocorticoids.
In conclusion, dexamethasone not only stimulates the maturation of the surfactant system by inducing the production of neuregulin 6 by foetal lung fibroblasts, but also changes the expression and dimerisation patterns of erbB receptors in foetal type II cells to enhance specific yet diverse signalling pathways in the developing foetal lung. Differences in erbB receptor activation between mature fibroblast-conditioned medium and dexamethasone might provide an explanation for some of the adverse effects of dexamethasone (e.g. arrest of alveolarisation) in the developing foetal lung. Further research is necessary to elucidate the exact signalling mechanisms through the erbB signalling network in this context. A better insight into the interaction of signalling pathways of hormones and growth factors in foetal lung cell maturation would enable more specific induction of maturational processes and a reduction in effects that disturb other components of lung maturation following pre-natal exposure to glucocorticoids 33, 34.
Acknowledgments
The authors wish to thank L. Pham for preparing the primary mouse fibroblast and type II cell cultures.
- Received November 11, 2005.
- Accepted July 14, 2006.
- © ERS Journals Ltd
References










