





Abstract
The increased number of lymphocytes in airways during an asthmatic response is believed to be the result of increased recruitment of these cells. However, it is possible that a decreased apoptotic rate could also contribute to the increased number. The aim of the present study was to investigate whether allergen airway provocation influences the apoptotic phenotype of lung and peripheral blood lymphocytes (PBL) in subjects with atopic asthma.
Bronchoalveolar lavage (BAL) lymphocytes and PBL from 12 asthmatic subjects previously challenged with allergen (n = 7) or saline (n = 5) were exposed to the apoptotic stimulus tributyltin (TBT) in vitro and assayed for apoptosis.
Airway allergen provocation resulted in decreased sensitivity of BAL lymphocytes to TBT-induced apoptosis, with 42.2% (range 33.9–62.5%) apoptotic cells before challenge versus 23.5% (range 15.3–42.4%) after challenge, while PBL were unaffected. The increased apoptosis resistance correlated with higher numbers of Bcl-2-expressing lymphocytes. Interestingly, baseline caspase-3-like activity was significantly elevated in viable BAL lymphocytes compared with viable PBL, and was unaltered by allergen exposure.
In conclusion, allergen inhalation renders bronchoalveolar lavage lymphocytes more resistant to apoptosis while peripheral blood lymphocytes were not influenced at all, indicating that the apoptotic phenotype of airway lymphocytes may play a role in asthmatic inflammation.
Allergic asthma is a chronic disease in which allergen-induced inflammatory processes in the airways contribute to the development of symptoms and may eventually lead to remodelling of the airway tissue. Inflammation at the sites of target organs is a pathological feature of the disease process and among the various types of cells involved in tissue infiltration and damage are T-lymphocytes, i.e. one of the main effector cells priming the local ongoing allergic immune response 1. These cells are involved in the local recognition of allergens, acting through secreting T-helper (Th)2 cytokines and thereby determining immunoglobulin (Ig)E synthesis and the mucosal recruitment of other inflammatory cells, such as neutrophils and eosinophils 2, 3. Although knowledge of the initiation of the allergic response has rapidly expanded during recent years, little is known about how the inflammatory response develops into a chronic inflammation.
Previous studies have demonstrated that reduced apoptosis of T-cells may play a role in asthma pathogenesis 4, 5. Apoptosis of lymphocytes was decreased at baseline levels in asthmatic subjects compared with normal controls and patients with obstructive pulmonary disease 6. Additionally, T-cells from asthmatic subjects fail to undergo the normal degree of apoptosis following Fas receptor ligation, and CD45RO+ T-cells in the inner airway wall of patients with mild and serve asthma were less apoptotic compared with controls 7, 8. The observed insufficient T-cell apoptosis may interfere with clonal deletion and maintenance of tolerance, resulting in T-cell accumulation and contributing to the chronic inflammation of asthma. Thus, previous data indicate alterations in the apoptotic phenotype of cells involved in asthmatic inflammatory processes 4, 6, 7. However, changes in apoptosis sensitivity following an allergen challenge are still elusive.
Apoptotic cell death results from the activation of an internally encoded suicide programme, which is induced by a variety of intrinsic and extrinsic signals and is an important tool in the regulation of the immune system. The amount of reactive T-cells in an immune response can be controlled by a process called “activation-induced cell death”, which mainly involves the activation of death receptors such as Fas 9. However, apoptosis of activated T-cells can also be induced independently of death receptor signalling by the induction of the mitochondrial pathway, which is activated by signals such as cytokine withdrawal or the presence of reactive oxygen species 10–12. Death receptor-mediated apoptosis involves the recruitment of the initiator caspase-8, while the initiation of the mitochondrial pathway leads to the cytoplasmic release of pro-apoptotic factors from the mitochondrion and the subsequent activation of caspase-9 13, 14. The caspases belong to a family of cysteine proteases, which play a central role in the transduction of apoptotic signals, either by cleaving and activating downstream caspases or by processing vital proteins that will lead to the death of the cell. Until recently, caspase-3 has been considered to be an exclusively pro-apoptotic protein whose activation has been believed to be the “point of no return” in the apoptotic pathway. However, new evidence has emerged, suggesting an additional role for caspase-3 together with caspase-8 in differentiation and proliferation of activated T-cells 15–21.
High caspase-3-like activity in nonapoptotic bronchoalveolar lavage (BAL) lymphocytes recovered from sarcoidosis patients has previously been demonstrated 22. These lymphocytes were also shown to be highly resistant to mitochondrial-mediated apoptosis induced by tributyltin (TBT) 22. In the present study, allergen inhalation challenge of individuals with asthma was used to delineate the processes of apoptosis occurring in the inflammatory phase of asthma. As little is known about mitochondrial-induced apoptosis of airway lymphocytes in asthma, the current authors were particularly interested to investigate the impact of this pathway in these cells. Therefore the authors used TBT, a mitochondrial toxin that specifically triggers the mitochondrial pathway by loss of mitochondrial membrane potential and cytochrome C release 23–25.
The BAL lymphocytes from asthmatic subjects provoked with allergen were more resistant to mitochondrial-mediated apoptosis, while no change in apoptosis sensitivity was evident in BAL lymphocytes obtained from subjects challenged with saline. The decreased apoptosis sensitivity in allergen-exposed BAL lymphocytes was associated with a higher proportion of Bcl-2-expressing lymphocytes. Furthermore, BAL lymphocytes from all subjects had an elevated caspase-3-like activity compared with peripheral blood lymphocytes (PBL), and airway exposure to allergen did not increase the activity further. These data indicate that airway inflammation in asthma is associated with a reduced apoptosis susceptibility of lung lymphocytes, which may lead to an enhanced survival of BAL lymphocytes and consequently prolonged inflammation.
MATERIALS AND METHODS
Subjects
In total, 12 atopic patients (three male and nine female) aged 22–46 yrs (median 27 yrs) with a history of mild allergic asthma participated in the study. All were nonsmokers except one who was an ex-smoker. All subjects in the study population had previously documented reactions following allergen bronchoprovocation. The patients allergic to animals did not have pets of their own and were asked to avoid animal contact during the study period. Their asthma was stable and controlled solely with β2-agonists taken as required. As an inclusion criterion in the study the forced expiratory volume in one second (FEV1) was required to be >70% of predicted. Prior to the study, bronchial hyperresponsiveness to methacholine with a provocative dose causing a 20% fall in FEV1 (PD20) of <2,530 μg was demonstrated. The methacholine provocation test was performed as previously described 26. Patient characteristics and clinical data before and after allergen provocation are presented in table 1⇓. The study was approved by the Ethical Committee at the Karolinska University Hospital (Stockholm, Sweden), and all subjects gave their written informed consent.
Patient characteristics, clinical parameters and bronchoprovocation data
Study design
The study was performed out of the pollen season. Patients were blinded and administered either saline or an allergen (table 1⇑) for which they had a positive history and radioallergosorbent result. The allergen extracts used were standardised and freeze-dried (Aquagen; ALK, Copenhagen, Denmark). PBL and BAL samples were obtained 2–3 weeks before (baseline) and 1 day after the allergen/saline challenge. The type of allergen used for challenge, cumulative allergen dose, peak drop during the early asthmatic reaction (EAR) and the PD20 values are presented in table 1⇑. All bronchial provocations were performed using a dosimeter-controlled jet nebuliser (Spira Electro 2; Respiratory Care Center, Hameenlinna, Finland). In brief, the allergen challenge was initiated by inhalation of dilutent. Provided that the FEV1 did not change >10%, allergen was inhaled every 15 min with half-log increments of the cumulated dose until the FEV1 dropped ≥20% from the post-dilutent baseline value 27. The occurrence of any late asthmatic reactions (LAR) were examined by measurements of peak expiratory flow rates every waking hour during the first 24 h following challenge. The patients were instructed to make additional recordings if any airway symptoms occurred.
BAL, differential cell count, separation and handling of cells
BAL was performed as previously described 28, 29. In brief, under local anaesthesia the flexible fibreoptic bronchoscope (Olympus BF type 1TR; Olympus Optical Co Ltd, Tokyo, Japan) was wedged into a middle lobe bronchus and sterile PBS solution at 37°C was instilled in five aliquots of 50 mL. After each instillation, the fluid was gently aspirated and collected in a siliconised plastic bottle kept on ice. The BAL was strained through a Dracon net (Millipore, Cork, Ireland), centrifuged at 400×g for 10 min at 4°C, and the cell pellet was resuspended in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma chemicals, St Louis, MO, USA). Cells were counted using a Bürker chamber and cell viability was determined by trypan blue exclusion. Peripheral blood mononuclear cells (PBMC) were obtained by Ficoll-Hypaque gradient centrifugation (Amersham Pharmacia Biotech, Uppsala, Sweden), washed three times in PBS and diluted in RPMI 1640. For differential cell counts, cytospins were prepared by cytocentrifugation (Cytospin 2; Shandon, Runcorn, UK) of 60,000 cells per slide at 20×g for 3 min. In order to determine cell differentials, cytospin slides were stained with May–Grünwald–Giemsa and 400 cells were counted microscopically. Mast cells were stained using two parts toluidine blue 0.5% (pH 0.5) and one part 0.2% haematoxylin solution. The number of mast cells from 10 vision fields, at a magnification of ×16, was counted microscopically. Both BAL cells and PBMC were kept on ice until the initiation of the experiments.
Induction and detection of cell death
BAL cells and PBMC were centrifuged at 400×g for 10 min at 4°C and resuspended in complete medium at a concentration of 1×106 cells·mL-1, complete medium (RPMI 1640 containing 5% foetal calf serum), 2 mM L-Glutamine, penicillin (100 U·mL-1) and streptomycin (0.1 mg·mL-1; all Sigma Chemicals). Apoptosis was induced by exposure of the cells to 2 µM TBT chloride (Sigma Aldrich, Munich, Germany) for 60 min at 37°C in a humidified air chamber with 5% CO2. Dead cells were detected using fluorescein isothiocyanate (FITC)-labelled annexin V and propidium iodide (PI; annexin V/PI; Pharmingen, San Diego, CA, USA) staining. Briefly, BAL cells and PBMC were resuspended in annexin-binding buffer containing annexin V-FITC and PI (100 µg·mL-1) and incubated for 15 min at room temperature. The samples were analysed using a flow cytometer (FACS Calibur; Becton Dickinson, Franklin Lakes, NJ, USA). BAL lymphocytes and PBL were identified and gated by forward and side light-scattering properties. Ten thousand events were analysed and dot plots were used to determine the percentage of apoptotic cells showing annexin V-FITC (fluorescence (FL)-1 channel) binding; a minority of these cells were also permeable to PI (FL-3 channel). “Apoptotic cells” in the Results section refers to lymphocytes stained with annexin V, with or without PI staining, mainly including cells in early apoptosis (annexin V positive/PI negative; encompassing the vast majority of the cells) but also cells in late apoptosis (annexin V positive/PI positive; minority).
Detection of caspase-3-like activity in intact cells
Caspase-3-like activity in intact BAL and PBL was measured using the cell-permeable substrate PhiPhilux-G2D2 (OncoImmunin, Inc., Kensington, MD, USA), according to the manufacturer’s recommendations. Briefly, cells were washed, resuspended in 50 µL substrate (10µM) and incubated for 1 h at 37°C in a humidified air chamber with 5% CO2 followed by immediate flow cytometric analysis (FL-2 channel). Caspase-3-like activity was expressed as mean fluorescence intensity from viable lymphocytes which were identified and gated by forward and side light-scattering properties.
Immunocytochemistry double staining
Cytospins were allowed to reach room temperature and the cells were fixed in paraformaledyde 4% for 15 min. Antigen unmasking was performed by boiling the cytospins in 10 mM citrate buffer for 15 min and then allowing cooling for 45 min at room temperature. Cells were permeabilised using ice-cold methanol for 10 min at -20°C. To avoid nonspecific staining, the cells smears were incubated in blocking buffer containing PBS, 5% horse serum and 0.1% bovine serum albumin for 1 h at room temperature or overnight at 4oC. The cell smears were incubated with goat anti-CD33 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted 1:100 in blocking buffer overnight at 4oC. Cell smears were washed three times for 5 min in Tris-buffered saline (TBS); in the second wash 0.1% TWEEN 20 was added. Alkaline phosphatase-conjugated rabbit anti-goat (Sigma Aldrich) was diluted 1:100 in TBS containing 5% horse serum, and 0.1% bovine serum albumin was then added for 1 h at room temperature. The cell smears were washed three times for 5 min in TBS, with 0.1% TWEEN 20 added in the second wash before staining using the alkaline substrate Fast Red for 10 min. The slides were washed in running water for 10 min, incubated in PBS for another 10 min, followed by a second primary antibody. The antibodies used were either mouse anti-human Bcl-2 1:50 (clone 124; Dako Cytomation, Glostrup, Denmark) and proliferating cell nuclear antigen (PCNA) 1:500 (clone PC10; Calbiochem, Darmstadt, Germany) or mouse IgG as isotype control (Dako Cytomation), which were diluted in blocking buffer. The cell smears were incubated with the second primary antibody overnight at 4°C. Endogenous peroxide activity was quenched by incubating the slides with 0.3% H2O2 in PBS for 30 min. Following incubation with secondary antibody, biotinylated horse anti-goat IgG (Vector Laboratories, Inc., Burlingame, CA, USA) diluted in blocking buffer 1:100 for 1 h, the cell smears were incubated with avidin-conjugated peroxidase substrate Vectasin elite ABC kit (Vector Laboratories, Inc.) for 90 min. 3,3′-Diaminobenzidine (Dako Cytomation) was used for colour development. The exposure time was 8 min for Bcl-2 staining, 5 min for the PCNA antibody and 10 min for the IgG isotype control antibody. The stained cytospins were counted in a blinded manner three times.
Statistics
Results are presented as median values with minimum and maximum values as range. Comparisons of data retrieved before and after provocation were made using Wilcoxon matched-pairs analysis. A p-value <0.05 was considered significant.
RESULTS
Clinical parameters
All patients provoked with allergen, but not those receiving saline, had an EAR defined as a ≥20% decrease in FEV1 within the first 3 h post-challenge. One of these patients also had a LAR, defined as >15% decrease in FEV1 during 3–24 h post-challenge (table 1⇑).
BAL and blood cell data
BAL recovery and cell viability did not differ in samples retrieved at baseline and after allergen or saline challenge. Although an increased BAL cell concentration was recorded in 10 out of the 12 patients, i.e. in both allergen- and saline-exposed individuals compared with baseline, there was only a statistically significant (p<0.05) increase determined in those patients challenged with saline (table 2⇓). The differential count of BAL cells revealed a significant increase in the percentage of eosinophils following allergen challenge (p<0.05). However, both allergen and saline seemed to influence the amount of eosinophils in the lungs, since three out of five patients challenged with saline and five out of seven patients challenged with allergen had an increase in the proportion of BAL eosinophils compared with baseline levels. A tendency of a relative increase in BAL lymphocytes was observed in patients challenged with allergen. Here, the differential count showed an increase of lymphocytes in six out of seven patients challenged with allergen compared with only one out of five challenged with saline (table 2⇓). The proportion of T-cells was 86% (75–91%) among the BAL lymphocytes and 74% (61–86%) in PBL. The differential cell counts of whole blood revealed no change in the cellular composition at baseline and after allergen or saline challenge (data not included).
Bronchoalveolar lavage cell count data before and after diluent or allergen exposure
Determination of apoptosis in BAL cells and PBL at baseline and after allergen/saline challenge
Paired BAL lymphocytes and PBL from all individuals were tested for their apoptosis sensitivity at baseline and post-challenge with either allergen or saline. Susceptibility to apoptosis induced by the mithochondrial pathway was investigated by stimulation with 2 μM TBT in vitro, an agent that specifically induces mitochondrial permeability transition, loss of mitochondrial membrane potential, cytochrome C release and apoptosis 24, 25. Apoptotic cells were identified using annexin V staining and fluorescence-activated cell sorter analysis. The rate of spontaneous apoptosis among BAL lymphocytes and PBL was not altered by airway allergen or saline exposure (tables 3⇓ and 4⇓).
Apoptosis susceptibility of bronchoalveolar lavage lymphocytes in asthmatic subjects at baseline and after allergen or saline inhalation
Apoptosis susceptibility of peripheral blood lymphocytes from asthmatic subjects at baseline and after allergen or saline inhalation
TBT exposure of BAL lymphocytes from patients in the allergen provocation group (n = 7) resulted in 42.2% (33.9–62.5%) apoptotic BAL lymphocytes at baseline. After airway allergen challenge, the same treatment of the BAL cells resulted in a significant decrease in apoptotic lymphocytes (p = 0.023), since only 23.5% (15.3–42.4%) of the lymphocytes became apoptotic (table 3⇑; fig. 1a⇓). This phenomenon was not recorded in the group of patients who inhaled saline (n = 5). In this group, TBT induced apoptosis in 37.8% (26.6–41.5%) of the BAL lymphocytes obtained at baseline and in 36.8% (14.5–48.5%; nonsignificant) of the lymphocytes retrieved 24 h post-saline challenge (table 3⇑; fig. 1b⇓).

Apoptosis susceptibility of bronchoalveolar lavage (BAL) lymphocytes at baseline and after a) allergen or b) saline challenge. BAL cells were exposed to 2 µM tributyltin (TBT) for 60 min followed by annexin-V staining and fluorescence-activated cell sorter analysis. Data depict the change in per cent apoptotic cells after TBT treatment compared to control cells, at baseline and after allergen or saline inhalation. ns: nonsignificant. #: p<0.023.
Inhalation of allergen or saline did not significantly alter the sensitivity of PBL to TBT-induced apoptosis. In the allergen provoked group, 35.7% (21.4–69.9%) of the control lymphocytes were apoptotic upon treatment with 2 μM TBT while the same treatment 24 h after challenge induced apoptosis in 47.0% (11.8–80.06%; nonsignificant) of the lymphocytes (table 4⇑; fig. 2a⇓). In the group of patients inhaling saline, TBT exposure of PBL induced apoptosis in 51.4% (28.6–75.1%) of the lymphocytes at baseline while the same treatment 24 h after challenge triggered apoptosis in 40.5% (24.6–75.8%) of the PBL (nonsignificant; table 4⇑; fig. 2b⇓).

Apoptosis susceptibility of peripheral blood lymphocytes (PBL) at baseline and after a) allergen or b) saline challenge. Influence of tributyltin (TBT) on the induction of apoptosis in PBL of asthmatic subjects provoked by allergen (a) or saline (b). Data represent the change in per cent apoptotic cells after TBT treatment, before and after allergen challenge. ns: nonsignificant.
Analysis of Bcl-2 and PCNA in BAL lymphocytes
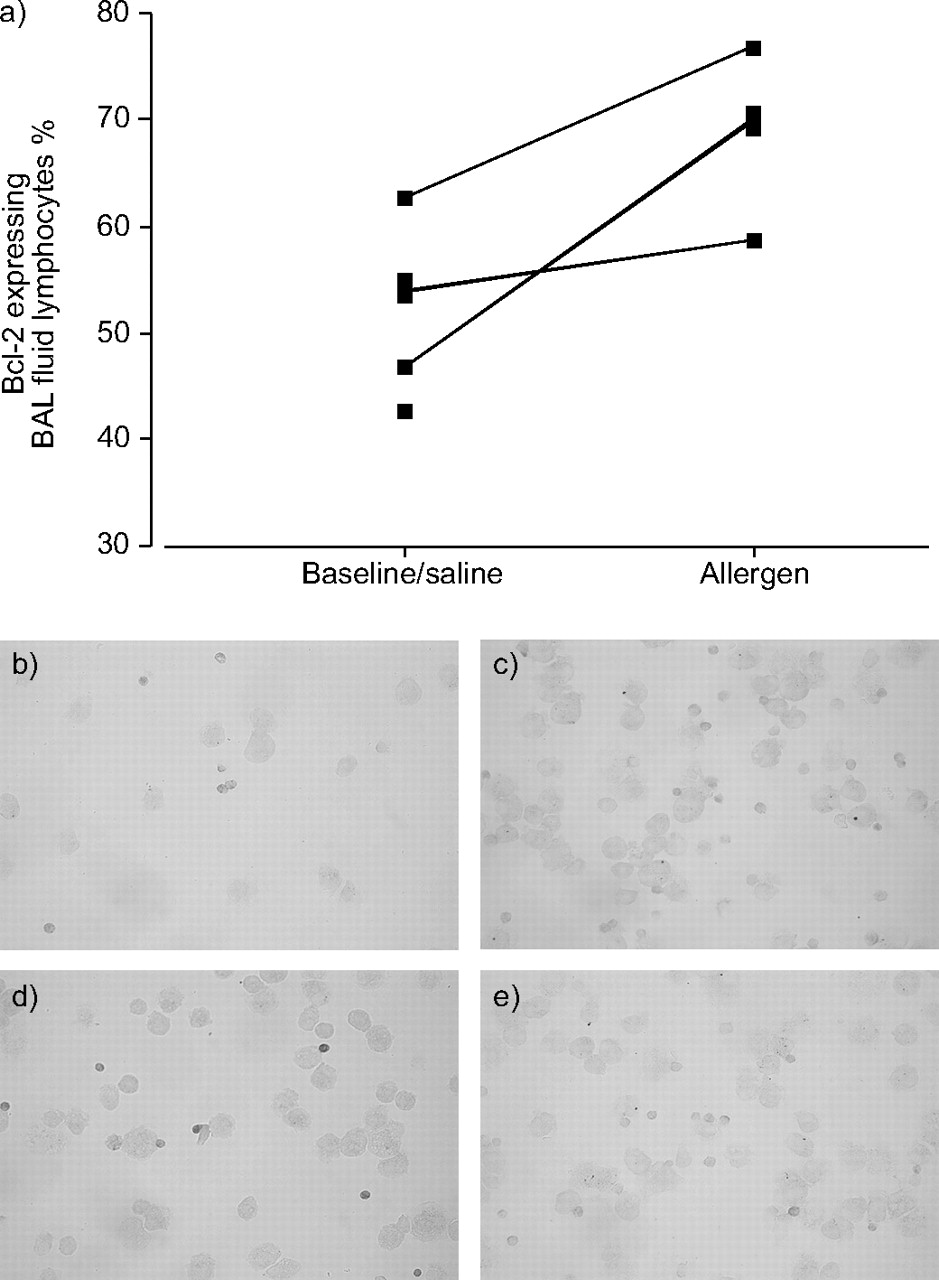
BAL lymphocytes (cytospins) from three subjects at baseline and post-allergen challenge, respectively, were stained for the expression of the anti-apoptotic protein Bcl-2 and for PCNA (proliferation marker) together with an antibody against CD3. Allergen exposure increased the proportion of Bcl-2-expressing BAL lymphocytes, the majority being T-cells, from 54% (47–63%) to 70% (59–77%) in the paired samples, i.e. at baseline and after allergen challenge from the same subject (fig. 3a⇓). Data from unpaired samples from three additional patients at baseline (patient No. 9), following allergen exposure (patient No. 10) and after saline challenge (patient No. 12) were in line with the previous results (55, 71 and 49% Bcl-2-expressing BAL lymphocytes in patient Nos 9, 10 and 12, respectively). One representative experiment of BAL cells from patient No. 11 stained for Bcl-2 at baseline and 24 h after challenge is also presented (fig. 3b–e⇓).

a) Bronchoalveolar lavage (BAL) fluid cells on cytospins from six subjects were stained using immunocytochemistry (ICC), for the expression of BcI-2. Paired samples (baseline and 24 h after airway allergen challenge from the same subject) are indicated with lines. Representative ICC experiments of BcI-2 expression, darker stain (3,3’-diaminobenziding) in BAL fluid lymphocytes from one subject, at baseline (b) together with isotype-matched control (c) and Bcl-2 expression after airway allergen challenge (d) and isotype-matched control stain in these cells (e).
All BAL lymphocytes analysed were PCNA negative at baseline and after saline and allergen challenge, indicating no proliferative activity among these cells (data not included).
Detection of caspase-3 activity in BAL lymphocytes and PBL at baseline and after airway allergen challenge
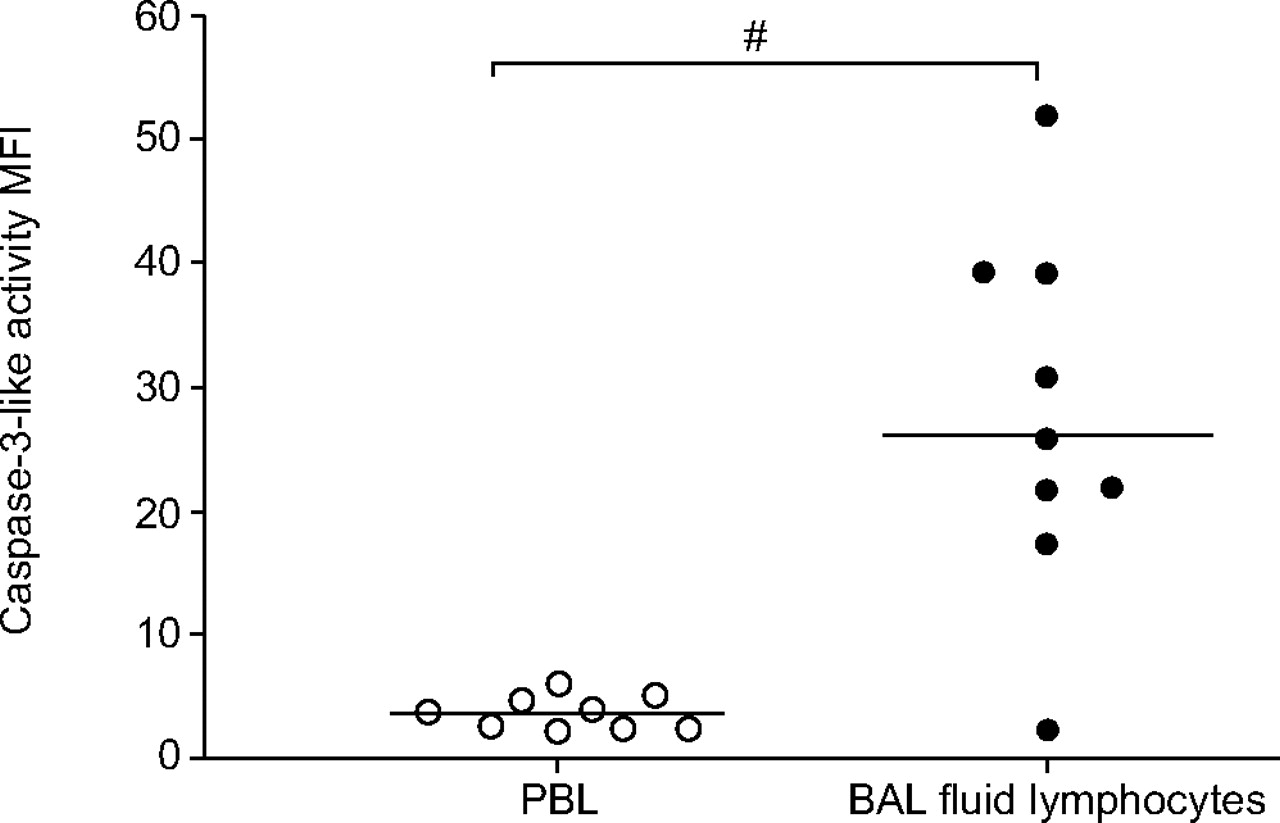
To evaluate whether the pro-apoptotic enzyme caspase-3 was activated in BAL lymphocytes and PBL, cells were loaded with a fluorigenic caspase-3 substrate and the production of the cleavage product was monitored using flow cytometry. BAL lymphocytes possessed significantly higher endogenous caspase-3 activity than PBL (p = 0.002; fig. 4⇓). However, allergen provocation did not significantly alter the basal caspase-3 levels in either BAL lymphocytes or PBL (fig. 5⇓).

Baseline caspase-3-like activity in peripheral blood lymphocytes (PBL) and in bronchoalveolar lavage (BAL) fluid lymphocytes from asthmatic subjects. Cells were loaded with the cell-permeable substrate PhiPhilux (10 μM) to detect caspase-3-like activity. Data are expressed as mean fluorescence intensity (MFI) and the lines indicate median values. #: p = 0.002.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Caspase-3-like activity in a) bronchoalveolar lavage lymphocytes and b) peripheral blood lymphocytes at baseline and after allergen challenge. MFI: mean fluorescence intensity; ns: nonsignificant.
DISCUSSION
The present study reported that BAL lymphocytes from asthmatic subjects-exposed to allergen were significantly more resistant to mitochondrial-mediated apoptosis compared with nonallergen-exposed BAL lymphocytes from the same subjects. This decreased sensitivity to apoptosis was accompanied by an increase in the proportion of BAL lymphocytes expressing the Bcl-2 protein. However, airway challenge with saline did not alter the sensitivity of BAL lymphocytes to TBT-induced apoptosis and neither did allergen or saline provocation influence the sensitivity of PBL to mitochondrial-mediated apoptosis. These data suggest that airway provocation with allergens triggers an inflammatory response, which results in the alteration of the apoptotic phenotype of lung lymphocytes but not of PBL. The observed discrepancy in apoptosis sensitivity between BAL and PBL following allergen challenge may reflect the different activation status of the cells in these two compartments; it is known that airway allergen challenge can cause an accumulation of activated allergen-specific lymphocytes in the airways 2, 30, 31. The present results are concordant with previous studies of apoptosis in lymphocytes activated in vitro 32 and in BAL cells from patients with sarcoidosis 22.
The major proportion of lung lymphocytes are T-cells 33, which in asthma are characterised by a Th2-phenotype 1. T-cells play a role in controlling chronic inflammation in asthma 34 and the number of T-cells infiltrating the bronchial airways is significantly increased in this disease 2, 31.
It is accepted that the increased numbers of mucosal T-cells is the result of an increased recruitment of these cells within the airways 3, 35, 36. However, it is also likely that the number of T-cells in the airways of asthmatic patients depends upon an increased survival of these cells. This hypothesis supports several studies in which it has been reported that cells engaged in the inflammatory asthmatic process are less apoptotic.
The potential role of the Fas receptor pathway in asthma has been intensively studied during recent years, albeit with contradictory results. Some studies have demonstrated a decreased expression of Fas receptor or Fas ligand 6 in asthmatic subjects while other studies have reported an increased expression of Fas receptor and Fas ligand in asthmatic subjects 37, 38. Another study showed that T-cells from asthmatic subjects failed to undergo apotosis following Fas receptor ligation, although these cells express the same levels of Fas and Fas ligand as in nonasthmatic subjects 7.
The current authors were particularly interested to study the sensitivity of lymphocytes to mitochondrial-mediated apoptosis in asthma, since this pathway has been suggested to play a substantial role in downregulation of an immune response 10. Therefore the present authors used TBT, a mitochondrial toxin that specifically triggers the mitochondrial pathway by loss of mitochondrial membrane potential and cytochrome C release 23–25.
The Bcl-2 family consists of both proteins that protect from mithochondrial-mediated apoptosis, such as Bcl-2 and Bcl-xL, and proteins that promote this apoptotic pathway, i.e. Bax, Bak, Bid and Bad 39. The Bcl-2 protein is mainly localised in the mitochondria membrane where it blocks the release of cytochrome C and consequently caspase activation and apoptosis 39–41. In the present study, an increase in resistance to mitochondrial-mediated apoptosis in BAL lymphocytes was observed following airway allergen challenge and this was associated with an increase in the proportion of Bcl-2-expressing lymphocytes. These data agree with several previous reports. In bronchial biopsies taken from asthmatic subjects, most T-cells do not appear to be apoptotic and express Bcl-2, suggesting that these cells may have the ability to live longer in the inflamed airways 4. This was also confirmed by results obtained using an allergen-induced cutaneous late-phase response model 42. Moreover, a recent study reported that cyclosporin treatment of asthmatic subjects prior to airway allergen challenge resulted in an increased apoptosis and reduced Bcl-2 expression among BAL lymphocytes 43. In addition, Bcl-2 expression was increased in sputum mononuclear cells from asthmatic patients compared with patients with chronic obstructive pulmonary disease and healthy controls 6.
Sensitised individuals with allergic asthma develop an EAR 10–20 min after airway allergen challenge, which resolves after ∼1 h. The bronchial constriction, increased vascular permeability and mucus production during EAR is caused by rapidly metabolised mediators, such as histamine, prostaglandins and leukotrienes, which are released by IgE cross-linked mast cells. Some individuals develop a LAR 3–4 h after challenge, which can persist for ≥24 h. LAR is caused by the induced synthesis and release of inflammatory mediators including leukotrienes, chemokines and cytokines, which recruits other leukocytes, such as eosinophils and T-lymphocytes to the bronchial mucosa 44. In the present study, airway allergen challenge induced EAR in all exposed subjects, although only one out of these seven individuals developed LAR. However, allergen exposure resulted in a significant rise in the proportion of BAL eosinophils, although an increase in lung eosinophils was also seen in three out of five subjects receiving saline. Interestingly, in six out of seven subjects, inhalation of airway allergen resulted in a nonsignificant increase in BAL lymphocytes and this tendency was not found among the saline-exposed individuals, suggesting an enhanced recruitment and/or survival of these cells after an allergen exposure which does not induce LAR 45, 46. The increased apoptosis resistance of BAL lymphocytes observed in the current study may reflect a cellular event that allergen, present in the normal milieu of the asthmatic individual, induces in airway lymphocytes. Decreased apoptosis sensitivity among these lymphocytes could contribute to the development of a subclinical inflammation in the bronchial mucosa resulting in thickening of the basement membrane and remodelling of the airways.
Several lines of evidence suggest additional nonapoptotic functions for traditionally pro-apoptotic caspases. Active caspase-3 participates in the differentiation of such diverse cell types as neurons, muscle cells, monocytes and erythroblasts 15–18. Moreover, the proliferative machinery of T-cells appears to require active caspase-8 and caspase-3, and the activation of caspase-3 has been proposed to be essential for cell cycle progression in B-cells 19, 47–49. Although it is established that caspases possess a nonapoptotic role in activated T-cells, it is not clear what initiates this activity and what the caspase-3 substrates are. However, Wu et al. 50 suggest that the transcription factors nuclear factor (NF)-ATc2 and NFAT, which are crucial for cytokine production and activation of T-cells, are substrates of caspase-3. BAL lymphocytes seem to have higher caspase-3-like activity, an observation that could be related to the higher activation status of lymphocytes in the lungs compared to peripheral lymphocytes 22. This was further confirmed in the present study, in which viable BAL lymphocytes from asthmatic subjects had a significantly higher caspase-3-like activity than viable PBL from the same subjects. However, no PCNA expression was detected in BAL lymphocytes from the six subjects tested, suggesting that the elevated caspase-3 activity observed was not associated with proliferation in these cells.
It is still unclear how cells with high caspase-3 activity could be rescued from the proteolytic degradation machinery that is normally active in an apoptotic cell. There must be tight regulation of the apoptotic machinery in activated cells in order to prevent caspase-3 from completing its commitment to degrade vital proteins. How such protection is achieved is not yet known, but compartmentalisation, conformational changes of the target proteins or protection via chaperones are possible mechanisms.
In conclusion, the current authors have demonstrated an alteration in the apoptotic machinery of bronchoalveoar lavage lymphocytes recovered from asthmatic subjects following airway allergen. This altered apoptotic phenotype may not be due to changes in the receptor inducible pathway but are definitely related to the mitochondrial apoptotic pathway. The present authors also recorded a significantly higher baseline level of caspase-3 activity in bronchoalveolar lavage lymphocytes from these asthmatic subjects compared with peripheral blood lymphocytes from the same subjects. These data indicate that airway inflammation in asthma is associated with a reduced apoptosis susceptibility, which may lead to an enhanced survival of lymphocytes in the bronchial mucosa and consequently prolonged inflammation.
- Received October 10, 2005.
- Accepted May 20, 2006.
- © ERS Journals Ltd
References










