





Abstract
CC chemokine ligand (CCL)1/I-309 is a potent attractant for T-helper cell type 2 lymphocytes. The present study investigates whether this cytokine is released in the bronchoalveolar fluid (BALF) of asthmatic patients.
Measurements of CCL1 using ELISA showed that levels of this cytokine were significantly elevated in BALF from asthmatics compared with normals (median (range) 193 (120–449) pg·mL−1 versus 30 (21–55) pg·mL−1). Differential cell counts in BALF showed that either lymphocyte or eosinophil numbers were elevated in asthmatic compared with normal subjects (10.8×103·mL−1 versus 1.0×103·mL−1 and 1.7×103·mL−1 versus 0.2×103·mL−1, respectively). There was a trend towards a significant correlation between CCL1 levels and lymphocyte numbers in BALF. Separation of BALF using sequential CCL1 affinity column and reverse-phase high-performance liquid chromatography allowed detection of biologically active CCL1. Using immunohistochemistry, CCL1 immunoreactivity was localised predominantly to the airway epithelium.
Interestingly, there was a significant correlation between CC chemokine ligand 1 levels and epithelial cell numbers in bronchoalveolar lavage fluid and between these cells and lymphocyte numbers. Moreover, interleukin-4, interleukin-13 and interferon-γ stimulated primary bronchial airway epithelial cells to release CC chemokine ligand 1. These findings suggest that CC chemokine ligand 1 may play a role in lymphocyte recruitment in bronchial asthma.
It is now widely recognised that asthma is a chronic inflammatory airway disease characterised by infiltration of eosinophils and T-cells. CD4 T-cells that produce T-helper cell (Th) type 2 cytokines, including interleukin (IL)-4, IL-5 and IL-13, play an important role in orchestrating immune and inflammatory processes that lead to immunoglobulin (Ig)E production and allergic inflammation 1. Activated CD4 T-cells have been a consistent finding in the bronchoalveolar lavage fluid (BALF) and bronchial biopsies of stable asthma 2, 3. CD4 lymphocytes have also been shown to increase further following allergen challenge 4, 5.
Lymphocyte recruitment from peripheral blood into the airways is regulated by chemokines. The chemokines are a large family of 8–14 kD heparin-binding peptides, which have been subdivided into four subfamilies on the basis of the position of either one or two cysteine residues located near the amino terminus of the protein (CXCL, CC chemokine ligand (CCL), CL and CX3CL) 6, 7. The cytokine CCL1/I-309 is a member of the CCL chemokine subfamily, which was initially discovered as a gene of unknown function by subtractive hybridisation from the human IL-2-dependent γδ T-cell line IDP2 8, but was subsequently shown to be a monocyte and Th2 cell attractant 9–11. CCL1 is located on human chromosome 17 12. Upon activation, CCL1 is produced by a number of cellular sources, including peripheral blood mononuclear cells, monocytes, activated T-lymphocytes and endothelial cells 8, 13, 14. In vitro studies have shown that CCL1 as well as both CCL22/monocyte-derived chemokine (MDC) and CCL17/thymus- and activation-regulated chemokine (TARC) induce migration of lymphocytes, especially of the Th2 phenotype 13, 14. CCL22 and CCL17 activate these cells through the CC chemokine receptor (CCR)4, whereas CCL1 uses CCR8 15, 16. It is now well established that CCL22 and CCL17 are released into the airways of asthmatic patients 17–19. However, the role of CCL1 in asthma remains to be shown. Evidence for the participation of CCL1 in an in vivo role in allergic inflammation derives from an animal study showing that neutralisation of CCL1 with an anti-CCL1 antibody significantly inhibits eosinophil recruitment into the lungs of sensitised mice 20. Another approach has been the use of CCR8 knockout mice. For example, Chensue et al. 21 demonstrated that CCR8-/- mice had impaired pulmonary eosinophilia. However, Chung et al. 22 were unable to show any impairment in both pulmonary Th2 cytokine responses and eosinophilia in CCR8 deficient mice. Although mouse models of asthma have shed light onto the pathogenesis of asthma, they do not necessarily mirror human asthma. In the present study, the authors have investigated CCL1 in the BALF of stable asthmatics.
METHODS
Subjects
A group of 22 symptom-free asthmatics treated with inhaled salbutamol alone and 13 healthy subjects volunteered to participate in the study. The clinical characteristics of both asthmatic and normal subjects are shown in table 1⇓. Their atopic status was confirmed by skin-prick testing with different allergens (i.e. Dermatophagoides pteronnisinus and D. farinae, mixed grass, tree pollen, cat and dog dander and cockroach). The ethics committee of the National Institute of Respiratory Diseases (Mexico City, Mexico) approved the study.
Clinical characteristics of subjects
Fibreoptic bronchoscopy
Bronchoscopy was undertaken according to National Institutes of Health guidelines 23. Bronchoalveolar lavage (BAL) was performed with saline solution. The recovered BALF was pooled and centrifuged, cells were separated and supernatant was frozen. For differential cell counts, 100 μL of cells (4×105 cells) were subjected to cytocentrifugation, and were air dried and stained.
High-performance liquid chromatography
High-performance liquid chromatography (HPLC) was performed at room temperature with an HP1100 liquid chromatography system (Agilent Technologies, Palo Alto, CA, USA) equipped with a quaternary pump and a variable wavelength detector.
Purification of lymphocyte chemotactic activity in BALF
BALF from five subjects was combined and concentrated using Amicon YM3 filters (Millipore Corporation, Bedford, MA, USA) before application to a CCL1 affinity column, which had been previously prepared by loading an anti-CCL1 antibody (AF272; R&D systems, Minneapolis, MN, USA) on a N-Hydroxysuccinimide-activated sepharose column (HiTrap, 5 mL; Pharmacia, Uppsala, Sweden) 24. Bound material was eluted from the affinity column and separated by reverse-phase chromatography, as previously described 24, 25. Fractions obtained from the column were lyophilised before assay for lymphocyte chemotaxis.
Isolation of CD4+ lymphocytes
Human CD4+ T-cells were purified from healthy donor peripheral blood using Ficoll-Hypaque (Pharmacia Biotech AB, Uppsala, Sweden) density gradient centrifugation, followed by isolation with immunomagnetic beads (CD4+ T-cell isolation kit; Milteny Biotech, CA, USA). Purity was routinely >95%. For chemotaxis CD4+ cells were stimulated with IL-2 for 5 days. Consistent with a previous report 26 at day 5 of incubation with IL-2, CD4+ cells were found to express the CCL1 receptor CCR8.
Chemotaxis assays
Chemotaxis of BAL HPLC fractions was performed using a multi-well microchemotaxis chamber, as previously described 27. Recombinant CCL1 was used as a positive control. Chemotaxis was expressed as a chemotactic index. The anti-CCL1 antibody monoclonal antibody (MAB) 272 was used to neutralise experiments.
SDS-PAGE and Western blot
Sodium dodecyl sulphate-polyacrylamide electrophoresis (SDS-PAGE) was performed using Phast-Gel high-density gel (Pharmacia) according to the manufacturer's protocol. Briefly, electrophoresis was performed at pH 6.4 with Phastgel SDS buffer strips (Healthcare Biosciences, Uppsala, Sweden) that contained 0.112 M Tris/0.112 M acetate buffer, and was visualised by use of a silver staining kit (Sigma, St Louis, MO, USA). Western blot analysis was performed to detect CCL1 using a specific polyclonal antibody (anti-human CCL1 antibody; Peprotech Inc., Rockhill, NJ, USA).
Immunohistochemistry
Bronchial biopsies were fixed in paraformaldehyde and were paraffin embedded. Immunohistochemistry for CCL1 was performed by indirect immunoperoxidase staining, as previously described 28. The sections were counterstained using Mayer's haematoxylin (Vector Laboratories Inc., Burlingame, CA, USA).
RNA preparation and analysis
Total RNA was isolated from BAL cells using the TRIzol method. Total cellular RNA was quantified and reverse transcribed for CCL1, CCR8, reduced glyceraldehyde phosphate dehydrogenase and β2 microglobulin PCR reaction, as previously described 29. Specific primers had the following sequences: CCL1 (Forward 5′-GTTGCTTCTCATTTGCGG-3′; Reverse 5′-GTAGGGCTGGTAGTTTCGG-3)′; CCR8 (Forward 5′-CATCACCCTCATGAGTGTGG-3′; Reverse 5′- CACGTTGAATGGGACCCAGA-3′); β2 microglobulin (Forward 5′-GCTTACATGTCTCGATCCGACTTAA-3′; Reverse 5′-CTCGCGCTACTCTCTCTTTCTGG-3′).
Culture and stimulation of human airway epithelial cells
Human bronchial airway epithelial cells (BAEC) were isolated from bronchial epithelia mucosa tissue and grown as previously described 24. Subsequently, cells were incubated with IL-4, IL-13 and interferon (IFN)-γ at concentrations of 10 ng·mL−1 for 24 h. The purity and identity of the cells were checked by immunohistochemistry using the anti-citokeratin MAB AE1/AE3 (Zymed, San Francisco, CA, USA).
Measurement of CCL1
Measurement of immunoreactive CCL1 in both 2×concentrated BALF and airway epithelial culture supernatant was performed using a two-AB sandwich ELISA kit (R&D Systems) following the manufacturer's protocol. The primary anti-CCL1 capture antibody was the MAB 272; the secondary antibody was a biotinylated anti-CCL1 antibody CCL1 (R&D Systems). A list of the antibodies used in the different experiments is shown in table 2⇓. A standard curve consisting of serial dilutions from 0.007–1 ng·mL−1 of recombinant human CCL1 (R&D Systems) was included in the assay. Concentration of CCL1 in samples was calculated from the standard curve, and, in the case of BALF, concentrations of this cytokine were corrected for the initial two-fold concentration. The lower limit of detection was 7 pg·mL−1 and the inter-assay coefficient variation was 5%.
Antibodies used in experiments
Statistical analysis
Analysis of CCL1 in BALF and BAL cell counts was performed using the Mann–Whitney U-test for unpaired data. Correlations between CCL1 and infiltrating cells in BALF were evaluated using Spearman's nonparametric rank correlation coefficient test. A value of p<0.05 was considered statistically significant.
RESULTS
Clinical findings
The median age for asthmatics and controls was 30 and 33 yrs, respectively. Lung function data showed that asthmatics suffered from mild-to-moderate asthma. The forced expiratory volume in one second (FEV1; median) % predicted in asthmatics and normals was 89.5% (range 70–120%) and 104.4% (90–138%), respectively. All asthmatic patients were atopic; control subjects were nonatopic. Clinical data are shown in table 3⇓. No complications were seen in any of the patients following bronchoscopy. Minor complaints, such as sore throat and mild cough, were seen in three patients.
Differential and total cell counts in bronchoalveolar lavage fluid(×103·mL−1)
BALF and cell population
BAL was obtained from all 35 subjects taking part in the study. There was no significant difference in the volume of fluid recovered from asthmatic and normal subjects (75±10 mL versus 62±12 mL; p<0.05).
Differential cell counts in BAL showed that lymphocytes, eosinophils and epithelial cells numbers were significantly elevated in asthmatics compared with normals (10.8×103·mL−1 versus 1.0×103·mL−1 (p<0.005), 1.7×103·mL−1 versus 0.24×103·mL−1 (p<0.01) and 1.3×103·mL−1 versus 0.02×103·mL−1 (p<0.05), respectively; table 3⇑). There was no difference in the number of macrophages and total cell counts in asthmatic and normal subjects.
Measurements of CCL1 in BALF
Measurements of CCL1 with ELISA showed that levels of this cytokine were significantly elevated in BALF from asthmatics compared with normals (median (range) 193 (108–449) pg·mL−1 versus 30 (15–55) pg·mL−1, p<0.001; fig. 1a⇓).

a) Levels of immunoreactive CC chemokine ligand (CCL)1 in the bronchoalveolar lavage fluid of asthmatic patients and normal subjects after correcting for the initial two-fold concentration. Horizontal line represents the median. b) Densitometric analysis of CCL1 mRNA expression normalised to reduced glyceraldehyde phosphate dehydrogenase (GAPDH) in asthmatics and normals. c) CCL1 mRNA expression in bronchoalveolar lavage cells from normal and asthmatic subjects; blots correspond with results for normals and asthmatics given in b). #: CCL1 gene expression in PMA-stimulated U937 cells as a positive control (+). ***: p<0.001.
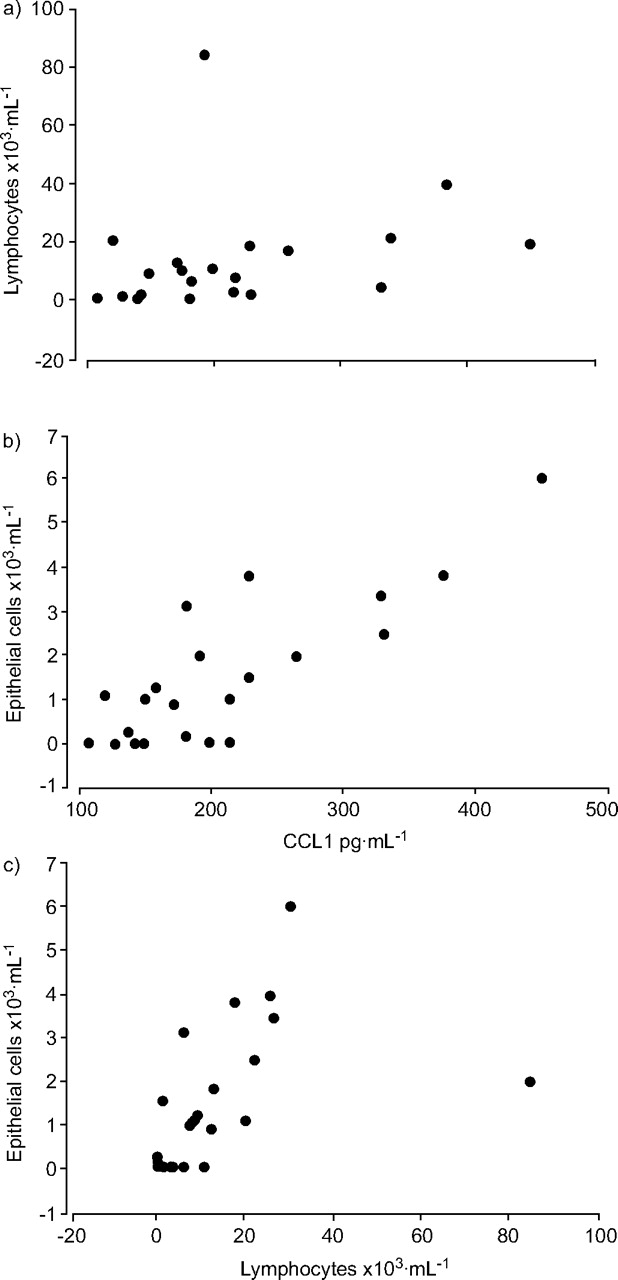
There was a trend towards a correlation between CCL1 levels and lymphocyte numbers in BALF, although this just failed to reach statistical significance (r = 0.4, p = 0.08; fig. 2⇓a). Interestingly, there was a significant correlation between CCL1 levels and epithelial cells (r = 0.73, p = 0.001; fig. 2b⇓) and between these cells and lymphocytes (r = 0.6, p = 0.02; fig. 2c⇓). However no significant correlation was seen between CCL1 and any other infiltrating leukocytes, including macrophages and eosinophils. In addition, a significant correlation between concentrations of CCL1 and bronchial hyperreactivity (provocation concentration of methacholine causing a 20% fall in FEV1 (PC20)) was not seen.

Correlations between a) CC chemokine ligand (CCL)1 levels and lymphocyte numbers (r = 0.4, p = 0.08), b) CCL1 and epithelial cells (r = 0.73, p = 0.001) and c) Iymphocytes and epithelial cells (r = 0.6, p = 0.02).
Using SDS-PAGE and Western blot CCLI was detected in BALF derived from asthmatics but not control subjects (fig. 3⇓).

Western blot for CC chemokine ligand 1. Lane 1: molecular weight markers; lanes 2 and 3: bronchoalveolar lavage fluid (BALF) derived from asthmatics; lanes 3 and 4: BALF derived from control subjects. kD: kilodaltons.
CCL1 mRNA in BAL cells
CCL1 mRNA was investigated in BAL cells derived from normal and asthmatic subjects. Constitutive CCL1 mRNA was observed in BAL cells from normal subjects. However, increased CCL1 mRNA expression was seen in BAL cells derived from asthmatic compared with normal subjects (fig. 1b⇑).
Biochemical and biological characterisation of lymphocyte chemotactic activity from BALF
To investigate lymphocyte chemotactic activity (LCA), BAL from five asthmatic subjects were combined (250 mL of BALF). Subsequent analysis of BALF using sequential CCL1 affinity column and reverse-phase chromatography allowed partial purification of a peak of LCA (fig. 4⇓). Using ELISA it was demonstrated that this peak of activity contained CCL1. Similar LCA profiles were obtained in three separate experiments using different pooled BALF samples. Incubation of BAL CCL1 with a neutralising anti-CCL1 antibody (MAB 272) blocked the BAL-LCA (fig. 5b⇓). For chemotaxis assays CCR8 was induced in CD4 lymphocytes by incubating these cells with IL-2 for 5 days (fig. 5a⇓).
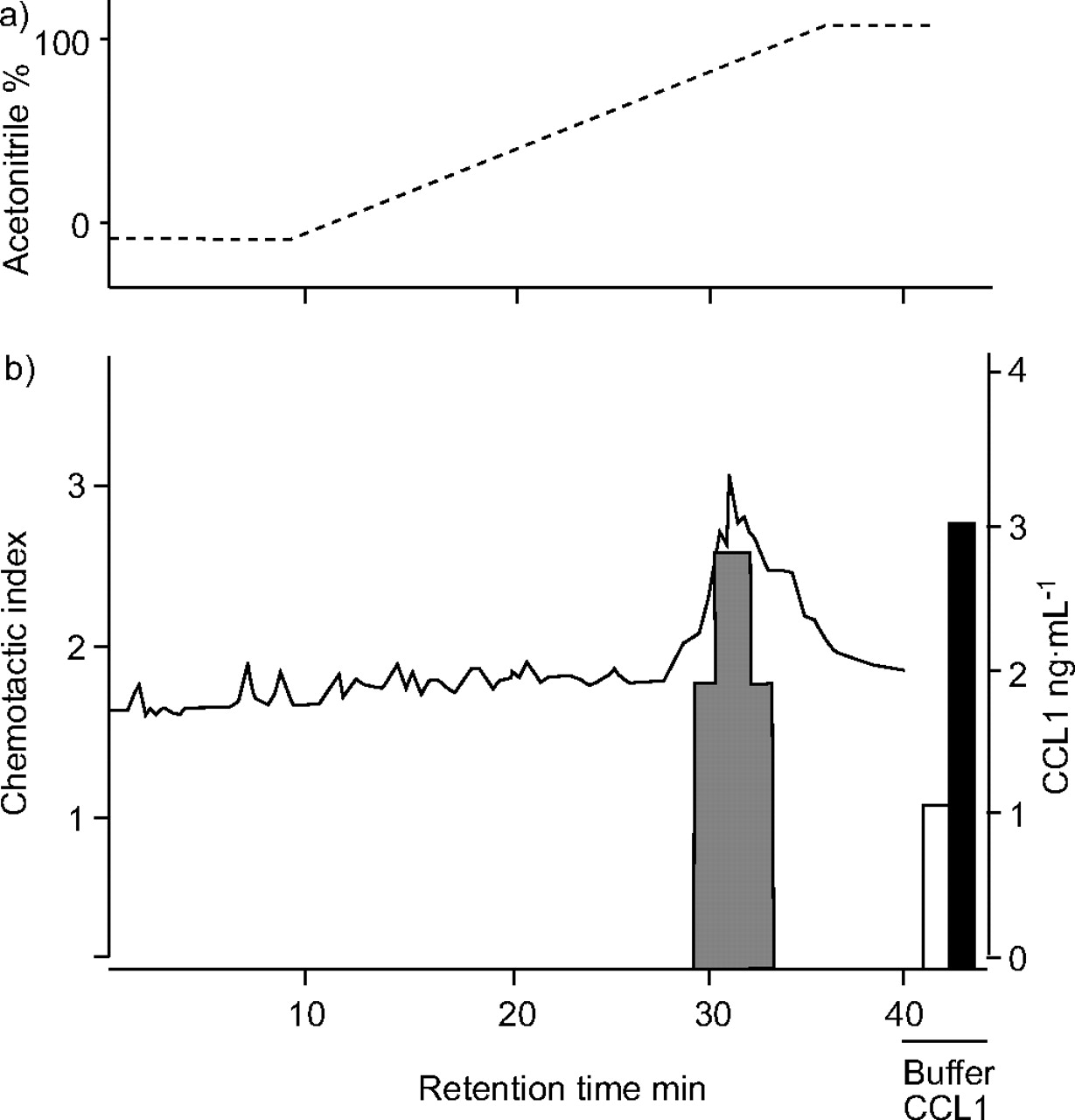
Lymphocyte chemotactic activity (LCA) from bronchoalveolar lavage fluid after sequential CC chemokine ligand (CCL)1/I-309 affinity chromatography and reverse-phase chromatography resolved into a peak of LCA (▒). CD4 chemotaxis in response to individual fractions is given as a chemotactic index. Negative control (buffer; □) per definition gave a chemotactic index of 1. Recombinant CCL1 (▪) was used as a positive control. -----: acetonitrile gradient; –––: CCL1 ELISA measurement.
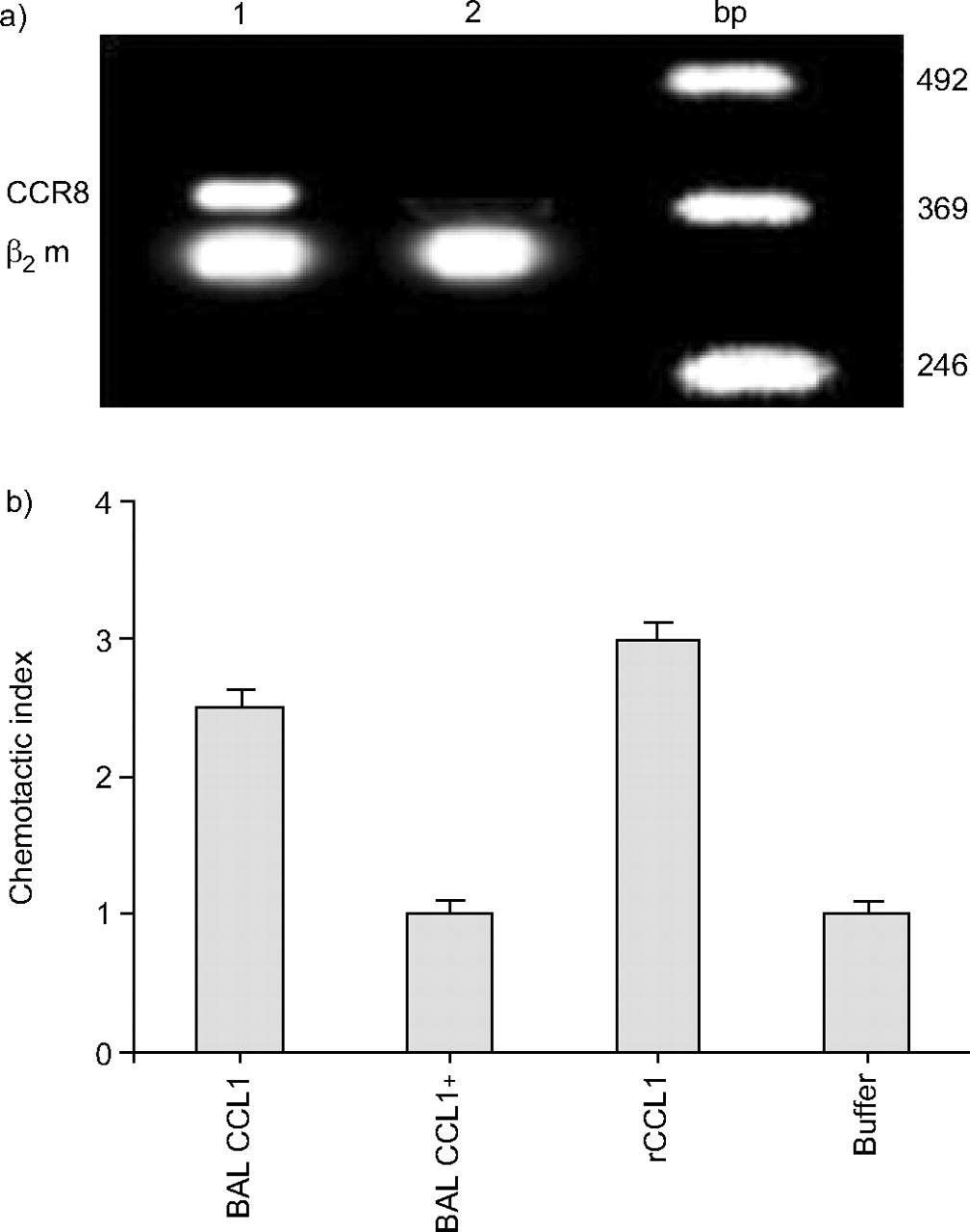
a) Interleukin-2-stimulated CD4+ cells express CC chemokine receptor 8 (CCR8; lane 1). Nonstimulated lymphocytes do not express this receptor (lane 2). b) Neutralisation of lymphocyte chemotactic activity (LCA) of bronchoalveolar lavage (BAL) CC chemokine ligand (CCL)1 (mean of three separate experiments). BAL CCL1 (5 ng·mL−1) was incubated with 1 µg of the monoclonal antibody 272 (neutralising AB to CCL1) for 15 min before the chemotactic assay. Chemotactic activity is shown as a chemotactic index. Recombinant (r)CCL1 (10−8 M) was used as a positive control. Negative control (buffer) per definition, gave a chemotactic index of 1. bp: base pair; β2 m: β2 macroglobulin.
CCL1 immunoreactivity in bronchial biopsies
To investigate potential cellular sources of CCL1, the technique of immunohistochemistry was applied to bronchial biopsies. CCL1 immunoreactivity was localised predominantly to the airway epithelium (fig. 6a⇓ and c). Similar findings were observed in biopsies derived from four different asthmatics. In contrast, CCL1 immunoreactivity was not seen in bronchial biopsies derived from normal subjects (data not shown). To further investigate this cytokine, nasal polyps were also analysed using immunohistochemistry. Interestingly, CCL1 immunoreactivity was also localised to the airway epithelium of nasal polyps (fig. 6d⇓).
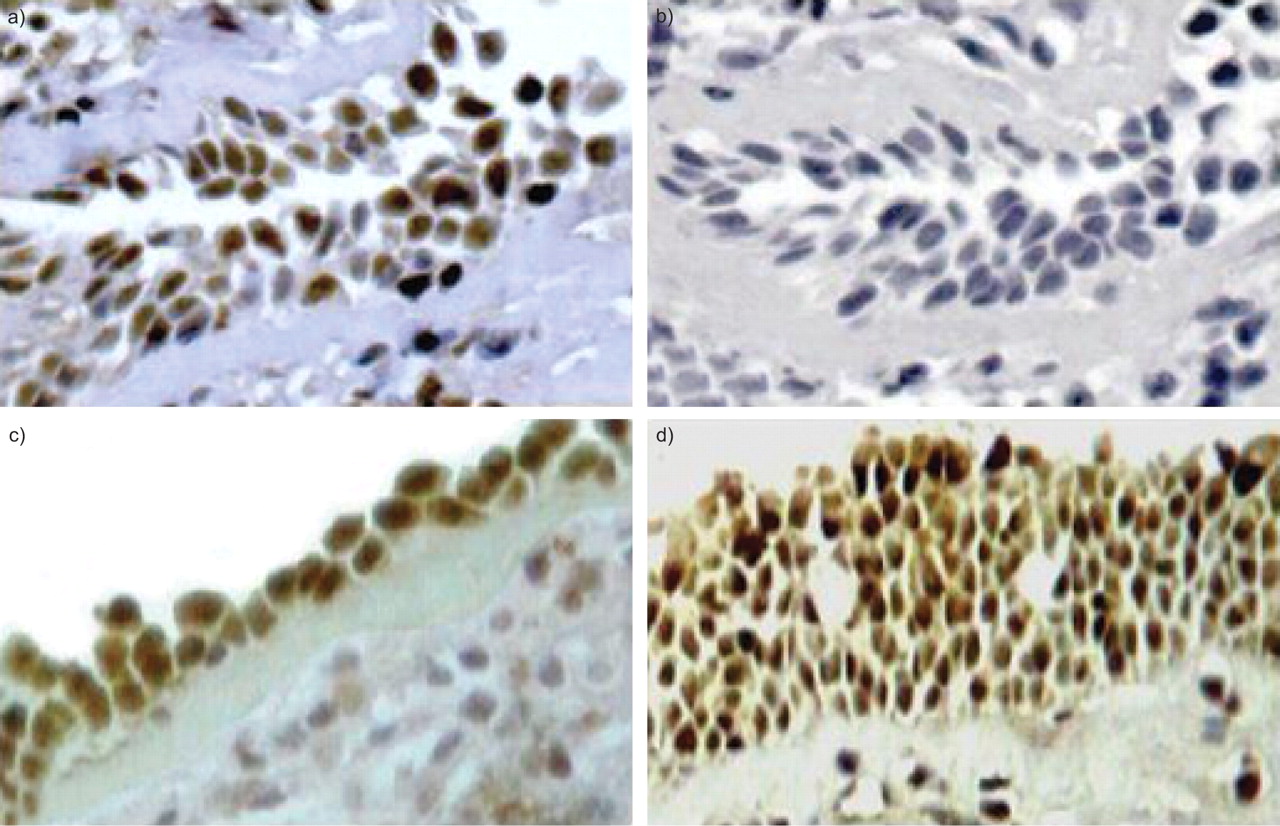
a and c) Localisation of immunoreactive CC chemokine ligand (CCL)1 to the airway epithelium of bronchial biopsies derived from two asthmatics. b) Negative control of a bronchial biopsy (instead of the anti-CCL1/I-309 antibody the isotype control antibody goat immunoglobulin G 15256 was used). d) Immunoreactive CCL1 to the airway epithelium of a nasal polyp. See Methods section for further information on immunohistochemistry.
Bronchial airway epithelial cells release CCL1 in vitro
Having demonstrated CCL1 immunoreactivity localised to the bronchial airway epithelium, the present authors next investigated whether primary BAEC release this chemokine upon stimulation with either Th1 (IFN-γ) or Th2 cytokines (IL-4 and -13). Measurements of CCL1 in the culture supernatant showed that these cells produced low levels of CCL1 constitutively. Among the cytokines, IL-4 was the strongest stimulus for the release of CCL1, followed by IL-13 and IFN-γ (fig. 7⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Levels of immunoreactive CC chemokine ligand 1 in the culture supernatants (n = 4) derived from both the cytokine-stimulated primary bronchial airway epithelial cells (BAEC) as well as nonstimulated BAEC supernatants (control). Error bars represent median values. IL: interleukin; IFN: interferon. #: p<0.02; **: p<0.01; ***: p<0.001.
DISCUSSION
A novel finding of the present study is the demonstration that CCL1 is released in high concentrations into the airway epithelial lining fluid of asthmatic patients. Interestingly, BAL CCL1 was found to activate CD4 cells as assessed in chemotactic assays. With immunohistochemistry, CCL1 was localised predominantly to the bronchial airway epithelium. Human bronchial epithelial cells were found to release high concentrations of CCL1 upon cytokine stimulation in vitro.
CCL1 was originally characterised as a chemotatic factor for monocytes, and subsequently shown to be a potent Th2 lymphocyte attractant 9–11. As Th2 lymphocytes play an important role in asthma, this cytokine has attracted major attention in allergic inflammation. The murine homologue of CCL1/I-309 is CCL1/T-cell activation-3 30, 31. CCL1/T-cell activation-3 has been implicated in several models of inflammatory diseases, including inflammatory bowel disease, experimental allergic encephalomyelitis and allergic asthma 20, 32, 33. To date, however, the role of this cytokine in asthma remains to be shown.
This is the first study to demonstrate that CCL1 is released into the airways of asthmatic patients. Indeed, levels of this cytokine were six-fold greater in asthmatics compared with normals. A previous group failed to detect CCL1 in BALF from asthmatics exposed to allergen challenge, but detected other chemokines such as CCL17 (TARC), CCL22 (MDC) and CXCL10 (interferon-inducible protein-10) 18. Their failure to detect CCL1 could be explained by the fact that CCL1 was investigated in neat BALF. It's also possible that the antibodies used in their ELISA were not sensitive enough to detect CCL1. In the present study, BALF was concentrated 2–10 times and it was demonstrated that 2×concentrated BALF was good enough to detect CCL1. The presence of CCL1 in BALF was further confirmed by Western blot. It is well established that BALF is a dilution of airway epithelial lining fluid of 1:100 to 1:200 34–37. A finding of 193 pg·mL−1 thus reflects a concentration of 3.8–38 ng·mL−1 in airway lining fluid. Expressing cytokines levels per mL of BALF after correction for the initial concentration is a well-established method 25, 37–39, although few researchers correct inflammatory mediators such as eosinophil cationic protein, IgE and other Igs according to either albumin or urea concentrations 40, 41. CCL1 levels were also calculated according to albumin concentrations in BALF. However, there was no statistical difference from that expressed per mL of BALF (data not shown). Interestingly, a European Respiratory Society Task Force report 42 concludes that results of acellular components should be expressed as amounts per mL in order to facilitate comparison of data from different works until a reliable external marker can be defined. Previous studies have detected other CD4-activating chemokines, including CCL17 and CCL22 in stable asthmatics 18, 37. The present authors therefore suggest that CCL1, in concert with other CD4-activating chemokines, such as CCL22 and CCL17, may be involved in the recruitment of lymphocytes into the airways of asthmatic patients.
In order to further investigate CCL1, the gene encoding of this cytokine was studied. Increased CCL1 mRNA expression was seen in BAL cells from asthmatic patients compared with that observed in cells obtained from normal subjects. This finding further suggests that CCL1 is highly regulated in asthma.
Having unequivocally shown increased immunoreactive CCL1 and CCL1 gene upregulation, the present authors wondered whether biologically active CCL1 could be identified in BALF. The present authors have previously shown that BALF constitutes an excellent tool to study cell attractants 25, 27. Indeed, separation of BALF by sequential CCL1 affinity chromatography and reverse phase-chromatography enabled the identification of a peak of LCA, which, by using ELISA, was found to correspond to CCL1. The biological activity associated with these fractions could be abolished completely using a neutralising antibody in CCL1. Thus, the finding that BAL CCL1 activates CD4 cells expressing CCR8 in vitro (fig. 4⇑) together with the demonstration that CCL1 levels showed a trend towards a correlation with lymphocyte numbers in BALF suggest that when released locally, this cytokine may attract CD4 cells into the airways of asthmatics.
To investigate the cellular source of the CCL1 detected in BALF, an immunohistochemical staining of bronchial biopsies was performed. This allowed the present authors to demonstrate that CCL1 immunoreactivity was predominantly localised to the airway epithelium. Moreover, levels of CCL1 correlated with the number of airway epithelial cells in BALF and these cells correlated with lymphocytes. This suggests that during the inflammatory process airway epithelial cells may release CCL1 and this cytokine in turn may recruit lymphocytes into the airways of asthmatics. Interestingly, the number of airway epithelial cells in BALF from asthmatics was greater compared with that observed in BALF from control subjects and it is likely that they may have accounted for the increased CCL1 mRNA expression observed in BAL cells. Thus, it could be suggested that epithelial cells shed from the airways could be a major cell source of CCL1 in vivo. To further investigate CCL1 production, primary BAEC were stimulated with either Th1 or Th2 cytokines. Interestingly, IL-4 was found to be a strong stimulus for CCL1 release from BAEC, followed by IL-13 and IFN-γ. Both IL-4 and IL-13 are known to play an important role in allergic inflammation. These cytokines activate B-cells, inducing class switching to IgE and expression of both CD23 and class II major histocompatibility complex antigens 43, 44. Increased mRNA encoding IL-4 and IL-13 has been reported in both bronchial biopsies and BAL cells from asthmatics 45. Taken together these observations suggest that IL-4, IL-13 and IFN-γ may induce CCL1 release into asthmatic airways; however, further studies must investigate in more detail the production of this cytokine by BAEC.
To date, there are no in vivo studies showing the direct effect of CCL1 on normal airways. Thus, it is not known whether CCL1 induces bronchial hyperreactivity. The present study did not find a correlation between CCL1 levels and bronchial hyperreactivity (PC20 methacholine), which is not surprising. The present authors suggest that CCL1 induces recruitment of Th2 cells, which in turn may release cytokines such as IL-4, IL-5 and IL-13. These cytokines are potent inductors of bronchial hyperreactivity. Thus, it is tempting to suggest that neutralising the effects of CCL1/CCR8 may have a substantial impact on allergic disease. Indeed, a previous study has shown that the use of an anti-CCL1 antibody inhibits the eosinophil recruitment into the lung of sensitised mice 20. Similarly, Chensue et al. 21 demonstrated that CCR8-/- mice had impaired pulmonary eosinophilia although other authors have failed to reproduce these findings 22. Interestingly, increased CCR8 immunoreactivity, localised to Th2 cells, has been previously found in bronchial biopsies derived from asthmatics and its expression correlated with bronchial hyperreactivity 18. More recently, CCR8 has been reported to be expressed in both Th1 and Th2 skin lymphocytes 46. The development of small CCL1 antagonists with clinical efficacy will define the role of this cytokine in asthma. However, future studies must also investigate the effect of corticosteroids, the mainstay treatment of asthma, on CCL1 release.
In summary, the present study has demonstrated that asthmatics release increased concentrations of CC chemokine ligand 1 in bronchoalveolar lavage fluid. The levels of this cytokine were found to be associated with an increased number of lymphocytes in bronchoalveolar lavage fluid. Bronchoalveolar lavage CC chemokine ligand 1-induced migration of CD4 cells expressing CC chemokine receptor 8 in a chemotactic assay. Using immunohistochemistry, CC chemokine ligand 1 immunoreactivity was localised predominantly to the bronchial airway epithelium. Interestingly, both CC chemokine ligand 1 levels and lymphocyte numbers correlated with airway epithelial cell numbers in bronchoalveolar lavage fluid. Moreover, bronchial airway epithelial cells were found to release CC chemokine ligand 1 in vitro. Taken together these findings suggest that CC chemokine ligand 1 may play a role in the recruitment of lymphocytes that characterise bronchial asthma.
Acknowledgments
The authors are grateful to B. Dávila for recruiting some of the volunteers for the present study.
- Received November 23, 2004.
- Accepted March 10, 2006.
- © ERS Journals Ltd
References










