





Abstract
To clarify the discrepancy between increasing resistance and conservative clinical effects of macrolides on macrolide-resistant Streptococcus pneumoniae, the authors evaluated the effects of sub-minimum inhibitory concentrations of macrolides on pneumolysin.
In vitro, S. pneumoniae was incubated with 1, 2 and 4 µg·mL−1 of clarithromycin (CLR) and azithromycin (AZM) for 8 h. Western blot analysis and haemolytic assay were performed to examine the production and activities of pneumolysin. In vivo, mice were infected with S. pneumoniae intra-nasally and treated with CLR (40 or 200 mg·kg−1 twice daily) or AZM (40 or 200 mg·kg−1 once daily) orally for 7 days. After 72 h post-infection, western blot analysis was performed to examine pneumolysin production in lungs. Survival rates were observed for 10 days.
In vitro, every concentration of macrolide inhibited pneumolysin production more than the control. CLR (2 and 4 µg·mL−1) and AZM (4 µg·mL−1) reduced the pneumolysin activities more than the control. In vivo, macrolides (200 mg·kg−1) reduced pneumolysin in murine lungs more than the control. CLR (40 and 200 mg·kg−1) and AZM (200 mg·kg−1) improved the survival rates more than the control.
The study results show that sub-minimum inhibitory concentrations of macrolides reduced pneumolysin. This might be related to the effectiveness of macrolides against pneumonia caused by high-level macrolide-resistant Streptococcus pneumoniae. Further investigations are necessary to evaluate the effects of macrolides on macrolide-resistant Streptococcus pneumoniae.
- Azithromycin
- clarithromycin
- macrolide-resistant Streptococcus pneumoniae
- pneumolysin
- sub-minimum inhibitory concentrations
The most prevalent community-acquired respiratory tract pathogen, Streptococcus pneumoniae, causes ≥20% of all cases of community-acquired pneumonia 1. Traditionally, the guidelines for treatment of outpatients with community-acquired pneumonia recommend macrolides as firstline treatment for patients with no comorbidity 2. However, widespread use of macrolides leads to an increase in macrolide-resistant S. pneumoniae (MRSP) 3.
Macrolide resistance in S. pneumoniae occurs as a result of two mechanisms: an alteration of the macrolide binding site by methylation in the 23S rRNA, encoded by erm B, and efflux pump, encoded by mef E/mef A, via which bacteria expel macrolides. Phenotypes resulting from the expression of the erm B gene generally exhibit high levels of macrolide resistance. Furthermore, the level of resistance resulting from the expression of the efflux pump is generally less than that produced by target site modification. Recent data from the USA has revealed that up to 27.9% of pneumococci are resistant to macrolides. Ultimately, up to 29.0% of MRSP are mediated by modification of the erm B gene 4.
Though a number of studies have related macrolide resistance in S. pneumoniae to macrolide failure 5, 6, other authors have experienced difficulties in finding a significant correlation 7. Nuermberger and Bishai 8 discussed the uncertainties of the efficacy of macrolides against MRSP. Ewig et al. 9 looked at patients hospitalised for community-acquired pneumococcal pneumonia and reported that there was no significant difference in mortality rate between patients infected with macrolide-susceptible versus macrolide-non-susceptible S. pneumoniae (12% versus 7%, respectively). Anzueto et al. 10 also reported that clarithromycin (CLR) had a good effect in the treatment of outpatients with acute exacerbations of chronic bronchitis caused by CLR-resistant pneumococci.
Some recent reports have shown the effects of sub-minimum inhibitory concentrations (sub-MIC) of macrolides on bacterial virulence factors 11. For example, sub-MIC of azithromycin (AZM) inhibited exotoxin A, total protease, elastase and phospholipase C production by Pseudomonas aeruginosa without affecting the growth of P. aeruginosa 12.
The cholesterol-dependent toxin pneumolysin (PLY) is one of the most important virulence factors of S. pneumoniae. PLY has multiple distinct functions that augment the virulence of S. pneumoniae during the early pathogenesis of invasive pneumococcal infection. PLY can disrupt epithelial tissues that form a mechanical barrier, and allow S. pneumoniae to penetrate into the blood stream. Studies have shown that PLY alone could reproduce the symptoms of the pneumococcal disease in lungs 13. Subsequent studies demonstrated that mutant pneumococci unable to produce PLY generate much less inflammation, delay bacteraemia, and limit multiplication of bacteria within the lung 14–16. Thus, PLY plays an important role during pneumococcal pneumonia.
To clarify the discrepancy between in vitro resistance and the conservative clinical effects of macrolides, this study examines the effects of sub-MIC of CLR and AZM on the PLY of high-level MRSP in vitro and in vivo.
MATERIALS AND METHODS
Micro-organism
A clinical isolate of MRSP was obtained from Nagasaki University School of Medicine (Nagasaki, Japan). NU4471 (minimum inhibitory concentration (MIC) of penicillin G, 2 µg·mL−1; serotype 19) was used in this experiment. The presence of resistance mechanisms for erm B and mef A genes was analysed using PCR 17. The bacterium was stored at −80°C until use.
Antimicrobial agents
CLR (Taisho Toyama Pharmaceutical, Tokyo, Japan) and AZM (Pfizer Japan Inc., Tokyo, Japan) were kindly supplied by the indicated companies. For the antimicrobial susceptibility test, each macrolide was dissolved in ethanol and diluted further with sterile water. For murine experiments, CLR powder was dissolved in 0.5 M HCl and diluted to a final concentration of 30 mg·mL−1 with sterile water, then sonicated for 30 min. AZM powder was suspended to a final concentration of 30 mg·mL−1 with 0.1% carboxymethyl cellulose. Both macrolides were prepared as recommended by the manufacturer. Macrolide stock solutions were diluted with sterile water immediately prior to use to yield the appropriate drug concentrations in a final administration volume of 0.3 mL. Drug solutions were stored at 4°C for no more than 72 h.
Antimicrobial susceptibility test
The MIC of each antibiotic was determined by the broth dilution method using Muller-Hinton broth (Difco Laboratories, Detroit, MI, USA) supplemented with 5% lysed horse blood (Nippon Biotest Laboratories Inc, Tokyo, Japan). Microtitre plates, containing 5.0×105 colony-forming units (cfu)/well of NU4471 were incubated with antibiotics at 35°C for 18 h, and the lowest concentration of the drug that prevented visible growth was considered the MIC 18.
Western blot analysis
Suspensions of NU4471 (3.0×106 cfu·mL−1) in cell culture medium (D-MEM/F-12: Invitrogen Corp., Carlsbad, CA, USA) with 1, 2 and 4 µg·mL−1 of CLR and AZM or sterile water (control) were incubated at 37°C for 8 h at late-log phase (n = 4) 19. Thereafter, pneumococci were lysed with protease inhibitor cocktails (Nacalai Tesque, Kyoto, Japan) with a sonicator (Ultrasonic Disruptor UD-201: Tomy Seiko, Tokyo, Japan) 20. The western blot procedure for PLY was then performed as described previously 21. Sample proteins (30 µg) of whole lysates were separated using SDS-PAGE (sodium dodecyl sulphate-polyacrylamide gel electrophoresis) and transferred onto nitrocellulose membranes. Blots were blocked and successively incubated with monoclonal murine antibodies to recombinant PLY (NCL-SPNm: Novocastra Laboratories, Newcastle-upon-Tyne, UK) and peroxidase-conjugated goat anti-mouse antibodies (Bio-Rad, Richmond, CA, USA). The reaction was developed with the ECL+ enhanced chemiluminescence western blotting detection reagents and exposed to Hyperfilm ECL (both from Amersham Biosciences UK, Buckinghamshire, UK). PLY was detected as a band at 53 kDa. Densitometry of the films was performed using a software program. Viable counts (cfu·mL−1) were also determined after 8-h incubation by plating appropriate dilutions on blood agar plates and incubated at 37°C for 18 h. The lowest level of detectable cfu·mL−1 was 50 cfu·ml−1.
Haemolytic assay
Suspensions of NU4471 with 1, 2 and 4 µg·mL−1 of CLR and AZM or sterile water (control) were incubated at 37°C for 8 h and lysed with a sonicator as described above (n = 4). The haemolytic activity was then determined using 1% (volume/volume) suspensions of fresh horse erythrocytes (washed three times) in PBS, as described previously 19.
Laboratory animals
For the study, 5-week-old female CBA/J specific-pathogen-free mice (body weight 16–20 g) were purchased from Charles River Japan (Yokohama, Japan). The CBA/J mouse model of S. pneumoniae has been described previously 22. All experiments were conducted taking into consideration the guidelines of the Laboratory Animal Center for Biomedical Research, Nagasaki University School of Medicine.
Experimental murine model of pneumococcal pneumonia
NU4471 was cultured on horse blood agar (Muller-Hinton ΙΙ agar: Becton, Dickinson and Company, Sparks, MD, USA, horse blood from same manufacturer) at 37°C for 24 h. It was then scraped and suspended in brain–heart infusion broth (Becton, Dickinson and Company) mixed with horse serum (Invitrogen Corp.) and cultured at 37°C for 6 h at mid-log phase. Bacteria were then harvested by centrifugation at 800×g for 3 min. The organisms were resuspended in normal saline and the final number of bacteria prepared was ∼107 cfu·ml−1, as determined by optical density. Infection was induced by intra-nasal inoculation of 0.05 mL bacterial suspension, containing about 5×105 cfu·mouse−1, into anesthetised mice (pentobarbital sodium, 40 mg·kg−1 i.p.) 23, 24.
Treatment regimens for the survival study
Treatment was initiated at 12 h post-infection. The regimen included: CLR at 200 mg·kg−1 and saline (control) administered 12, 24, 36, 48 and 60 h post-infection; AZM at 200 mg·kg−1 administered 12, 36 and 60 h post-infection 25. Antibiotics were dispensed orally. Mice (four in each group) were killed by cervical dislocation at 72 h post-infection. The lungs were dissected out under aseptic conditions and suspended in saline (50 µL).
Bacteriological examination and immunoprecipitation of lungs for western blot analysis
For bacteriological examination, the lungs were homogenised just after death using a Polytron homogeniser (PT3000; Kinematica, Littau-Lucerne, Switzerland). The number of bacteria in the lungs was determined as described previously 23. Thereafter, 1 mL of lysis buffer was added to the lung lysates, incubated on ice for 10 min, and centrifuged at 12,000×g at 4°C as described previously 26. Lung extracts were isolated and 8 mg of proteins were rotated for 1 h at 4°C with 60 µg of monoclonal murine antibodies to recombinant PLY (NCL-SPNm) cross-linked to protein G beads (Amersham Biosciences AB, Uppsala, Sweden). The beads were washed four times with 1 mL of lysis buffer and boiled for 5 min in SDS-denaturing sample buffer. Finally, western blot analysis was performed as described above.
Treatment regimens for study of survival rates
Treatment was initiated at 12 h post-infection and continued for 7 days. The regimens included: CLR at 40 or 200 mg·kg−1 twice daily; AZM at 40 or 200 mg·kg−1 once daily; and saline (control) twice daily. Seven mice were included in each treatment group. Antibiotics were administered through oral gavage. The survival rates were determined daily over a 10-day post-infection period 25.
Statistical analysis
Data are expressed as mean±sem. Differences in the protein synthesis and haemolytic activities of PLY or the number of viable bacteria between macrolide-treated groups and the control group were evaluated using ANOVA. Survival analysis was performed using the log-rank test, and survival rates were calculated via the Kaplan–Meier method. A p-value <0.05 was considered statistically significantly.
RESULTS
Minimum inhibitory concentration of macrolides for Streptococcus pneumoniae NU4471
Both of the MICs of CLR and AZM were ≥256 µg·mL−1. The presence of both erm B and mef E/A genes in NU4471 was confirmed by PCR (data not shown).
Effect of macrolides on pneumolycin protein synthesis
Figure 1a⇓ shows the representative results of western blot analysis of the suppressing effects of sub-MIC (1, 2 and 4 µg·ml−1) of CLR and AZM on the protein synthesis of PLY by NU4471. After 8 h of incubation, the production of PLY was reduced in the presence of 1, 2 and 4 µg·mL−1 of CLR and AZM. Moreover, the inhibitory effect of 4 µg·mL−1 of CLR on the production of PLY was better than that of 4 µg·mL−1 of AZM. There was no difference in the number of bacteria recovered in the presence or absence of macrolides after 8 h of incubation (fig. 1b⇓).

a) The representative results of western blot analysis of the suppressing effects of sub-minimum inhibitory concentrations of clarithromycin (CLR) and azithromycin (AZM) on the protein synthesis of pneumolysin by NU4471 after 8 h of incubation. b) The number of colony-forming units (cfu) in media treated with saline (control), CLR at 1, 2 and 4 µg·mL−1 and AZM at 1, 2 and 4 µg·mL−1 were 8.33±0.03, 8.27±0.14, 8.33±0.11, 8.49±0.08, 8.20±0.12, 8.31±0.08 and 8.23±0.11 log10 cfu·mL−1, respectively. Data present the mean±sem of four experiments.
Effect of macrolides on haemolytic activities of pneumolysin
The effects of sub-MIC (1, 2 and 4 µg·mL−1) of CLR and AZM on the haemolytic activities of PLY by NU4471 are shown in figure 2⇓. Data reflect absorbance at 541 nm. After 8 h of incubation, the haemolytic activities were significantly lower in the presence of CLR (2 and 4 µg·mL−1) and AZM (4 µg·mL−1) than control (p<0.0001). The inhibitory effect of 4 µg·mL−1 of CLR on haemolytic activity was better than that of 4 µg·mL−1 of AZM (p<0.05).

Effects of sub-minimum inhibitory concentrations of clarithromycin (CLR) and azithromycin (AZM) on the haemolytic activities of pneumolysin (PLY) by NU4471. After 8 h of incubation, the haemolytic activity of PLY treated with saline (control), CLR at 1, 2 and 4 µg·mL−1 and AZM at 1, 2 and 4 µg·mL−1 were 1.07±0.01, 0.95±0.04, 0.50±0.08, 0.24±0.10, 1.03±0.03, 1.00±0.03 and 0.54±0.03, respectively. Data present the mean±sem of four experiments. #: p<0.0001 versus control; *: p<0.05 versus AZM at 4 µg·mL−1.
Effect of macrolides on the production of pneumolysin in murine lungs
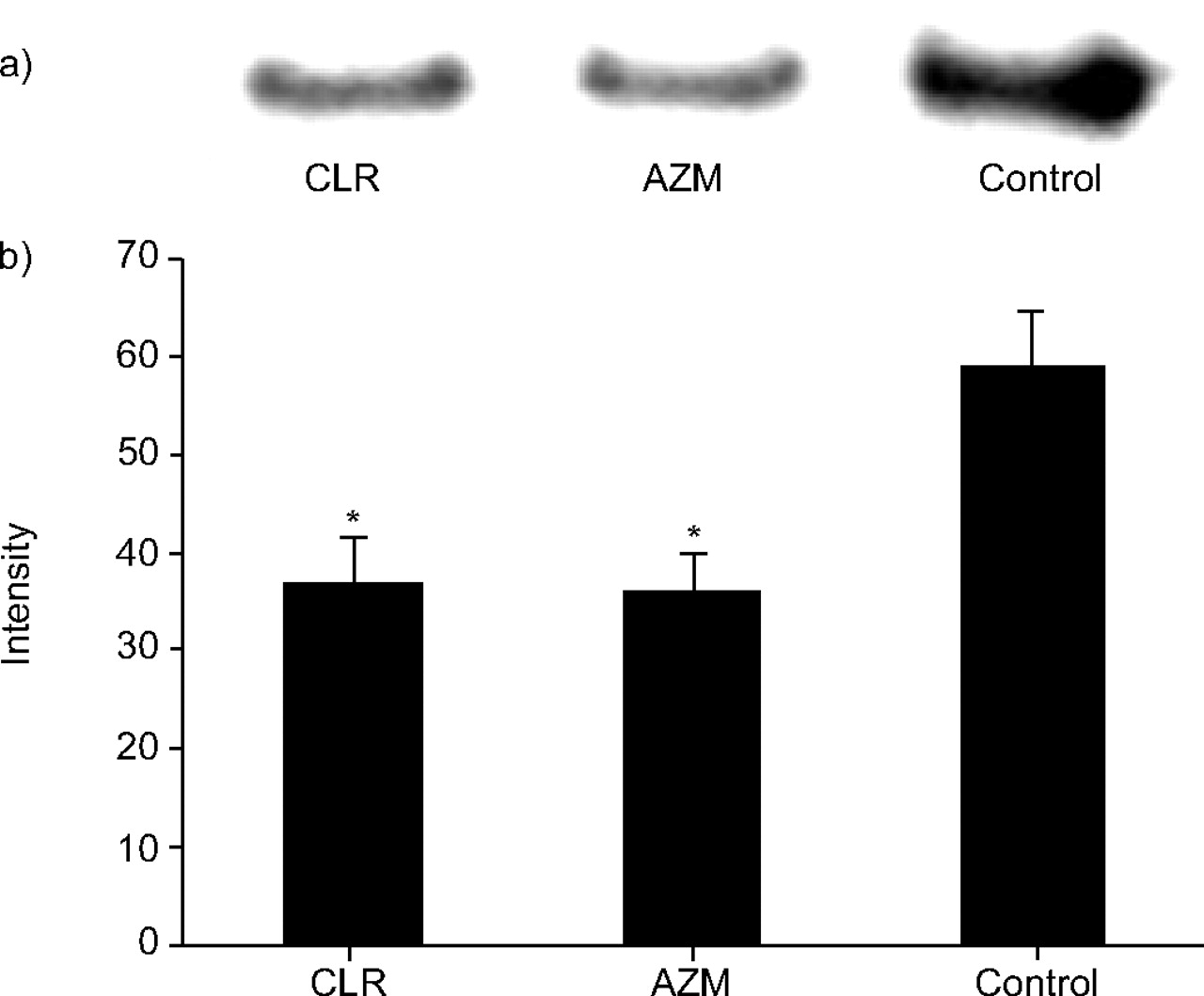
Figure 3⇓ shows western blot analysis of the inhibitory effects of CLR at 200 mg·kg−1 twice daily, AZM at 200 mg·kg−1 once daily and saline (control) twice daily on the production of PLY by NU4471 in murine lungs. Treatment with CLR at 200 mg·kg−1 and AZM at 200 mg·kg−1 significantly reduced the production of PLY in murine lungs compared with the control (p<0.05).
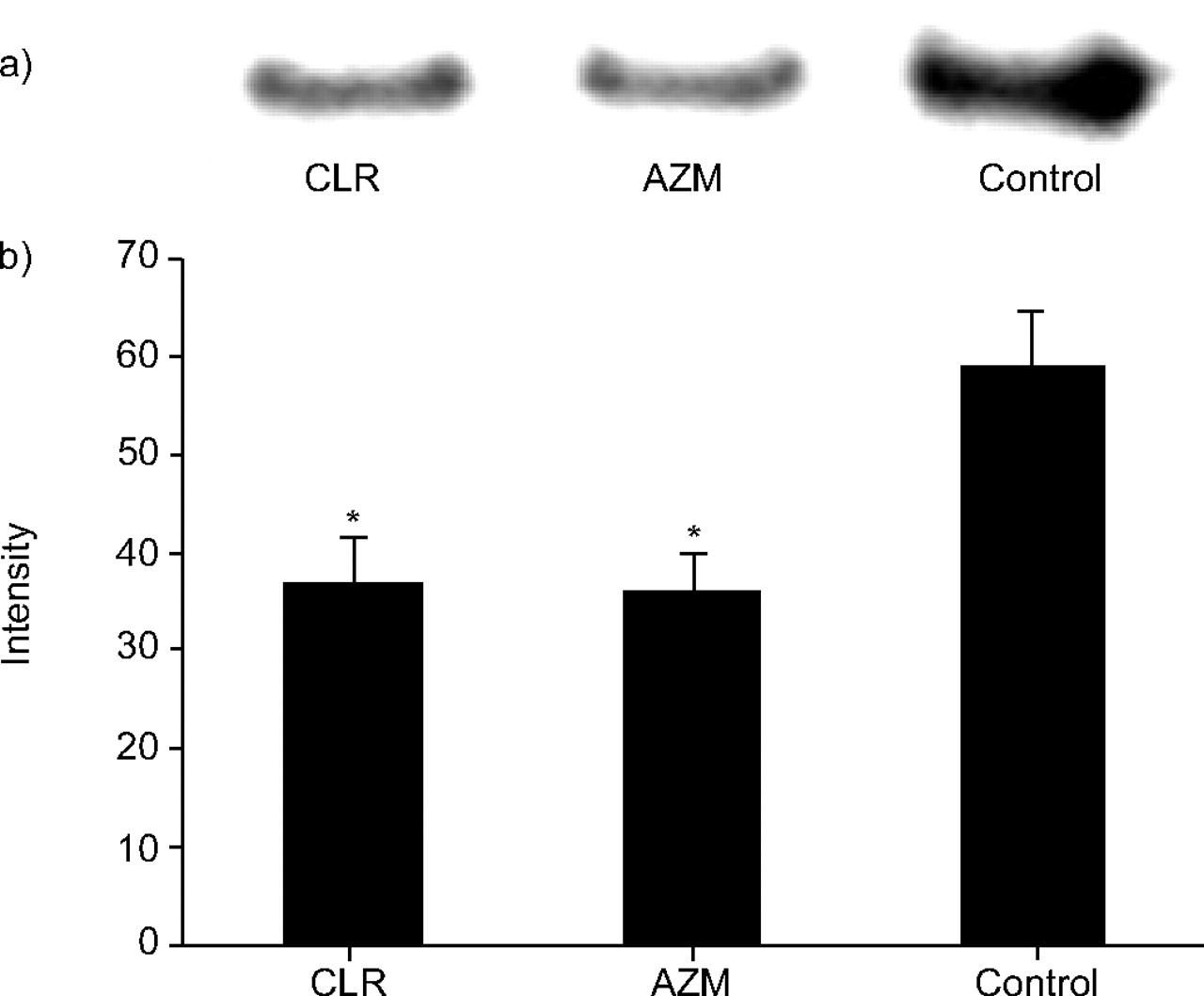
a) Representative results of western blot analysis of the inhibitory effects of clarithromycin (CLR) at 200 mg·kg−1 twice daily, azithromycin (AZM) at 200 mg·kg−1 once daily and saline (control) twice daily on the production of pneumolycin by NU4471 in murine lungs. b) Signal intensity of each band treated with CLR at 200 mg·kg−1, AZM at 200 mg·kg−1 and control. Data present mean±sem of four mice. *: p<0.05 versus control.
The numbers of viable bacteria in mice lungs treated with CLR, AZM and saline were 7.21±0.27, 7.68±0.48 and 7.66±0.18 log10 CFU·lung−1, respectively. There was no difference in the total CFU of viable bacteria in the murine lungs recovered after treatment with CLR, AZM and saline.
Survival rates
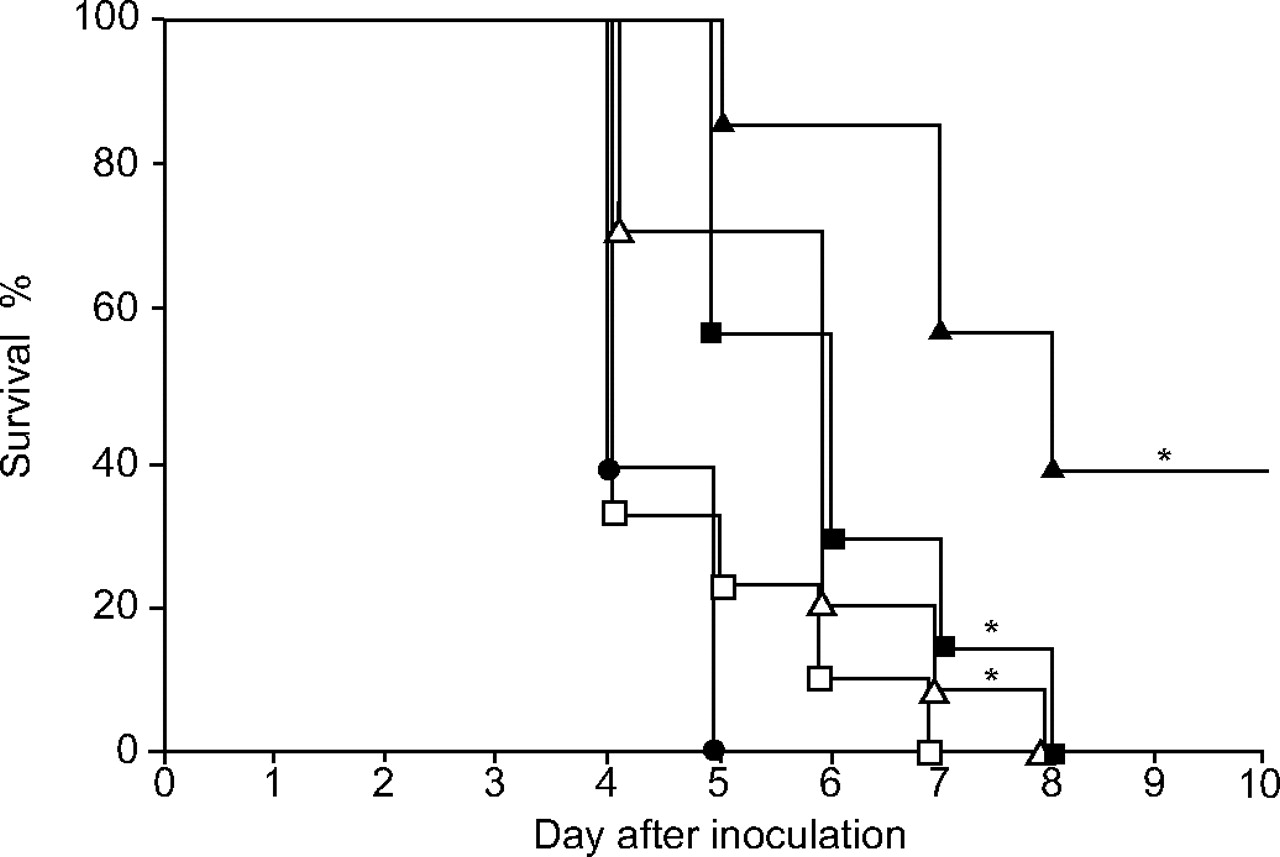
The survival rates of mice infected with NU4471 and treated with CLR at 40 or 200 mg·kg−1 twice daily, AZM at 40 or 200 mg·kg−1 once daily and saline (control) twice daily are shown in figure 4⇓. Treatment with CLR at 40 and 200 mg·kg−1 and AZM at 200 mg·kg−1 significantly improved the survival rates compared with the control (p<0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Kaplan–Meier survival curves of mice (seven in each group) infected with NU4471 and treated with clarithromycin (CLR; two different doses), azithromycin (AZM; two different doses) or control (saline) for 7 days. •: mice treated with saline; □: mice treated with AZM at 40 mg·kg−1; ▵: mice treated with AZM at 200 mg·kg−1; ▪: CLR at 40 mg·kg−1; ▴: CLR at 200 mg·kg−1. *: p<0.05 versus control.
DISCUSSION
The present study has shown the effects of sub-MIC of CLR and AZM on PLY of high-level MRSP in vitro and in vivo.
In vitro, sub-MIC of CLR and AZM inhibited protein synthesis and haemolytic activities of PLY of high-level MRSP. In addition, the PLY-suppressive activity of CLR was greater than that of AZM. In the preliminary study, the inhibitory effect of CLR was also greater than that of erythromycin and josamycin (data not shown). A macrocyclic lactone ring attached to two or more sugar moieties characterises the structure of macrolides. It is possible that the difference in the potency is due to the difference in the chemical configuration and hence the ability to cross membrane barriers of pneumococci or binding to the 23S rRNA.
Previous studies suggested that sub-inhibitory levels of macrolides could inhibit bacterial virulence factors in vitro 11, 12. Lagrou et al. 19 also reported that sub-MIC of erythromycin reduced the haemolytic activity of PLY by MRSP. Spreer et al. 21 reported that in comparison to standard therapy with ceftriaxone, protein synthesis inhibitors such as rifampin and clindamycin reduced the release of PLY by S. pneumoniae in human cerebrospinal fluid during therapy of pneumococcal meningitis. The present results are in agreement with those studies. Tsui et al. 27 reported that, at sub-MIC, members of the macrolide-lincosamide-streptogramin group modulate specific groups of bacterial promoters. Although the mechanism of transcriptional modulation is not known, these modulation may lead to the inhibition of PLY gene expression at sub-inhibitory concentrations of macrolides.
It has been shown that in vivo, macrolides reduce the production of PLY in murine lungs infected with high-level MRSP. In the present study, the doses of CLR and AZM were chosen to achieve target parameters similar to those observed in healthy humans 25. Nevertheless, neither CLR nor AZM reduced the viable number of pneumococci lower than control in murine lungs. This finding suggests that the concentrations of CLR and AZM in the lungs were below MIC levels. Therefore, CLR and AZM reduced the PLY in murine lungs even at sub-MIC.
PLY is a multifunctional pneumococcal virulence factor that appears to augment intrapulmonary growth and dissemination during the early pathogenesis of S. pneumoniae infection. Inactivation of the PLY gene of pneumococci results in an avirulent mutant 28. Murine monoclonal antibodies to PLY could prolong the survival of mice with pneumococcal pneumonia 29. The inhibition of PLY in murine lung can therefore lead to improvements in the clinical outcome of pneumococcal pneumonia.
This study has also demonstrated that CLR and AZM improve the survival rates of immunocompetent mice suffering from pneumonia caused by high-level MRSP. One report suggests that low-level MRSP can be overcome by increasing the concentrations of macrolides at the site of infection to kill the bacteria 30. However, there are few studies about the clinical outcome of pneumonia caused by erm B-mediated high-level MRSP. Hoffman et al. 25 reported that CLR and AZM did not improve the survival rates. In contrast, in the present immunocompetent mouse model, CLR and AZM were effective even against high-level MRSP. Although the total number of viable pneumococci in murine lungs was similar to that of those treated with control, the life of mice treated with macrolides was prolonged. These prolonged survival rates may be driven by the reduced release of virulence factors of S. pneumoniae, the weak pathogenesis of bacteria and by the immunomodulatory effects of macrolides in immunocompetent mice. The present authors believe that these results may be closer to the clinical effects of macrolides because macrolides are usually prescribed for immunocompetent outpatients.
A limitation of the study is that the authors could not compare the level of PLY to other bacterial virulence factors. However, it is felt that sub-MIC of macrolide may also inhibit other virulence factors, with the exception of house keeping protein. The results suggest that inhibition of PLY production in murine lungs by macrolides plays an important role, at least in part, in the improvement of survival rates from pneumonia caused by high-level MRSP.
In conclusion, this study has demonstrated that sub-minimum inhibitory concentrations of clarithromycin and azithromycin reduce pneumolysin of high-level macrolide-resistant Streptococcus pneumoniae both in vitro and in vivo. These results might be related to the effectiveness of macrolides against pneumonia caused by high-level macrolide-resistant Streptococcus pneumoniae. Further investigation is still needed to clarify the effect of macrolides against macrolide-resistant Streptococcus pneumoniae.
Acknowledgments
The authors are grateful to K. Maesono for excellent technical support.
- Received October 6, 2005.
- Accepted January 10, 2006.
- © ERS Journals Ltd
References










