





Abstract
Human β-defensin (hBD)-3, a 45 amino acid antimicrobial peptide, was originally isolated from human skin. hBD-3 mRNA has also been detected in the airways by RT-PCR. While hBD-3 may be involved in antimicrobial defences within the respiratory tract, the presence of hBD-3 peptide in the respiratory system has not yet been confirmed.
The antimicrobial activity of the synthesised hBD-3 peptide was measured by a radial diffusion assay and a colony count assay. The present authors confirmed the presence of hBD-3 peptide in homogenates of human lung and serum using reverse-phase HPLC coupled with a highly sensitive RIA. The localisation of the hBD-3 peptide was investigated by immunohistochemistry. In addition, the serum concentrations of hBD-3 were measured by RIA.
hBD-3 exhibited a strong antimicrobial activity, which was unaffected by increasing salt concentrations. Immunohistochemically, the current authors observed the expression of hBD-3 in bronchial and bronchiolar epithelial cells. The mean±sd serum concentration of hBD-3 in patients with bacterial pneumonia was 239.4±17.8 pg·mL−1 in the acute phase and, decreased to 159.3±20.1 pg·mL−1 after the completion of therapy.
In conclusion, these findings will help elucidate the role of human β-defensin-3 in host immune responses and identify the pathophysiological significance of this molecule in respiratory infections.
To protect against infections, the respiratory tract possesses a variety of defence mechanisms, including coughing, mucociliary clearance and epithelial cell secretions. The airway surface fluid, which contains bactericidal molecules, such as lysozyme, lactoferrin, secretory phospholipase-A2 and defensins, also exhibits antimicrobial activity.
Defensins, which are strongly cationic, single-chain peptides with molecular masses ranging from 3.5–5 kDa 1, 2, exhibit a broad spectrum of microbicidal activity against gram-positive and gram-negative bacteria, mycobacteria, fungi and certain enveloped viruses 1–3. Defensins are classified into the α- and β-defensin subfamilies based on the pairing of cysteines in three disulphide bridges 1. Six α-defensins and four β-defensins have been previously identified in humans 1. Human neutrophil peptides (HNPs) 1–4 (HNP-1 to HNP-4), localised to the azurophilic granules of neutrophils, are secreted in the early stages of inflammation, as an antimicrobial agent during systemic infectious disease 4, 5. The two other remaining α-defensins, human defensins 5 and 6, are made primarily by Paneth's cells of the small intestine and epithelial cells of the female urogenital tract 1. Human β-defensin (hBD)-1 is constitutively expressed in the epithelia of the urogenital and respiratory tracts 6, while hBD-2 is present in the skin, respiratory tissues and gastrointestinal tract 7, 8. A novel hBD, hBD-4, was recently discovered by genomic sequence analysis mapping to chromosome 8p23, the locus containing a cluster of all known α- and β-defensins 9. Use of a whole Escherichia coli affinity column capable of binding cationic antimicrobial peptides isolated hBD-3 from lesional psoriatic scales from patients with psoriasis 2. Using RT-PCR, hBD-3 mRNA expression was detected in the airway, skin and reproductive organs 2, 3. While hBD-3 may be involved in antimicrobial defences within the respiratory tract, the presence of hBD-3 peptide in the respiratory system has not yet been observed. In the present study, the authors have confirmed the expression of hBD-3 peptide in human lung. Moreover, concentrations of hBD-3 were determined in serum from patients with bacterial pneumonia and these were compared with those of HNP-1, hBD-1 and hBD-2.
MATERIALS AND METHODS
Peptide synthesis
A protected peptide of the hBD-3 peptide was assembled from six segments and a carboxyl-terminal Ne-2-chlorobenzyloxycarbonyl-l-lysine benzyl ester monotosylate using t-butyloxycarbonyl/benzyl chemistry. Each protected segment, corresponding to amino acids 1–8, 9–15, 16–25, 26–31, 32–37 and 38–44, was elongated on resin using N-(9-(hydroxymethyl)-2-fluorenyl)succinamic acid as a linker as described 10. The fully protected peptide was subjected to hydrogen fluoride reduction in the presence of p-cresol. The resulting reduced peptide was purified by reverse-phase (RP)-HPLC. Oxidative folding reaction was performed for 48 h at 4°C in 2 M ammonium acetate buffer (pH 7.8) in the presence of reduced (GSH) and oxidised (GSSG) glutathione at a final ratio of 1:100:10 (reduced hBD-3:GSH:GSSG). A distinct peak eluted at a shorter retention time than that of the fully reduced peptide by RP-HPLC analysis (Zorbax 300SB-C18; Zorbax, Agilent, Palo Alto, CA, USA; at 70°C). This peak was isolated from the reaction mixture by RP-HPLC on an YMC C18 column (YMC, Kyoto, Japan). The crude peptide obtained from this procedure was further purified by RP-HPLC and ion-exchange chromatography (IEX) on CM-Sepharose (Sephadex LH-20; Pharmacia Biotech, Uppsala, Sweden). To obtain the acetate form, the purified peptide was passed through a Muromac column (Muromachi Technos Co. Ltd, Tokyo, Japan) and then a Sephadex LH-20 column (the yield of the oxidised peptide is 55% of that of the reduced peptide). The purity of the resulting synthetic hBD-3 preparation was confirmed by RP-HPLC, IEX-HPLC, capillary zone electrophoresis, amino acid analysis and matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) mass spectrometry (observed m/z was 5156.4). The disulfide connectivity of synthetic hBD-3 was determined to be cysteine (Cys)11–Cys40, Cys18–Cys33 and Cys23–Cys41 by enzymatic digestion with a mixture of trypsin and chymotrypsin at pH 7.8, followed by molecular weight and sequence analyses as described 10. The full length hBD-2 peptide was synthesised by the solid phase technique as previously described 11.
Bactericidal assay
Radial diffusion 12 and colony count 13 assays were used to examine antimicrobial activity. The present authors tested synthetic hBD-2 and hBD-3 antimicrobial activity by radial diffusion assay using E. coli strain HB101 (Toyobo Co. Ltd, Osaka, Japan). Briefly, bacteria were cultured overnight at 37°C in trypticase soy broth (TSB; Nissui Pharmaceutical Co. Ltd, Tokyo, Japan). An aliquot of this culture was transferred to the fresh TSB and incubated for 4 h at 37°C to obtain cells in logarithmic-phase growth. Following the precipitation of bacteria by centrifugation at 800×g for 10 min, samples were washed in PBS and quantified spectrophotometrically by measuring optical density at 620 nm. A culture volume containing 1×106 bacterial colony-forming units (cfu) was then added to 10 mL warm (40–50°C) autoclaved PBS containing 3 g of TSB medium and 1% low electroendosmosis type agarose. After rapid dispersion of bacteria, the bacteria-containing agar was poured into plates to form a uniform layer. Wells measuring 3 mm in diameter were then created in the agar using a gel punch. Diluted peptides or 5 µL control samples were added to each well; samples were then incubated for 18–24 h at 37°C. Antimicrobial activity corresponds to difference in size between the clear zone surrounding the wells containing defensins and those containing control samples.
The bactericidal activities of hBD-2 and hBD-3 were also examined by colony count assay using Staphylococcus aureus (MRSA; NUMR 101, which was isolated from clinical samples at Nagasaki University Hospital (Nagasaki, Japan) 14), E. coli HB101 and Pseudomonas aeruginosa PAO1 (supplied by T. Hayashi, Dept of Microbiology, Miyazaki University, Miyazaki, Japan). Bacteria (S. aureus: 1×107 cfu·mL−1; E. coli and P. aeruginosa: 5×105 cfu·mL−1) were incubated for 2 h at 37°C in 50 µL incubation media containing varying concentrations of hBD-2 or hBD-3. Two incubation media conditions were tested: 1) low salt conditions (Na+ 95 mEq·L−1, Cl− 90 mEq·L−1, K+ 25 mEq·L−1, osmolarity 210 mOsm·kg−1; pH 7.1) and 2) high salt conditions (Na+ 137 mEq·L−1, Cl− 130 mEq·L−1, K+ 4.2 mEq·L−1, osmolarity 270 mOsm·kg−1; pH 7.4). The incubation mixtures were serially diluted, spread on nutrient agar plates and incubated for 18 h at 37°C. The bactericidal activity is expressed as the colony reduction ratio, defined as the ratio of killed bacteria to control bacteria. All assays were performed in three independent tests with duplicates. The statistical significance of the assay was evaluated using a two-tailed t-test; p-values <0.05 were considered significant.
Preparation of antiserum
hBD-3 (2.5 mg) was conjugated to bovine thyroglobulin (15 mg) using 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide hydrochloric acid (400 mg) as described 15. Samples were then dialysed against 2 L of 0.9% sodium chloride five times to remove unconjugated material. The antigenic conjugate solution (0.7–2.3 mL) was used to immunise three New Zealand white rabbits by multiple intra- and subcutaneous injections. After administering booster shots every 2 weeks, the animals were bled 7 days after each injection. The study and all the experimental protocols were approved by the Ethics Review Committee for Animal Experimentation of Miyazaki Medical College (Miyazaki, Japan).
Study population
For experiments of western blotting, immunohistochemistry and chromatographic characterisation, the current authors obtained human normal lung tissues from three patients at surgery, including: 1) a male aged 61 yrs with lung cancer; 2) a male aged 74 yrs with lung cancer; and 3) a female aged 38 yrs with pulmonary mucormycosis. The patient with pulmonary mucormycosis also exhibited insulin-dependent diabetes mellitus, while the other two patents had no complications that could induce an immunosuppressive condition. All subjects smoked.
For RIA experiments, 20 healthy volunteers (10 male, 10 female) and 20 patients (17 male, three female) with bacterial pneumonia who visited the National Sanatorium Miyazakihigashi Hospital (Miyazaki, Japan) were enrolled in the present study. Healthy volunteers (mean±se age 38.5±7.7 yrs) were typically younger than the patients with pneumonia (age 70.0±17.2 yrs). Serum was obtained from patients in the acute phase of infection (before treatment with antibiotics). A total of 12 out of 20 patients also provided serum samples after completing antibiotic therapy. In the present study, the author's defined completion of therapy as the time point of recovery, patient's examinations revealed normal temperature, no respiratory symptoms and normalisation of the data of C-reactive protein levels. The diagnosis of pneumonia was based on clinical, laboratory (including microbiological) and radiological data in accordance with the guidelines from the Infectious Disease Society of America 16. Bacterial pneumonia was confirmed by Gram's staining tests and quantitative sputum cultures when possible. The current authors could detect the pathogens in only six out of 20 patients by cultures, with those six patients testing positive for Streptococcus pneumoniae (three), Haemophilus influenza (one), S. aureus (one), and E. coli (one). Although the causative pathogens of the remaining 14 patients were not identified, the patients recovered following empirical therapy with β-lactam antibiotics, prompting their inclusion in this study. All healthy individuals and patients gave written informed consent.
Immunohistochemistry
Human lung tissues, isolated as described above, were fixed in 3.7% formaldehyde in 10 mM PBS (pH 7.2), dehydrated in a graded ethanol series and embedded in paraffin. Cut sections (3-µm thick) were deparaffinised in xylene, rehydrated in graded ethanol series and then washed in Tris-buffered saline (TBS) containing Tween 20 (DakoCytomation Co. Ltd, Kyoto, Japan). For antigen retrieval, the sections were incubated in 1 µg·mL−1 proteinase K (DakoCytomation) for 30 min at 37°C, and treated with 6% hydrogen peroxidase for 60 min to inactivate endogeneous peroxidases. Nonspecific binding was inhibited by incubation in a protein block (DakoCytomation) for 3 h at 37°C. Preparations were incubated overnight at 4°C with anti-hBD-3 antiserum at final concentration of 1:50,000. Staining was visualised using the Dako CSA system (DakoCytomation) according to the manufacturer's protocol. Control studies utilised normal rabbit serum or anti-hBD-3 antiserum that had been pre-absorbed with 100 pg·mL−1 hBD-3.
Western blot analysis
Human lung tissues, isolated as described above, were heated at 95–100°C for 10 min in a 10-fold volume of water to inactivate intrinsic proteinases. After cooling to 4°C, acetic acid (CH3COOH) and hydrochloric acid were added at final concentrations of 1 M and 20 mM, respectively. Following homogenisation in a polytron for 15 min, the homogenate was centrifuged at 18,500×g for 30 min at 4°C. The resulting supernatant was applied to a Sep-Pak C-18 cartridge (Waters, Milford, MA, USA) pre-equilibrated in 0.5 M CH3COOH. Peptides were eluted in 50% acetonitrile (CH3CN) containing 0.1% trifluoroacetic acid (TFA). The eluate was separated using 12% reducing sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Amersham Pharmacia Biotech, Piscataway, NJ, USA) in 20% methanol, 25 mM Tris-hydrochloric acid, and 0.2 M glycine. Nonspecific binding was blocked by incubating the membranes with 3% skimmed milk in TBS with 0.1% Tween 20 for 2 h at room temperature. Immunoreactive proteins were detected by incubating the membranes with rabbit anti-hBD-3 antibody (1:2,000, respectively) overnight at 4°C. Between each step, the membranes were washed three times for 10 min with TBS/0.1% Tween 20. Subsequently, the membranes were incubated for 1 h with anti-rabbit Immunoglobulin (Ig)G conjugated to horseradish peroxidase (1:3,000), rewashed, and developed using ECLTM reagents (Amersham Pharmacia Biotech) and exposed to film.
Radioimmunoassay procedure
hBD-3 was radioiodinated by the lactoperoxidase method 17. The 125I-labelled peptide was purified by RP-HPLC on a TSK ODS 120A column (Tosoh Co. Ltd, Tokyo, Japan). RIA reaction mixtures were incubated in 150 mM sodium phosphate (pH 7.0) containing 0.25% N-ethylmaleimide-treated bovine serum albumin, 100 mM NaCl, 25 mM EDTA·2Na, 0.05% NaN3, 0.1% Triton X-100 and 3.1% Dextran T-40. Diluted samples or standard peptide solutions (100 µL) were incubated for 24 h in 100 µL of diluted antiserum number 2–7 (final concentration: 1:1,000,000). A solution of the tracer, 16,000–18,000 counts per min (cpm) of 125I-labelled peptide in 100 µL reaction buffer, was incubated with samples for 24 h. Normal rabbit serum and anti-rabbit IgG goat serum were then added for an additional 12 h incubation. Bound and free ligands were separated by centrifugation. All procedures were performed at 4°C. Samples were assayed in duplicate.
Chromatographic characterisation of immunoreactive hBD-3 in lung and serum
Lung
Human lung tissue, isolated and prepared as described above for western blotting, was examined by RP-HPLC on a TSK ODS SIL 120A (Tosoh Co. Ltd) column using a linear gradient of 10–50% CH3CN containing 0.1% TFA at a rate of 1.0 mL·min−1 for 40 min. All fractions were assayed for hBD-3 by RIA.
Serum
Normal human serum (8 mL) diluted two-fold with 0.9% saline was applied to a Sep-Pak C-18 cartridge pre-equilibrated with 0.9% saline. Adsorbed peptides were eluted in 50% CH3CN solution containing 0.1% TFA. The eluate was fractionated by RP-HPLC; all obtained fractions were assayed by RIA.
Quantification of serum concentrations of hBD-3 in normal subjects and patients with pneumonia
Serum samples (1 mL) were applied to a Sep-Pak C-18 cartridge pre-equilibrated with 0.9% saline. Adsorbed peptides were eluted in 50% CH3CN containing 0.1% TFA. Lyophilised eluates were assayed by hBD-3-specific RIA. The serum concentrations of HNP-1, hBD-1 and hBD-2 were also assayed by previously established RIA methods 8, 18, 19. Data are expressed as the mean±sem. Differences between groups were examined using the Mann-Whitney U-test; p-values <0.05 were considered significant.
RESULTS
Bactericidal activity of hBD-3
The present authors examined the bactericidal activity of synthesised hBD-3 by radial diffusion assay. hBD-3 exhibited a dose-dependent bactericidal activity against E. coli, as was seen with hBD-2 (fig. 1⇓). The bactericidal activities of hBD-3 against S. aureus, E. coli and P. aeruginosa were also investigated by colony count assay using two different electrolyte concentrations (fig. 2⇓). The 90% inhibitory concentration (IC90) values for hBD-2 against S. aureus were >125 µg·mL−1 in low and high salt media, respectively; against E. coli were 40 and 60 µg·mL−1 in low and high salt media, respectively; against P. aeruginosa, the IC90 values were 22 µg·mL−1 and >125 µg·mL−1 under low and high salt condition, respectively. For hBD-3, the IC90 against S. aureus were 1.0 and 0.9 µg·mL−1 in low and high salt media, respectively; against E. coli were 13 and 14 µg·mL−1 under low and high salt condition; against P. aeruginosa, the IC90 were 17 µg·mL−1 in both low and high salt media.

Bactericidal activities of human β-defensin (hBD)-2 (▪) and hBD-3 (•) against Escherichia coli strain HB101. Quantitated increases in zone size represent measurement of the zone size at each defensin concentration minus the zone size of the central control well (3 mm). Data are presented as the mean±se.

Bactericidal activities of human β -defensin (hBD)-2 (•) and hBD-3 (○) against a) Staphylococcus aureus (MRSA; clinical isolate) in low electrolyte medium, b) S. aureus in high electrolyte medium, c) Escherichia coli HB101 in low electrolyte medium, d) E. coli in high electrolyte medium, e) Pseudomonas aeruginosa PAO1 in low electrolyte medium, and f) P. aeruginosa in high electrolyte medium. Bactericidal activity is expressed as the percentage of bacteria killed, obtained from the ratio of surviving bacteria in treated versus control sample. Data are presented as mean±se. *: p<0.01; **: p<0.001 versus hBD-2.
Identification of hBD-3 peptide in the lung and serum
In the three studied lung samples, hBD-3 immunoreactive cells were diffusely observed in the bronchial and bronchiolar epithelium (fig. 3a⇓ and b). Airway epithelial cells showed strong and granular cytoplasmatic immunostaining. hBD-3 immunoreactivity could not be detected in either the mucus layers or alveolar epithelial cells. Tissue immunoreactivity was abrogated by pre-absorption of the antiserum with 100 pg·mL−1 hBD-3 peptide (fig. 3c⇓).

a) Bronchiolar epithelial cells stained positively in the immunohistochemical reaction and at higher magnification (b and c). d) No immunoreactivity in tissues when following pre-adsorption of antiserum with 100 pg·mL−1 human β-defensin-3 peptide. Scale bar = 500 µm (a), 100 µm (b) and 50 µm (c, d).
By RIA, antiserum number 2–7 recognised hBD-3 with a high affinity at final dilutions of 1:1,000,000 (35% binding). Half-maximal inhibition occurred at 60 pg·tube−1. Peptide remained detectable at the low levels of 6.0 pg·tube−1. At 50% binding, the respective intra- and inter-assay coefficients of variation were 3.6 and 4.6%. This antiserum did not exhibit any cross-reactivity against HNP-1, hBD-1, or hBD-2. Immunoreactive hBD-3 could also be identified in human lung and serum by the combination of RP-HPLC with RIA (fig. 4⇓). In both samples, hBD-3-immunoreactive peaks eluted at the same position as the synthetic hBD-3 peptide. Western blotting experiments also demonstrated the existence of hBD-3 peptide in human lung tissue (fig. 5⇓).

Representative reverse phase (RP)-HPLC profiles of human β-defensin (hBD)-3 immunoreactivity (ir-hBD-3; •). Samples were obtained from a) 300 mg human lung tissue and b) 8.0 mL of human serum. Fraction volumes of 0.5 mL were obtained by RP-HPLC using a TSK ODS SIL 120A (4.6 Å×150 mm) column and a linear gradient of 10–60% acetonitrile (CH3CN; ---) containing 0.1% trifluoroacetic acid at a rate of 1.0 mL·min−1 for 40 min. Arrows indicate the elution position of synthetic hBD-3.

Western blotting analysis of human β-defensin (hBD)-3 peptide expression in human lung tissue. Lane 1: 10 ng recombinant hBD-3 peptide. Lane 2: 10 mg human lung tissue extract (this sample corresponds to the sample in figure 4a⇑). MW: molecular weight.
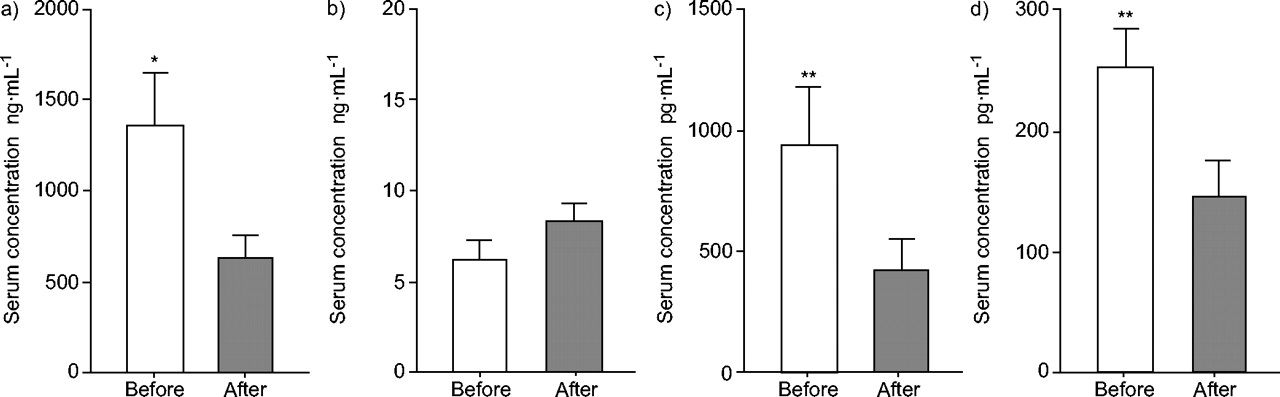
The mean±se serum concentrations of hBD-3 in normal individuals averaged 142.9±10.6 pg·mL−1, with no observable differences between the sexes (table 1⇓). The serum concentrations of hBD-3 in 20 patients with an acute phase of bacterial pneumonia were higher than those seen in normal subjects (246.8±25.4 pg·mL−1; p<0.01). The alterations in the serum concentrations of the four defensin family peptides HNP-1, hBD-1, hBD-2 and hBD-3 are detailed in figure 6⇓ for 12 patients with bacterial pneumonia during antimicrobial chemotherapy. While the serum concentrations of HNP-1, hBD-2 and hBD-3 increased during the acute phase of pneumonia, those of hBD-1 did not change throughout the clinical course of the infection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The effect of chemotherapy on the serum concentrations of four defensins in 12 patients with bacterial pneumonia: a) human neutrophil peptide-1, b) human β-defensin (hBD)-1, c) hBD-2 and d) hBD-3. Data are presented as mean±se. *: p<0.05; **: p<0.01 versus after completion of the therapy.
Serum concentrations of humanβ-defensin-3 in normal subjects and patients with bacterial pneumonia
DISCUSSION
Defensins comprise a family of genes whose products have complementary functions. However, the systemic distribution and antimicrobial activities of these peptides differ significantly between defensin molecules 1. hBD-3 is a novel defensin peptide originally isolated from the scales of a patient with psoriasis 2. While hBD-3 mRNA was detected in human trachea by RT-PCR 2, 3, the presence of hBD-3 peptide has not previously been demonstrated in the human respiratory system. Synthetic hBD-3 exhibits a broad-spectrum antimicrobial activity against both gram-positive and gram-negative bacteria 2, 3, while hBD-1 and hBD-2 are more potent against gram-negative bacteria 1, 2. Recently, host protease cathepsins were shown to degrade defensins; thus, overexpression of cathepsins may produce an environment favouring bacterial infection and colonisation during chronic pulmonary infectious diseases 20. All defensin molecules are strongly cationic, facilitating their interactions with bacteria and allowing the formation of channels within the negatively charged cytoplasmic membranes 21. The antimicrobial activities of desalted airway surface fluids (ASF) obtained from both cystic fibrosis and normal xenografts were significantly higher than crude ASF obtained from their xenografts 22. These results suggest that high sodium chloride concentrations may inactivate defensin antimicrobial activity by weakening the electrostatic interactions between defensins and the cytoplasmic membrane. It is plausible that, under these conditions, a greater number of the charged residues in defensins can reduce the salt effect that interferes with the binding of defensins and the negatively charged lipid components of membranes 23. The present authors ascertained the antimicrobial activities of synthetic hBD-2 and hBD-3 under low and high electrolyte conditions. hBD-3 contains more positively charged amino acid residues than other β-defensins 23, which may account for its similar antimicrobial activity under different sodium concentrations.
The hBD-3 peptide could be detected within the bronchial and bronchiolar regions, but not in the alveolar epithelium. In contrast, hBD-2 is expressed by type-2 alveolar epithelial cells 24. This differential distribution of these two defensins may contribute to their unique functions in innate host defences to invading microbes within the respiratory system.
Serum concentrations of hBD-3 in patients with bacterial pneumonia increased during the acute phase, normalising after treatment with antibiotics. Although the mechanism of biosynthesis and release of endogenous hBD-3 peptide are not well understood, the presence of hBD-3 peptide in the bronchial and bronchiolar epithelia and the change in serum hBD-3 concentrations during antibiotic therapy for bacterial pneumonia suggest that serum hBD-3 might be secreted by bronchial and bronchiolar cells in bacterial pneumonia.
The current authors previously determined the concentrations of three additional defensins (HNP-1, hBD-1 and hBD-2) in the serum and bronchoalveolar lavage (BAL) fluid during pulmonary disease states 9, 24–29. The concentrations of both HNP-1 and hBD-2 increased in the serum and BAL fluid during the acute phase of respiratory infections, returning to basal levels after the completion of therapy 25–27. Serum HNP-1 and hBD-2 concentrations also increased in noninfectious pulmonary diseases, including idiopathic pulmonary fibrosis 27 and diffuse panbronchiolitis 24, 29. The serum concentrations of hBD-3 also increased in the acute phase of bacterial pneumonia, normalising in the recovery phase. These results suggest that serum hBD-3 concentration mirrors the activity of bacterial pneumonia. hBD-3 gene expression in the respiratory epithelium was also markedly upregulated by interferon-γ stimulation and P. aeruginosa infection 2, 3, indicating that pathogenic microorganisms and inflammatory cytokines stimulate hBD-3 synthesis and release during respiratory infection.
The presence of human β-defensin-3 in the airways, along with its strong bactericidal activity, suggests that human β-defensin-3 functions in humans in innate mucosal defence within the respiratory tract. Establishment of a quantitative assay for human β-defensin-3 activity will provide a better understanding of the physiological role and pathophysiological significance of this molecule in respiratory infectious diseases.
Acknowledgments
The authors would like to thank T. Minematsu (Nichinan Aisenkai Hospital, Nichinan, Japan) for invaluable advice in the experiments of bactericidal activities. The authors would also like to thank N. Chino (Peptide Institute, Osaka, Japan) for his excellent technical advice in peptide synthesis and, S. Yanagi and S. Tajiri (University of Miyazaki, Miyazaki, Japan) for their excellent technical assistance.
- Received September 10, 2004.
- Accepted October 3, 2005.
- © ERS Journals Ltd
References









