





Abstract
T-helper cell type (Th)2 lymphocytes play an important role in the initiation, progression and persistence of allergic diseases, including asthma. However, little is known about immunoregulatory mechanisms that determine susceptibility to, severity of, or persistence of asthma. The concept of a disturbed Th1/Th2 balance, although having furthered the present understanding of immunoregulation in asthma, has recently been named a “procrustean paradigm”, because of its failure to adequately explain many (pre)clinical observations.
In recent years, the general knowledge regarding the regulation of infectious, autoimmune diseases, asthma and allergen immunotherapy by T-regulatory (Treg) cells, has rapidly increased. Many different Treg subsets have been described, including CD8+ Treg cells, natural killer (NK) cells and several different CD4+ Treg cell subsets.
In this review, the authors will focus on two major and well-described CD4+ Treg cell subsets. These consist of naturally occurring CD25+ Treg cells and adaptive Treg cells that are postulated to prevent immune responses against self-antigens and adaptive immune responses, respectively.
The adaptive T-regulatory cells are further subdivided into T-regulatory cells type 1 and T-helper cell type 3 that mediate suppression exclusively via the cytokines interleukin-10 and transforming growth factor-β, respectively.
Foreign antigens, including allergens or pathogens, that enter the body are taken up by so-called antigen-presenting cells (APC), which process the antigens and present peptides, thereof, in the context of major histocompatibility complex class (MHC) II molecules on their cell surface. A fraction of the CD4+ T-lymphocytes, also called T-helper (Th) lymphocytes, are activated by cognate interaction of the T-cell receptor (TCR) with the peptide-MHC II complex on APC when there are sufficient co-stimulatory signals, i.e. CD28:B7 1. Once activated, the Th-cells orchestrate adaptive antigen-specific cell-mediated and humoral immune responses. Incited by the role of CD4+ T-lymphocytes in the production of immunoglobulin (Ig) E antibody by B-lymphocytes, subsequent studies in the late 1980s demonstrated that they are also involved in the pathophysiology of allergic asthma 2, 3.
The discovery of the mutual inhibitory effects of Th1 and Th2 cells in mice 4, prompted the postulate that an imbalance between these two arms of the immune response would underlie Th1-mediated autoimmune diseases as well as Th2-mediated allergic diseases, including asthma 5, 6. However, the balance between Th1 and Th2 cells appeared insufficient to explain many experimental observations 7–9. First, Th1 cells do not always appear beneficial in mouse models of allergic asthma, and were found to contribute to or to exacerbate, disease manifestations 10–14. This is in accordance with the observed appearance of Th1 cells or the cytokine interferon (IFN)-γ, not only in chronic atopic dermatitis and asthma, but also during allergic sensitisation 15–18. Secondly, Th2-skewed parasitic helminth infections did not appear to be associated with increased manifestations of allergy and asthma but, on the contrary, appeared to protect against these diseases 19. Finally, epidemiological data from the 1960s onwards have shown a parallel rise in the prevalence of Th2-mediated allergic diseases, including asthma and Th1-mediated (auto-) immune diseases, such as type 1 diabetes, multiple sclerosis, and Crohn's disease 20, 21. Whereas these data do not exclude a mutual inhibitory effect of Th1 and Th2 responses, a more powerful control mechanism is likely to be at play. At present, naturally occurring and adaptive T-regulatory cells (Treg) cells (nTreg and aTreg, respectively) are taking the centre stage as the crucial immunoregulatory cells that are capable of suppressing Th1- and Th2-mediated adaptive immune responses in a cell contact-dependant fashion directly or by acting on APCs. They also appear to offer exciting new views of the initiation and progression of asthma and other allergic diseases. In figure 1⇓, the major characteristics of CD4+ Treg cells are depicted.

Major characteristics of subsets of CD4+ T-regulatory (Treg) cells based on cell-surface markers, immunosuppressive cytokine secretion and suppressive action. nTreg: natural Treg; aTreg: adaptive Treg; Th; T-helper cell; Tr1: T-regulatory cell type 1; APC: antigen-presenting cells; TGF: transforming growth factor; IL: interleukin.
NATURAL T-REGULATORY CELLS
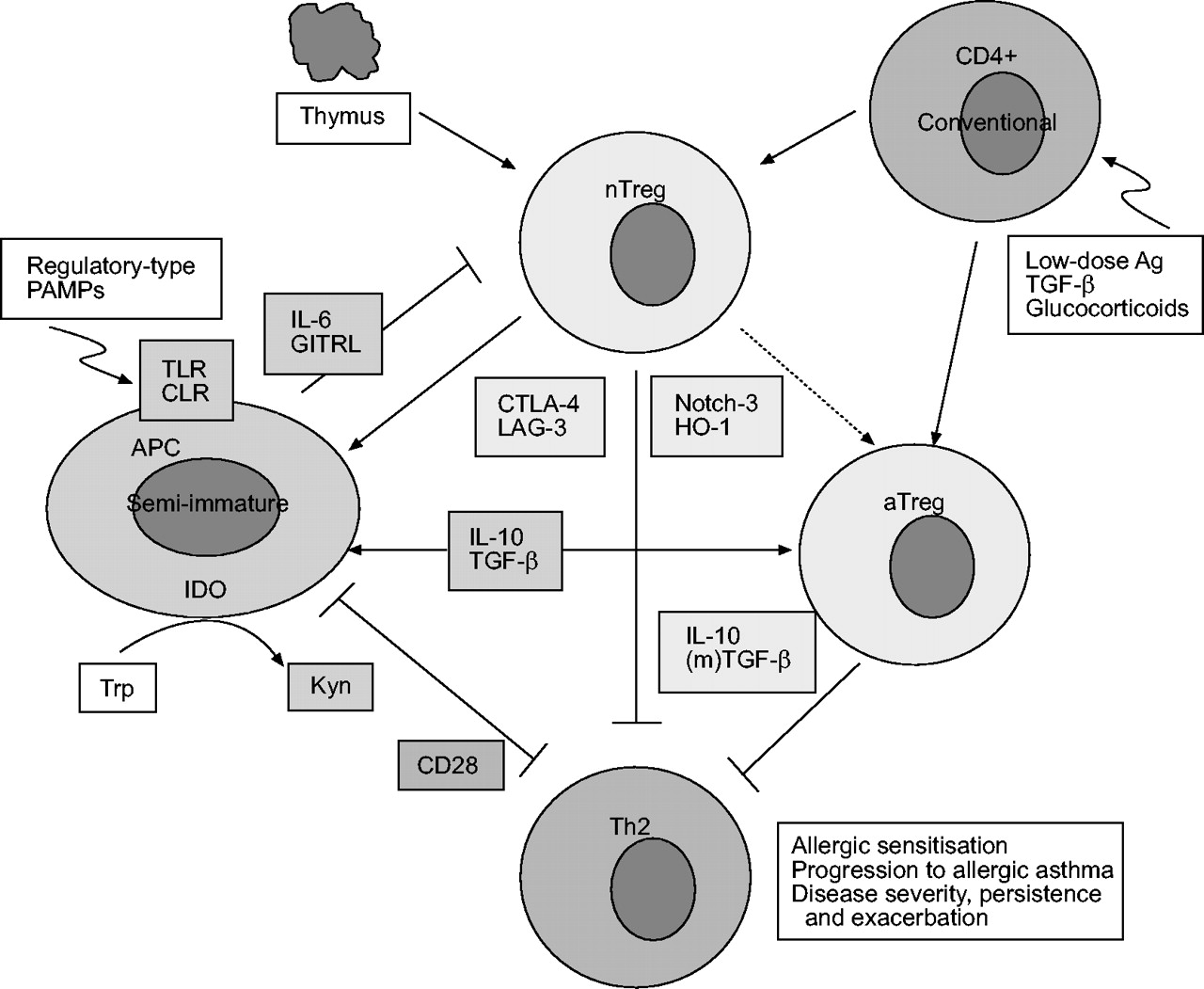
CD4+CD25+ nTreg cells are generated in the thymus and reside in the blood and other peripheral lymphoid tissues at a frequency of 5–10% of all CD4+ cells and in the bone-marrow ≥20%, both in mice and in humans 22–25. Passive transfer of nTreg cells has been shown to suppress autoimmune diabetes, inflammatory bowel disease and transplant rejection in rodents. nTreg cells express a normal α/β TCR repertoire and can be activated by anti-CD3 24, 26, 27. It has been shown that nTreg cells have a high TCR affinity for self-peptides that might be encountered in the periphery, which may explain the constitutive expression of the activation marker, CD25 27–29. However, the target autoantigens recognised by nTreg cells are, at present, unknown. Although still controversial, accumulating data reports that nTreg cells can also be generated in the periphery from conventional CD4+ T-cells upon exposure to pathogen-derived antigens 30, low-dose antigenic peptide via prolonged subcutaneous infusion 31, the cytokine transforming growth factor (TGF)-β 32–34, or glucocorticoids 35 (fig. 2⇓).

Schematic representation of the suppressive mechanisms of T-regulatory (Treg) cells subsets and their interactions with each other and antigen-presenting cells (APCs), in particular on dendritic cells. Natural Treg cells (nTreg) are predominantly generated in the thymus, but can also develop from conventional CD4+ cells by specific conditions or signals. nTreg cells suppression of effector T-helper cell type 2 (Th2) cell responses involves the cell surface molecules CTLA-4, Notch-3, and LAG-3, and the intracellular enzyme, haeme oxygenase (HO)-1. CTLA-4 and CD28 deliver opposite signals to APCs, thereby regulating indoleamine 2,3-dioxygenase (IDO) activity and its anti-proliferative effects on effector T-cells. The suppressive effects of nTreg cells are abrogated by GITRL and interleukin (IL)-6 that can be expressed by APCs. Immature or semi-mature APCs producing IL-10 or transforming growth factor (TGF)-β generate T-regulatory cell type 1 (Tr1)- and Th3-type aTreg cells, respectively. Production of these cytokines by APCs can be induced by so-called regulatory-type pathogen-associated molecular patterns (PAMPs). Data suggest that nTreg cells also mediate the development of aTreg (Tr1, Th3) cells. IL-10 and (membrane-associated) TGF-β derived from aTreg cells and potentially nTreg cells suppress effector Th2 cell responses directly or by acting on APCs. Trp: tryptophan; Kyn: kynurenin; TLR: Toll-like receptor; CLR: C-type lectin receptor; Ag: antigen. See text for further details regarding these interactions
Studies performed so far with human CD4+CD25+ nTreg cells, without exception, confirmed data obtained in mouse studies 2, 23, 27, 36. Human CD4+CD25+ nTreg cells show poor proliferative responses upon stimulation with anti-CD3, even in the presence of interleukin (IL)-2, suppress proliferative responses of conventional CD4+CD25- T-cells, and produce low levels of the immunosuppressive cytokines, IL-10 and TGF-β, while the suppression, at least in vitro, cannot be overcome by the addition of neutralising antibodies to these cytokines or their respective cell-surface receptors. However, a role for IL-10 or (membrane-associated) TGF-β in vivo, cannot be completely excluded, as indicated by mouse experiments. Their involvement may be indirect, since nTreg cells were found to aid the generation of aTreg cells that mediate suppression by these cytokines (fig. 2⇑) 39, 40.
In vitro, nTreg cells appear to suppress adaptive T-cell responses exclusively by T-cell:T-cell/APC contact. The molecular basis for this suppression is unknown at present. The cell-surface molecules, CTLA-4 41, 42, Notch-3 43, and LAG-3 44, as well as the intracellular enzyme, haeme oxygenase-1 (HO-1) that generates the immunosuppressant, carbon monoxide 45, have all been shown to play a role. Interestingly, CTLA-4 ligation of B7-1 or B7-2 on dendritic cells (DCs) was found to result in the induction of the enzyme indoleamine 2,3-dioxygenase (IDO) in DCs 46, 47. IDO is the rate-limiting enzyme in the conversion of tryptophan to kynurenin and other down-stream metabolites. It may also mediate potent immunosuppression by T-cell starvation, due to the depletion of tryptophan or antiproliferative and pro-apoptotic effects of its down-stream metabolites 48, 49. In contrast to CTLA-4 ligation of DC-expressed B7 molecules, their ligation by CD28 stimulates T-cells and interestingly, induces IL-6 production that prevents upregulation of IDO 50, 51. In summary, nTreg cells may suppress effector T-cells by different pathways, as schematically depicted in figure 2⇑.
A major issue in the study of Treg subsets is the lack of discriminative cell-surface markers (fig. 1⇑). CD25 expression has been linked to the subpopulation of nTreg cells. However, CD25 is a general activation marker of T-cells and CD4+CD25+ cells may, therefore, contain recently activated conventional T-cells. Thus, in a “resting” situation when there is no ongoing immune response, CD25 may be an appropriate marker for nTreg cells, but during active disease it is not sufficiently distinctive for this purpose. However, as nTreg cells appear to express very high levels of CD25, this could be useful to differentiate between activated T-cells and nTreg cells 52. nTreg cells are further characterised by a set of cell-surface markers, none of which are able to completely discriminate between nTreg cells and activated conventional T-cells. These markers include CTLA-4, CD45RBlow/CD45RO (mouse/human), CD38, CD62L, glucocorticoid-induced tumour necrosis factor (TNF) receptor (GITR, also known as TNFRSF18) and neuropilin-1, of which the latter is the only one that is down-regulated on activated conventional T-cells (fig. 1⇑) 24, 53. Furthermore, subpopulations of nTreg cells have been described based on the expression of integrins. Among these, αEβ7 (CD103) appeared to define a subpopulation of nTreg cells that is highly potent in suppression. This population is considered to be restricted to mucosal surfaces, since it binds via αEβ7 to E-cadherin expressed by mucosal epithelial cells 54. Interestingly, these αEβ7+ nTreg cells express high levels of adhesion molecules and chemokine receptors that suggest a role at inflammatory sites 55. In addition, Stassen et al 37 have described α4β7+ and α4β1+ nTreg cells populations that induce T-regulatory cells type 1 (Tr1) cells or Th3, respectively. It can be concluded that nTreg cells can exert powerful immunosuppressive effects and current knowledge is rapidly progressing, although many details still need to be unravelled and observations reproduced before the bigger picture will emerge.
FOXP3 AND NATURAL T-REGULATORY CELLS
Expression of specific transcription factors have been shown to be crucial to the differentiation of Th cell subsets from naïve, unpolarised T-cells. Notably, T-bet and GATA-3 are crucial to Th1 and Th2 cytokine expression, respectively 56. Recently, the transcription factor, Foxp3, was shown to be selectively expressed by nTreg cells both in mice and humans 57–60. The Foxp3 mRNA level in resting nTreg cells is 100-fold higher than in resting and activated conventional CD4+ T-cells. Therefore, Foxp3 represents a more specific marker for nTreg cells than cell-surface molecules currently used, such as CD25, CTLA-4, and GITR. However, a major disadvantage for nTreg characterisation is that Foxp3 expression is currently determined by quantitative PCR and not by flow cytometry.
Interestingly, ectopic expression of Foxp3 by naïve T-cells appeared essential to the generation of Treg cells as demonstrated by cell-surface markers and cell–cell contact dependent, but IL-10- and TGF-β-independent, antiproliferative effects on T-cells 58. Vice versa, Foxp3 knockout or mutant (“scurfy”) mice lacked CD4+CD25+ nTreg cells and developed a lethal autoimmune syndrome 59, 60. Moreover, TGF-β can induce Foxp3 expression in naïve T-cells leading to nTreg cell generation in mice 32, 33. Data demonstrates that Foxp3 is a master regulatory gene for the development and function of nTreg cells and confirms its role in the suppression of autoimmune diseases. The crucial role of the human homologue FOXP3 in nTreg cells has now also been firmly demonstrated in humans by Yagi et al. 61. They demonstrated that the FOXP3 gene and its protein product were preferentially expressed in peripheral CD4+CD25+ nTreg cells of normal individuals. Furthermore, naïve conventional CD4+CD25- T cells could be converted into nTreg cells by ex vivo retroviral gene transfer of FOXP3, as confirmed by cell-surface markers and cell–cell contact-dependent antiproliferative effects on conventional T-cells. The fact that FOXP3 is essential to nTreg cell function may open novel therapeutic strategies aimed at the induction of FOXP3 to convert conventional CD4+ T-cells into nTreg cells with the ultimate goal to reverse aberrant Th2-mediated allergic asthmatic responses.
ADAPTIVE T-REGULATORY CELLS
The aTreg cells are generated in the periphery from naïve T-cells after encountering antigens presented by tolerogenic DCs, which are functionally distinct from immunogenic DCs that promote Th1 or Th2 cell generation 62. As already indicated, aTreg cells are subdivided into Tr1 and Th3 that mediate suppression via the cytokines, IL-10 and TGF-β, respectively (fig. 1⇑). The suppressive action of these cytokines is predominantly directed at inhibiting the expression of MHC II and costimulatory molecules by APCs. In addition, direct effects at the T-cell level have been described. TGF-β inhibits proliferation and cytokine secretion by resting, but not activated, T-cells 63 and IL-10 may sensitise activated T-cells to the suppressive action of TGF-β by enhancing TGF-β receptor type II expression on these cells 63. TGF-β has been shown to induce the expression of immunosuppressive IL-10 by T-cells 64. Thus, IL-10 and TGF-β may have synergistic, suppressive effects on T-cell proliferation and immunostimulatory cytokine secretion.
The aTreg subsets variably express CD25, depending on their level of activation and cannot be distinguished solely by the expression of IL-10 or TGF-β, because it is not an exclusive property of these subsets. In contrast to Th1 and Th2 cells, Tr1 cells can also be characterised by absent or low IL-2 and IL-4 production, although they may express IL-5, IFN-γ and TGF-β. Th3 cells may, in addition to TGF-β, also express variable amounts of IL-4 and IL-10. Hence, demonstration of the involvement of Tr1 or Th3 cells in adaptive immune responses requires determination of a discriminative cytokine profile.
ROLE OF DENDRITIC CELLS IN GENERATING ADAPTIVE T-REGULATORY CELLS
DCs play a crucial role in the generation of all adaptive T-cell subsets, including aTreg cells, which has been reviewed in detail elsewhere 65, 66. In short, immature DCs in peripheral tissues acquire pathogens and or antigens from their vicinity through phagocytosis, endocytosis and pinocytosis, and mature upon recognition of so-called “danger signals”. These signals are derived either directly from encountered pathogens/antigens or from endogenous mediators that are present or generated in the local microenvironment 62, 65. Microbial pathogens contain ligands (pathogen-associated molecular patterns: PAMPs) that are recognised through so-called “pattern recognition receptors” (PRRs) of which the family of Toll-like receptors (TLRs) and C-type lectin receptors (CLRs) are prime examples 67–70. Bacteria harbour an abundance of PAMPs ranging from cell wall components (lipopolysaccharides, lipoproteins, peptidoglycans, lipoarabinomannans) to DNA containing unmethylated CpG motifs 69. Recognition of the PAMPs by TLRs causes signalling through a conserved pathway leading to the activation of nuclear factor (NF)-κB, a family of transcription factors implicated in many inflammatory responses and crucial to the maturation of DCs 69, 71. The maturation of DCs is characterised by high levels of expression of MHC-II and costimulatory molecules such as, B7-1, B7-2, and CD40 and by pro-inflammatory cytokine secretion. TLRs are not only involved in DC maturation, but also in the abrogation of Treg cell-mediated suppression. Thus, the combined action may enable development of full-blown immune responses 72. However, TLRs are also instrumental to the induction of tolerogenic DCs, indicating that the codes instructing development of immunogenic or tolerogenic DCs are very complex.
Tolerogenic DCs generally are semi-mature cells with increased expression of MHC II and B7-2, but low levels of expression of CD40 and absent production of the pro-inflammatory cytokines, IL-6 and TNF-α 73. Repetitive stimulation of T-cells with immature, antigen-presenting DC has been shown to generate IL-10 producing Tr1 cells 74–76. Likewise, inhibition of DC maturation by inhibiting NF-κB activation with the biologically active form of vitamin D3 (1,25(OH)2VitD3) or glucocorticoids, induces Tr1-like cells in vitro and tolerance in mouse models of transplantation and autoimmune diseases in vivo 77–79. However, Tr1-like cells can also be induced in vitro in the absence of DCs by activating naïve CD4+ T-cells in the presence of immunosuppressive drugs e.g. 1,25(OH)2VitD3 or glucocorticoids 80.
The mechanisms by which so-called tolerogenic DCs induce the development of aTreg cells is not completely understood. The induction of Tr1 or Th3 cells is related to the production of IL-10 or TGF-β by (semi/immature) DCs, respectively 65, 73. Many pathogens, including viruses, parasites, fungi and bacteria, and isolated, so-called regulatory-type PAMPs have been shown to induce the production of either one or both of these cytokines by DCs, and to facilitate induction of aTreg cells in this way (fig. 2⇑) 30, 65, 70. The local microenvironment also appears to influence the development of tolerogenic DCs, and the subsequent generation of aTreg cells as shown by two forms of tolerance induced via the mucosal route 81. Tolerance by intranasal application of ovalbumin has been associated with the appearance of DCs in the lung that could generate Tr1 cells ex vivo. It appeared to be mediated by DC-derived IL-10 and ICOSL: ICOS interaction 82, 83. Tolerance by intragastric ovalbumin administration, so-called “oral tolerance”, has been associated with TGF-β-producing DCs that generate Th3 cells 81. A role for commensal bacteria, present at mucosal surfaces in particular the gut, in the induction of these allergen-specific aTreg responses cannot be excluded 84–86. One of the molecular mechanisms may be the inhibition of NF-κB activation in epithelial cells by commensal bacteria, leading to an anti-inflammatory and tolerogenic microenvironment 87, 88. Whether similar processes occur at the airway mucosa is unknown at present. The instruction of DCs by microbial ligands leading to immunogenic or tolerogenic responses may be an important link to the hygiene hypothesis of allergic diseases, including asthma. Moreover, this knowledge may be of use in the development of novel therapeutics that target DCs 89.
T-REGULATORY CELLS AND ASTHMA
Theoretically Treg cells may interfere with the development of allergic diseases and asthma at different stages, such as allergic sensitisation, progression to allergic inflammation, airway remodelling and airway hyperresponsiveness (AHR), and persistence of disease manifestations. However, at present little is known about the role of Treg cells in allergy and allergic asthma.
Sensitisation to environmental allergens is known to occur typically in early childhood or even before birth, but subsequent progression to persistent atopic asthma, which may not manifest itself for several years, is restricted to only a subset of atopics 90. This has led to the hypothesis that the induction of a down-regulatory Th1 response, supposedly by microbial stimulation, during the first years of life, prevents allergic sensitisation and progression to atopic asthma 91–93. Irrespective of this hypothesis, Treg cells may also be instrumental in preventing allergic sensitisation and progression to asthma. The nTreg cells are present in cord blood and probably play an immunoregulatory role in intrauterine life 94. Recently, Thornton et al. 95 described how cultures of cord blood T-cells, in the presence of allergens, led to a relative increase in the number of CD4+CD25+ Treg cells. The increase did not appear to be due to proliferation of these nTreg cells, but rather to apoptosis of the proliferating conventional T-cells. Interestingly, these nTreg cells acquired suppressive properties during culture with an allergen. The significance of the in vitro observation for understanding allergic sensitisation and progression to asthma is unclear. More research into the mechanisms by which nTreg cells promote the development of tolerance to harmless environmental allergens during intrauterine and neonatal life is needed. It may lead to novel strategies for primary prevention of allergic diseases, including asthma.
Two recent studies have examined the role of CD4+CD25+ T-cells in adult allergic individuals. Bellinghausen et al. 96 found similar activities of peripheral blood nTreg cells from grass or birch pollen allergic rhinitis patients and healthy controls. They proliferated poorly, produced low levels of cytokine and inhibited proliferation and production of Th1 and Th2 cytokines, but not IL-10, by conventional CD4+CD25- T-cells. Moreover, the inhibition was not reversible by antibodies to IL-10, TGF-β or CTLA-4, but could be reversed by IL-2, which is in agreement with nTreg cell function. Since no differences were observed between healthy and atopic donors, they concluded that nTreg cells are present and functional in most atopic patients. In contrast herewith, Ling et al. 97 showed that nTreg cells from atopic donors suppressed proliferation and IL-5 production of autologous CD4+CD25- T-cells significantly less than nTreg cells from healthy donors, in particular during the pollen season. The latter study also verified the true nTreg cell nature of the isolated CD4+CD25+ cells by measurement of FOXP3 mRNA levels. Since both studies used allergen in the presence of APCs for T-cell stimulation, allergen-specificity of the nTreg cells was demonstrated, so confirming that nTreg cells can be generated in the periphery upon exposure to foreign antigens or pathogens. A likely explanation for the contradictory findings is the time point at which the blood samples were taken. In the study of Bellinghausen et al. 96 this may have been out of the pollen season, when, as shown by Ling et al. 97, nTreg cell activity of allergic patients was substantially lower than during the pollen season. Furthermore, Bellinghausen et al. 96 did not analyse whether the CD4+CD25+ T-cells expressed higher levels of FOXP3 mRNA compared with conventional CD4+CD25- T-cells, thereby not verifying the purity of the T-cell populations used. Although further studies are needed, observations suggest that there may be an inverse correlation between nTreg cell activity and clinical manifestations of allergic disease, as previously observed for autoimmune diseases 98. If so, it needs to be determined whether clinical disease manifestations are the cause or the consequence of decreased nTreg activity.
An interesting observation regarding the role of nTreg cells in the manifestation of allergy comes from Karlsson et al. 99 in children with cow's milk allergy. Children who outgrew their allergy had higher numbers of CD4+CD25+ cells in their blood than those who maintained clinically active allergy. The increased numbers coincided with decreased in vitro T-cell proliferative responses to the food-allergen, bovine β-lacto globulin, but not with proliferative responses to polyclonal stimulation, again indicating the presence of allergen-specific nTreg cells. These had marked suppressive activity, since depletion of CD25+ cells led to a striking (five-fold) increase in the T-cell proliferative response to the food allergen. The suppression by the CD4+CD25+ T-cells was partially dependent on cell contact and not due to increased IL-10 or TGF-β production, supporting the conclusion that these cells were indeed nTreg cells. However, involvement of the immunosuppressive cytokines cannot be completely excluded since no blocking studies were carried out. Studies so far indicate the potential role of nTreg cells in the spontaneous reversal of asthma and other allergic diseases and further studies may shed new light on this intriguing phenomenon.
While aTreg cells are clearly implicated in allergen immunotherapy (referred to later), their putative role in the prevention and severity of allergic asthma is largely unknown at present. The role of IL-10 producing T-cells in the severity of allergic asthma in adults has been studied by Matsumoto et al. 100 by comparing mild, severe-stable and severe-unstable asthmatics. The frequency of IL-10 producing CD4+ T-cells as determined by flow cytometry after anti-CD3/anti-CD28 stimulation was lower in the severe unstable group as compared with the mild or the severe-stable group and could not be attributed to the use of inhaled glucocorticoids. Although these data need confirmation, there is an indication that the number of IL-10 producing cells, potentially Tr1 cells, may be related to the prevention of asthma exacerbations. Akdis et al. 101 analysed the frequency of allergen-activated T-cells according to their cytokine secretion profile in healthy and pollen or house-dust mite (HDM) allergic individuals. They observed a delicate balance in the frequency of Tr1 cells and Th2 cells. The Tr1 cells represented the dominant subset in healthy individuals, whereas high frequencies of IL-4 secreting T-cells were present in allergic individuals. Interestingly, blocking of the Tr1 activity enhanced allergen-induced Th2 cell activation in vitro. Furthermore, they demonstrated that the suppressive action of the Tr1 cells in vitro was mediated by the joint action of IL-10, TGF-β, CTLA-4 and PD-1. Data suggest that Tr1 cells represent a Treg cell subset involved in preventing the development of allergic diseases.
Despite the limited number of studies, it can be concluded that nTreg and aTreg cells are important in the prevention of allergic sensitisation as well as progression to established allergic diseases, including asthma. In established disease, they function to reduce severity and to prevent exacerbations. Although these studies need to be expanded and confirmed by others, Treg cells may offer new possibilities for the monitoring of diseases and the development of novel therapeutic strategies.
GLUCOCORTICOIDS AND T-REGULATORY CELLS
Glucocorticoids are widely used in the treatment of allergic asthma for their potent anti-inflammatory effect. It has recently been shown that pre-incubation of CD4+CD25+ nTreg cells with glucocorticoids, potentiated their antiproliferative effects on allergen-stimulated CD4+CD25- T-cells from atopic as well as nonatopic donors 102. Interestingly, the glucocorticoid-induced potentiation of suppressive activity was paralleled by enhanced IL-10 production and anti-IL-10 could reverse the glucocorticoid-dependent, but not the baseline, suppression. Thus, nTreg cell-derived IL-10 mediates the glucocorticoid-induced suppression. If operative in vivo, it may represent an additional mechanism by which glucocorticoids inhibit inflammation. Glucocorticoids can also influence Tr1/nTreg cell development in vivo, as shown by Karagiannidis et al. 35. They demonstrated how inhalation or systemic glucocorticoid treatment of asthma patients increased the levels of FOXP3 as well as IL-10 mRNA in peripheral blood CD4+ T-cells. Moreover, glucocorticoids increased these mRNA levels in short-term cultured CD4+ T-cells. Altogether, the data demonstrates that glucocorticoid treatment potentiates the immunosuppressive effects of nTreg and/or Tr1 cells, probably by increasing IL-10 production. The Treg population generated by glucocorticoid treatment displayed characteristics both of nTreg cells (FOXP3) and Tr1 cells (IL-10), but whether it is a pure population of CD4+CD25+ nTreg cells or a mixture of nTreg cells and Tr1 cells cannot be concluded from these studies. Likewise, it is unknown whether the induction of Tr1 cells is dependent on the presence of nTreg cells. In literature, there is still controversy as to whether Tr1 cells can express FOXP3 24, 61. Furthermore, in contrast to the mouse, activation of human naïve CD25- T-cells can result in FOXP3 expression and associated suppressor activities 24, 57, 103. The data described above supports the concept that nTreg cells, with high FOXP3 expression and suppressive activity, can develop from conventional peripheral T-cells under specific in vivo activation conditions 24. It is an intriguing concept, since it opens up the possibility for novel therapeutic strategies aiming at stable induction of nTreg/Tr1 cells. However, this may be a more complex goal than the data suggests since glucocorticoid treatment in asthma does not lead to long-term suppression.
NATURAL T-REGULATORY CELLS AND MOUSE MODELS OF ALLERGIC ASTHMA
Mouse models of allergic asthma have been very helpful in the understanding of the immunological mechanisms of IgE regulation, Th1/Th2 balance, eosinophilic airway inflammation and development of AHR. Furthermore, many experimental therapeutic approaches have first been examined in mouse models. The mouse has also been widely used with regards to the role of Treg cell subsets in immunoregulation, in particular in models of autoimmune and infectious diseases. However, mouse studies on the role of Treg cells in different aspects of allergic disease and asthma are less abundant and often contradictory. In different, sometimes rather artificial, models both stimulatory and inhibitory effects of nTreg cells have been observed on IgE production, Th2 responses and airway eosinophilia, whereas no effect on AHR has been observed 104–107. In the studies cited above the nTreg cells were characterised by the expression of CD25 only and in most models endogenous antigen expression or a T-cell receptor transgenic system was used. Therefore, it cannot be excluded that some of these observations are confounded by the involvement of aTreg cells, Tr1 cells in particular 107. Recently, van Oosterhout et al. 108 demonstrated in a BALB/c mouse model of asthma that nTreg (CD4+CD25+CD45RBlowGITR+) cells are present in the lung tissue of sensitised mice and increase upon allergen inhalation. Inhibition of nTreg cell suppression by ligation of GITR 109, 110 potentiated respiratory allergen-induced AHR and IgE production, and Th2 cytokine levels in lung lavage fluid, but not airway eosinophilia 108. Increased levels of Th2 cytokines were also found in recall cultures of lung T-cells in the presence of nTreg cell-inhibiting anti-GITR. Data was suggestive of the suppressive potential of nTreg cells. However, since GITR was recently demonstrated on activated conventional CD4+ T-cells and can function as a costimulatory molecule 111–114, it cannot be excluded that the potentiation of asthma manifestations by GITR ligation is due to potentiation of Th2-cell effector function. Interestingly, the natural ligand of GITR (GITRL) is expressed by resting DCs, macrophages and B-cells, which strongly suggests that APCs modulate the immunosuppressive function of nTreg cells 110, 115–117. Considering these findings, local stimulation of nTreg cell number or function in the airways may be a promising therapeutic approach that deserves further exploration.
ADAPTIVE T-REGULATORY CELLS AND MUCOSAL TOLERANCE
In the mouse, primary exposure to the model antigen, ovalbumin, via the respiratory mucosa either by inhalation or intranasal administration, has been shown to induce tolerance to the generation of an IgE, but not Th1-associated IgG2a, response and to subsequent antigen-induced airway manifestations of asthma 118–121. This form of tolerance appeared to be mediated by the induction of Treg cells, but controversy exists regarding the subset(s) involved 122. Active induction of Tr1-like cells by IL-10 producing DCs 82, 83 and of Th3/nTreg like cells by a TGF-β-dependent mechanism 123 has been observed after primary ovalbumin exposure. An explanation for these different observations may be the dose of ovalbumin used. Whereas Akbari et al. 82 treated mice intranasally with a relatively high dose, Ostroukhova et al. 123 exposed mice to a low dose of aerosolised ovalbumin. In contrast to ovalbumin, intranasal treatment with a low dose HDM extract has been shown to induce allergic sensitisation and subsequent development of allergic airway inflammation and AHR 124. Interestingly, Cates et al. 125, 126 showed that allergic sensitisation and subsequent inflammation in response to intranasal administration of HDM were mediated by granulocyte macrophage-colony stimulating factor (GM-CSF). Vice versa, it has been shown that induction of GM-CSF production in epithelial cells by adenoviral GM-CSF gene transfection induces allergic sensitisation and allergic inflammation upon primary intranasal ovalbumin exposure 127. The different capacities of allergens, such as HDM and pollen, and the innocuous antigen, ovalbumin, to cause allergic sensitisation may be related to proteolytic capacities of the allergens. These trigger GM-CSF production by airway epithelial cells, thus causing DCs maturation 128–131. As mentioned earlier, immature DCs, in particular when they produce IL-10 or TGF-β, are potent inducers of aTreg cells. Moreover, suppression by nTreg cells is critically dependent on immature DCs and is readily reversed by maturation of these cells in response to GM-CSF 132. Another epithelium-derived factor that may be involved in this process is thymic stromal lymphopoietin (TSLP) which can activate DCs to produce Th2-attracting chemokines and to induce naïve CD4+ and CD8+ T cell differentiation into effector cells with a typical pro-allergic phenotype 133–135. However, it is unclear at present whether airway epithelium releases TSLP upon contact with proteolytic allergens. A major question regarding allergic sensitisation versus tolerance induction in humans is why airborne allergens induce allergic sensitisation in atopic, but not in nonatopic, individuals 132. In addition to the proteolytic capacities of many airborne allergens, other factors such as the genetic background are involved. More knowledge about these factors may reveal new opportunities for prophylactic treatment of children with higher risks of developing allergic diseases.
T-REGULATORY CELLS AND THE HYGIENE HYPOTHESIS
The postulate of the hygiene hypothesis that there would be an inverse relation between the extent of exposure to microbial agents during early childhood and the prevalence of Th2-mediated allergic diseases, has been supported by substantial evidence. However, the supposed underlying mechanism, that Th1 reactions as induced by microbial agents would suppress development of allergen-induced Th2 reactions, appear untenable. Notably, helminth infections that elicit strong Th2 reactions with high levels of IgE appeared protective against IgE-mediated allergies 19. Moreover, the prevalence of Th1-mediated autoimmune diseases has risen in parallel with Th2-mediated allergies 21, although a decrease would have been expected, if Th1 and Th2 reactions would have effective mutually inhibitory activity. The new concept, therefore, assumes that the rise in allergic and autoimmune diseases is the result of a decrease in immunoregulatory control of Th1 as well as Th2 reactions, probably due to decreased exposure to microbial agents 8, 19, 136. How microbial agents would restrain development of untoward immune responses is not fully understood at present, but there is increasing awareness that stimulation of Treg cell numbers or function, by microbial agents, may be involved. One of the mechanisms by which microbial agents would facilitate Treg cell induction has already been referred to in this review. Several regulatory-type PAMPs stimulate development of tolerogenic DCs by inducing IL-10 and/or TGF-β production in these cells. However, since other PAMPs may promote development of mature, immunogenic DCs 65, 137, current understanding of the net result of DC exposure to intact microorganisms is presently insufficient. Nevertheless, pathogen-specific Tr1 cells were found in humans or mice upon infection with viruses (hepatitis C, Epstein-Barr), bacteria (Bordetella pertussis, Helicobacter hepaticus, Mycobacterium tuberculosis) or the parasite, Onchocerca volvulus as reviewed by Mills 30. Likewise, nTreg cells of undetermined specificity were found in HIV-infected humans and mice infected with viruses (Herpes simplex), bacteria (H. hepaticus, H. pylori), parasites (Schistosoma mansoni, Leishmania major, Plasmodium spp., Litomosoides sigmodontis), or fungi (Pneumocystis carinii, Candida albicans) 30, 138. In many of the mouse infection models, in vivo depletion or transfer studies have shown that the pathogen-induced Treg cells inhibit specific T cell responses and inflammatory immunopathology and prevent complete pathogen elimination from the host 30. The Treg cell-dependent pathogen survival is not necessarily harmful to the host, because, as demonstrated for L. major, it leads to persistent immunity against re-infection, while complete eradication leads to loss of immunity 139. Thus, the Treg cell-induced persistence of low numbers of pathogens in the body would result in persistently elevated levels of Treg cell activity. This, in turn may help to prevent the development of untoward allergic and autoimmune reactions by the so-called bystander suppression, since the suppressor effector function of nTreg cells and suppression by IL-10 derived from Tr1 cells are antigen nonspecific 26, 140, 141. In other words, Treg cell-inducing pathogens may elevate the level of T cell regulation and so prevent sensitisation to environmental allergens, as schematically depicted in figure 3⇓. In addition to various pathogens, the intestinal flora may be a persistent cause of increased Treg cell surveillance in the body. This is clearly indicated by the observation that murine CD4+ T cells isolated from enteral lamina propria, mesenteric lymph nodes, or peripheral blood did not react to an extract of the sonicated lumenal content of colon and cecum, while vigorous proliferation was seen upon depletion of the CD4+CD25+ T cells 142.

Schematic representation of the hypothetical effect of environment and gut microflora-induced T-regulatory (Treg) cell surveillance on the manifestation of allergic responses. a) In a less “hygienic” environment or in the case of a healthy, lactic acid bacteria-rich gut flora (▒), the level of Treg cell surveillance (═) is high enough to restrain manifestation of an allergen-specific T-cell activation. Pathogens generally surpass T-cell regulation in the first stages of infection by providing danger signals, resulting in T-cell-dependent adaptive immune responses aimed at eliminating the pathogen. b) In Western societies with high levels of hygiene and a disturbed composition of gut flora (▒), Treg cell surveillance (═) is decreased, so allowing development of conventional T cell-dependent allergen-specific responses, for instance increased levels of specific immunoglobulin E against a certain, relatively weak allergen (¶) or manifest symptoms of allergic disease induced by another allergen, which is more potent for the host (#). Peaks denote the strength of specific T-cell responses to allergens or pathogens. Reprinted, with permission, from the Annual Review of Immunology, Volume 22 © 2004 by Annual Reviews www.annualreviews.org.
Whether all or only certain species of the intestinal bacterial flora contribute to Treg cell surveillance in the body is not known at present. However, regarding the protective activity of probiotic lactic acid bacteria against development of allergic diseases 143, 144 and observations that children with higher counts of lactic acid bacteria in their faeces had a lower prevalence of allergic disease 145, it is tempting to speculate that the level of Treg cell activation by intestinal flora is influenced by its composition. Besides commensal intestinal bacteria, exposure to the orofaecal or foodborne pathogens, hepatitis A virus, H. pylori, and Toxoplasma gondii, as evidenced by seropositivity, appeared to be associated with a lower prevalence of atopy in Italian male cadets, especially in the case of seropositivity for at least two of the pathogens 146. In the cumulative seropositives, allergic asthma appeared rare and allergic rhinitis infrequent. Since no associations were found between (cumulative) seropositivity for pathogens transmitted by other routes and the prevalence of atopy, it was concluded that inadequate stimulation of the gut-associated lymphoid tissue would enhance the risk of atopy. This conclusion could not be substantiated by a study of Danish subjects aged 15–69 yrs. Whereas it confirmed a lower prevalence of atopy in subjects with seropositivity for at least two of the above-mentioned orofaecal or foodborne pathogens, it did not find an association between atopy and cumulative seropositivity for the intestinal pathogens, Clostridium difficile, Campylobacter jejuni and Yersinia enterocolitica 147. Thus, although it is still not clear whether stimulation of gut-associated lymphoid tissue is essential for providing protection against the development of atopy and atopic disease, the nature of stimulation of the immune system by intestinal pathogens is essential. Whereas all gastrointestinal pathogens are associated with food or personal hygiene, H. pylori and T. gondii differ from the others by generally establishing chronic interactions with their host. Treg cells are probably instrumental to this chronicity and were shown to be induced by H. pylori. These have not yet been demonstrated for T. gondii, but the observation that the in vitro CD4+ T cell hyporesponsiveness induced by this pathogen could be reversed with IL-2, points to the possible involvement of Treg cells 148. The mechanism of the protective effect of hepatitis A virus, that does not induce a chronic infection, is probably different. It has been related to the presence of TIM-1, a specific receptor for this virus on CD4+ T cells, since carrying particular polymorphic variants of TIM-1 appeared strongly associated with protection against atopy, but only if carriers had serological evidence of past infection 149. Therefore, it was suggested that hepatitis A virus would favourably change T-cell differentiation in carriers of the protective TIM-1 variants. Dependence of protection on these variants may also explain why some studies did not find an association between past hepatitis A virus infection and atopy 150, 151.
Inhibition of allergic responses by microbe-specific Tregs through the bystander suppression is not the only inhibitory mechanism. Microorganisms were also shown to act as an adjuvant to the induction of allergen-specific Treg cells. Treatment of ovalbumin-sensitised mice with a mixture of ovalbumin and heat-killed Listeria monocytogenes in incomplete Freund's adjuvant was shown to inhibit development of AHR or to reverse already established AHR. Moreover, it reduced airway inflammation, eosinophilia, mucus production, and antigen-specific IgE and IL-4 production, while increasing specific IFN-γ production 152. The inhibition was shown to be mediated by Th1-like Foxp3-expressing T cells, a novel type of aTregs 153. In another mouse study, subcutaneous <1?twb=.25w?>administration of the heat-killed saprophyte, Mycobacterium vaccae, in an aquous vehicle 3 weeks before intraperitoneal <1?twb=.25w?>sensitisation with ovalbumin absorbed to alum inhibited subsequent induction of eosinophilic airway inflammation and AHR by respiratory ovalbumin challenge 154. Effects have been attributed to ovalbumin-specific, CD4+CD45RBlow aTreg cells that mediated suppression by IL-10 and TGF-β and CD11c+ APCs in lung tissue producing IL-10 and TGF-β, and IFN-α 155. Although this approach is still far from being used as a practical application in humans, the demonstration that inhibitory allergen-specific Treg cells can be induced by means of a suitable adjuvant and treatment schedule holds promise for the future.
T-REGULATORY CELLS AND ALLERGEN IMMUNOTHERAPY
Although allergen immunotherapy has been practiced since 1911 156, recent developments in purification of extracts and the general understanding of the mechanisms which bring about its action have increased its applicability and hold promise for improved, future treatment of allergic diseases. Allergen immunotherapy consists of repeated subcutaneous injections with increasing doses of allergen. It leads, in the majority of allergic patients, to a reduction of allergen-induced inflammation and allergic symptoms, reduced medication requirement, and a marked improvement in lung function, as has been summarised in a meta-analysis 157. The clinical and anti-inflammatory effects of allergen-immunotherapy can last years after discontinuation, which makes this therapy the only current disease-modifying treatment for allergic diseases 158. However, despite the efficacy of this classical form of therapy for the treatment of monoallergic asthma patients, it seldom results in complete alleviation of all asthma symptoms 157, 159, 160. Moreover, in comparison with the easy to use and clinically very efficacious treatment with inhalation glucocorticoids, immunotherapy is hampered by side-effects at high allergen doses, especially in asthmatics and the necessity for frequent allergen injections over prolonged periods of time discourage physicians from applying immunotherapy on a large scale.
The beneficial effects of allergen immunotherapy are likely to be mediated through the reduction of allergen-induced inflammation. Immunological processes underlying these effects remain largely unknown at present, but several potential mechanisms have been postulated 161, 162. As T cells orchestrate the inflammatory response, efforts since 1990 have been focused on activation and differentiation of T-lymphocytes in relation to allergen immunotherapy. Both allergen-specific hyporeactivity and a shift in the cytokine profile of reacting T-lymphocytes (Th2 to Th1) have been proposed as underlying mechanism (fig. 4⇓) 164–166. In 1998, Akdis et al. 167 were the first to provide evidence for a role of IL-10 and Tr1 cells in the beneficial effect of allergen immunotherapy in bee-venom allergic patients. They observed that the therapy-induced decrease in T-cell proliferation and production of IL-5 and IL-13 in response to bee-venom stimulation in vitro could be fully antagonised by neutralising IL-10. These data suggest an important role for IL-10-producing Tr1 cells in allergen immunotherapy. Findings were extended by recent immunotherapy studies with airborne allergens that showed increases in allergen-specific IL-10-producing Tr1 and TGF-β-producing Th3 type cells in the blood or airway tissue 168–170. Whether the therapy-induced cytokines mediate the immunosuppressive effects remains unknown at present. Interestingly, in a mouse model of allergen immunotherapy 171, Vissers et al. 172 recently demonstrated that blocking of the IL-10 receptor largely abrogated the beneficial effects of therapy, namely the reduction of bronchoalveolar lavage eosinophil numbers and AHR to methacholine.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immune deviation associated with the beneficial effects of allergen immunotherapy. After successful immunotherapy, increased serum levels of allergen-specific “blocking” immunoglobulin (Ig)G antibodies, in particular IgG4 and IgA, are observed, whereas IgE production is suppressed. The changes in serum Ig isotypes are paralleled by a shift in allergen-specific T-cells from T-helper cells type 2 (Th2) to Th1 and the appearance of allergen-specific adaptive T-regulatory cells (aTreg: Th3 and T-regulatory cell type 1), characterised by an increased production of interferon (IFN)-γ, transforming growth factor (TGF)-β and interleukin (IL)-10, respectively. AHR: airway hyperresponsiveness. Copyright © 2001 Massachusetts Medical Society. Adapted with permission from Kay et al. 163.
In humans, increased IL-10 production after immunotherapy is not limited to T cells. B cells, monocytes and macrophages also produce this cytokine after immunotherapy 167, 168. All three types can present antigen and by producing IL-10 they may be involved in the suppressive effects of immunotherapy, either directly or through the induction of Tr1 cells. In agreement herewith, it was recently demonstrated that antigen-pulsed macrophages suppress antigen-induced airway manifestations of asthma in an IL-10 dependent fashion 173. IL-10 has been shown to decrease IgE production and to enhance IgG4 production in human B-lymphocytes in vitro. So, the immunotherapy-induced production of IL-10 by Tr1 cells may also be responsible for the observed increase in production of allergen-specific IgG4 antibodies 169, 174. These antibodies may contribute to the immunosuppressive effects of immunotherapy by blocking IgE-facilitated antigen-presentation 169, 175. In addition, at least in the mouse, immune-complexes of antigen and IgG may induce Fcγ receptor-mediated IL-10 production by macrophages 176. These data suggest a feedback stimulatory loop between IL-10, IgG4 and generation of Tr1 cells.
It can be concluded that the positive effects of allergen immunotherapy are associated with the appearance of Tr1 and Th3 cells (fig. 4⇑). The mechanism by which these aTreg cells are induced by immunotherapy is presently unknown. However, if the details of this immunological pathway will be elucidated, novel strategies to improve immunotherapy may become available, not only for better treatment of allergic diseases, but also for efficient treatment of allergic asthma possibly with long-term improvement of disease. Well-described pathways to induce aTreg cells are antigen presentation to T-cells by immature DCs or the presence of IL-10 or TGF-β in the local microenvironment. In this respect, combination of allergen immunotherapy with administration of compounds that inhibit DC maturation, such as 1,25(OH)2-VitD3 or glucocorticoids, may be an interesting strategy to pursue.
CONCLUSION
It is apparent that Treg cells may offer an explanation for many of the (pre)clinical observations that cannot be adequately explained by a disturbed Th1/Th2 balance. Although not yet studied extensively, data indicate that Treg cells interfere with the development of asthma and other allergic diseases at different stages, such as allergic sensitisation, progression to established allergic disease and asthma and severity and persistence of disease. Treg cells may also offer an explanation for the hygiene hypothesis. Microbial ligands can instruct DCs to become tolerogenic or immunogenic and likewise, induce protective Treg cells or diseases-inducing Th1 or Th2 cells. In addition, microbial ligands may directly promote nTreg cell function through TLRs or abrogate its function by inducing IL-6 expression in DCs 73, 132, 177, 178. At the basis of this complex and intriguing interaction between innate and adaptive immune responses, is the genetic background. Polymorphisms in TLR-2 and CD14 have been demonstrated to influence atopy and asthma susceptibility 136, 179–181. In addition, polymorphisms of CTLA-4, which is important for nTreg cell function, have been associated with asthma 182–185.
Therapeutic opportunities to exploit the immunosuppressive effects of T-regulatory cells for prophylactic or therapeutic treatment and allergen immunotherapy may become feasible when current knowledge progresses. Although glucocorticoids appear to induce transient natural T-regulatory cells/T-regulatory type-1 like cells, stable induction may require additional, yet unknown, signals. Ex vivo generation and expansion of T-regulatory cells followed by autologous transfer is currently being explored for the treatment of autoimmune diseases and transplant rejection 98, 186–188. When such treatment leads to long-term improvement of disease, it may be valuable to explore its application for the treatment of allergic asthma, in particular for those patients that are difficult to treat with currently available drugs. Alternatively, blocking the signals that restrain natural T-regulatory cell function, such as the natural ligand of GITR and interleukin-6 may potentiate the suppressive function of natural T-regulatory cells in vivo.
Note added in proof: it is now feasible to determine intracellular FOXP3 protein expression by flow cytometry in order to directly identify nTreg cells 189.
- Received February 1, 2005.
- Accepted June 10, 2005.
- © ERS Journals Ltd
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵










Jump To
- Article
- Abstract
- NATURAL T-REGULATORY CELLS
- FOXP3 AND NATURAL T-REGULATORY CELLS
- ADAPTIVE T-REGULATORY CELLS
- ROLE OF DENDRITIC CELLS IN GENERATING ADAPTIVE T-REGULATORY CELLS
- T-REGULATORY CELLS AND ASTHMA
- GLUCOCORTICOIDS AND T-REGULATORY CELLS
- NATURAL T-REGULATORY CELLS AND MOUSE MODELS OF ALLERGIC ASTHMA
- ADAPTIVE T-REGULATORY CELLS AND MUCOSAL TOLERANCE
- T-REGULATORY CELLS AND THE HYGIENE HYPOTHESIS
- T-REGULATORY CELLS AND ALLERGEN IMMUNOTHERAPY
- CONCLUSION
- References
- Figures & Data
- Info & Metrics