





Abstract
Evidence has been accumulating that chronic inactivity leading to muscle disuse is unlikely to be the only explanation for the peripheral muscle dysfunction of chronic obstructive pulmonary disease (COPD) patients.
Although a new concept of myopathy was recently proposed, the question of disuse and/or a form of myopathy is still being debated. This review proposes definitions for the terms used in this debate, discusses the relevant studies and concludes that the evidence points to a myopathy associated with muscle disuse in COPD.
COPD myopathy implies pharmacological and/or pathophysiological mechanisms that need to be identified in order to optimally orient therapeutic strategies. The literature indicates that corticosteroids, inflammation, hypoxaemia and oxidative stress are among the factors contributing to COPD muscle dysfunction, but their relative contributions have not been fully elucidated.
This review presents the advances in understanding each of these mechanisms, especially the data showing that muscle oxidative stress occurs and contributes to muscle dysfunction in chronic obstructive pulmonary disease. The current review also reports the studies that have elucidated the molecular mechanisms underlying this stress in chronic obstructive pulmonary disease by demonstrating alterations in oxidant and/or antioxidant systems. Finally, the review considers how inflammation and hypoxaemia may trigger oxidative stress in chronic obstructive pulmonary disease muscles and presents the therapeutic modalities that should be proposed to prevent it.
Peripheral muscle dysfunction, which is now recognised as one of the main systemic effects of chronic obstructive pulmonary disease (COPD), contributes greatly to the exercise intolerance and reduced quality of life of COPD patients 1–4.
Despite the considerable advances in research since the 1980s, the causes of peripheral muscle dysfunction have yet to be fully elucidated. Based on the theoretical model of Young 5, this dysfunction in COPD has primarily been explained by the sedentary lifestyle commonly observed in patients. In support of this notion, a series of studies clearly showed that physical training improves peripheral muscle function, exercise tolerance and quality of life in COPD 6–8. At the time of these studies, the terms “detraining”, “deconditioning” and “muscle disuse” were used to characterise the peripheral muscle alterations. However, the question today is the following: is the model of Young 5 a restrictive view of the peripheral muscle dysfunction of COPD patients? In light of the most recent research, the current authors believe so. Indeed, evidence has been accumulating that a sedentary lifestyle is unlikely to be the only mechanism underlying this dysfunction, suggesting the role of an intrinsic muscle disease or myopathy 9, 10. The term “steroid-induced myopathy” was first proposed in relation to COPD in 1990 11 and it was employed again in 1996 12. However, a more general reference to “myopathy” in COPD was made at a European Respiratory Society seminar in 1999 13 and the term then appeared in the literature in 2002 14. Although a clear distinction between “myopathy” and “muscle disuse”, “detraining” or “deconditioning” needs to be drawn, there is currently no agreement on how to do so. To cite a verse from Horace's Ars Poetica (17–13 BC), “Verbaque provisam rem non invita sequentur”, which can be translated as: “The subject once foreknown, the words will easily follow”. The current authors believe that in order to resolve the debate on COPD peripheral muscle, the first step must be the clear definition of each of the relevant terms. For example, is disuse a synonym for, or a cause of, deconditioning? Are disuse and myopathy close in meaning or not? The first objective of this review will, thus, be to consider the meanings of these terms and to propose standard definitions, and then to review the latest studies that address the question of whether the peripheral muscle dysfunction of COPD is caused by disuse, detraining, deconditioning and/or a form of myopathy.
The latest data in the literature argue in favour of a myopathy, which implies at least one or more pharmacological and/or pathophysiological mechanism. As will be seen in the second part of this review, various studies have already reported long-term steroid use, systemic inflammation and hypoxaemia as potential contributors to the peripheral muscle dysfunction in patients with COPD 3, 12, 15, 16. Yet even taken together, these mechanisms seem insufficient to explain all the abnormalities observed in the peripheral muscles of COPD patients 9, which leads one to suspect there are still other causes.
Recently, oxidative stress has been investigated as a potential mechanism in at least a part of the dysfunction. Oxidative stress, which is defined as an imbalance between the formation of and protection against reactive oxygen or nitrogen species (RONS), can result in structural and functional muscle abnormalities, quite similar to those observed in COPD peripheral muscle. Furthermore, evidence has also been accumulating that oxidative stress occurs in the lung and plasma of COPD patients and contributes to the development and progression of the disease 17, 18.
The second objective of this review will be to present some of the most recent research on muscle oxidative stress in COPD, particularly those studies focused on determining whether oxidative stress occurs in the resting and/or exercising peripheral muscle of COPD patients and whether it contributes to muscle dysfunction. This review will thereafter discuss the potential mechanisms underlying this stress and its relation to the pathophysiology of COPD. Lastly, this review presents the therapeutic strategies suggested by the literature that should be proposed to prevent muscle oxidative stress in these patients.
GENERAL DEFINITIONS
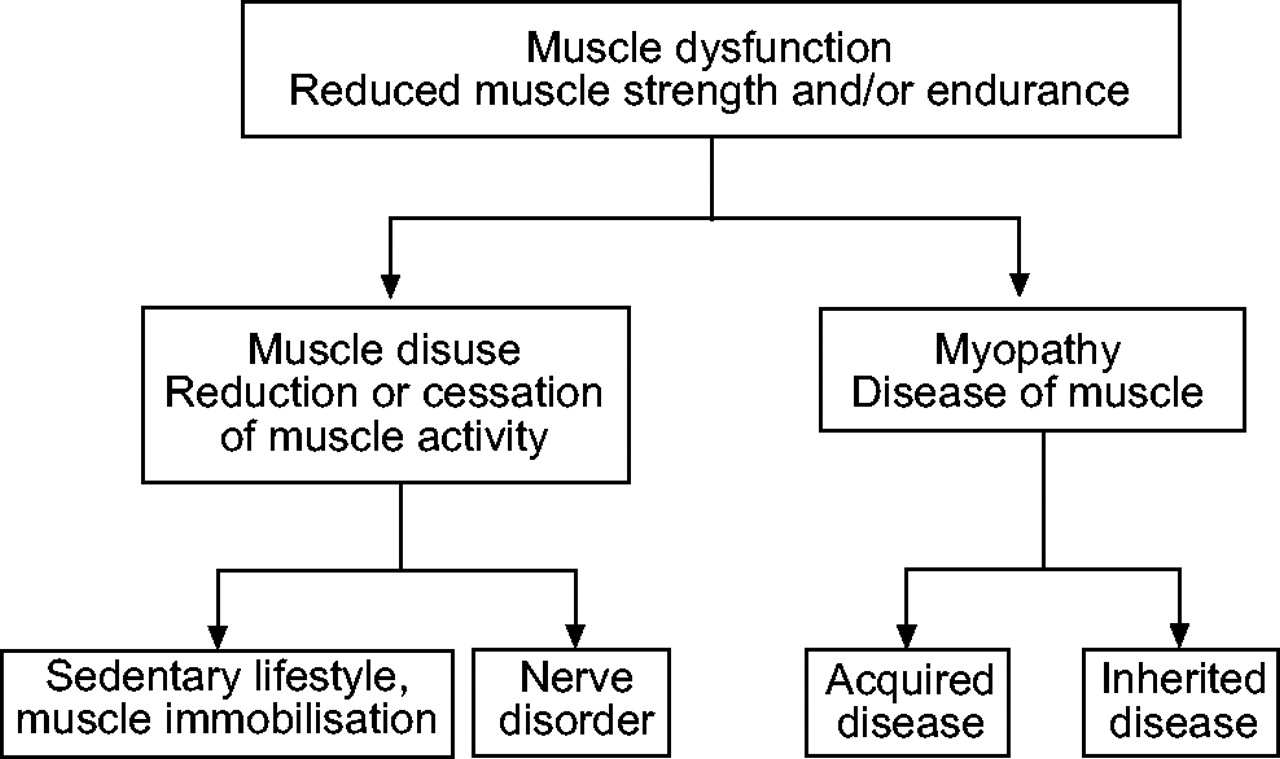
As its etymology suggests (from Latin: “functio” = performance), muscle dysfunction is characterised by a reduction in muscle strength (defined as the capacity of the muscle to develop maximal force) and/or endurance (defined as the capacity of the muscle to maintain a certain force over time), both of which are determinants of performance 19. Once a phenomenon is defined, however, it is necessary to determine its mechanisms. In the case of muscle dysfunction, the general assumption is that it results from muscle disuse and/or myopathy (fig. 1⇓). However, these are two distinct mechanisms that need to be clearly distinguished.

Muscle disuse and myopathy are two distinct mechanisms that can both lead to peripheral dysfunction.
Definition of muscle disuse
Muscle disuse is defined as a reduction or cessation of muscle activity 19 that results from a sedentary lifestyle, voluntary muscle immobilisation (bed rest, hind limb suspension, and so on) or nerve disorder. Disuse leads to several adaptive changes, commonly referred to as “musculoskeletal deconditioning”, which mainly include a reduced proportion of type I fibres and oxidative enzyme capacity, muscle fibre atrophy and reduced muscle capillarisation. Together, these alterations result in reduced muscle endurance and strength 20. It is important, however, to bear in mind that the magnitude of musculoskeletal deconditioning is greatly dependent on the pre-existing morphological and biochemical properties of the muscles, as well as the cause and duration of the disuse. Here it is clear that muscle disuse and deconditioning are causally related, so that the two terms cannot be used equivocally. The term “muscle detraining” is specifically used in the field of sports sciences and is defined as the partial or complete loss of training-induced adaptations in response to an insufficient training stimulus or reduced muscle activity. However, the overall meaning and the characteristics of musculoskeletal detraining and deconditioning are quite similar, so that in fact no clear distinction exists between the two processes. Lastly, skeletal muscle has a remarkable plasticity that allows it to adapt to variable levels of functional demand, so that disuse-induced muscle changes are usually reversible with an adequate period of physical training 21.
Definition of myopathy
Myopathy (from Greek: μυσ, παθη, mus, pathê: pathology of the muscle) is defined as a “disease of muscle unrelated to any disorder of innervation or the neuromuscular junction”, thus implying that the muscle is in a pathological condition and cannot function normally 19. Different forms of myopathy have been determined and these are classified into specific groups depending on the aetiology, such as hereditary, metabolic, inflammatory, endocrine, and toxic causes. However, these aetiologies are not exhaustive and do not exclude the possibility of other causes. Whatever its form, myopathy is characterised by several metabolic, morphological and/or structural muscle abnormalities that lead to dysfunction. It is interesting to note that myopathological alterations can be either equally or heterogeneously distributed among muscle groups. Lastly, myopathy-induced structural, metabolic and functional alterations have been reported to be only partially reversible with appropriate therapeutic strategies 22–24.
DISUSE AND/OR A FORM OF MYOPATHY IN COPD
In patients with COPD, peripheral muscle dysfunction is clearly evidenced by the significant reduction in both strength and endurance in the ambulatory muscles of these patients in comparison with healthy subjects 25–29. Other studies have shown several morphological, structural and biochemical abnormalities that could contribute to the dysfunction 1–4, 16. However, there is currently no accord on the nature of this dysfunction. Is the dysfunction of COPD peripheral muscle due to disuse alone, a form of myopathy, or both?
The hypothesis of muscle disuse implies that patients with COPD present either a nerve disorder or a reduction in muscle activity due to a sedentary lifestyle. Studies using magnetic stimulation of the femoral nerve have demonstrated that neuromuscular transmission is preserved in the ambulatory muscles of these patients 30, 31. In contrast, COPD muscle undergoes varying degrees of disuse because of a sedentary lifestyle 29 engendered by dyspnoea and/or psychosocial disorders 5, 32.
If disuse alone accounts for the peripheral muscle alterations observed in COPD, the muscle properties should be quite similar in COPD patients and healthy long-term sedentary individuals. The first step then is to define these individuals: a long-term sedentary subject should be someone who has not engaged in any physical training or regular physical activity for many years, and this should be the case for as many years as possible. To the current authors' knowledge, only one study specifically recruited healthy individuals on the basis of long-term sedentary lifestyle. Houmard et al. 33 recruited subjects (≈60 yrs) who reported always having had a sedentary lifestyle with no prolonged periods of regular exercise, especially in the previous 2 yrs.
These healthy subjects were characterised by a severe reduction in maximum oxygen uptake (V′O2,max (24 mL·min−1·kg−1)) and 41% of type I fibres in the vastus lateralis. In line with this finding, other studies have reported 40–50% of oxidative fibres in the vastus lateralis of healthy sedentary subjects (≈60 yrs) recruited on the basis of a specific physical activity questionnaire and/or a similar low value of V′O2,max (≈20 mL·min−1·kg−1) 9, 34–36. Moreover, it is important to consider that the percentage of type I fibres observed in the vastus lateralis of age-matched very active healthy subjects ranges 60–65% 37. These results clearly indicate that muscle disuse can induce a one-third reduction in the proportion of type I fibres (fig. 2⇓).
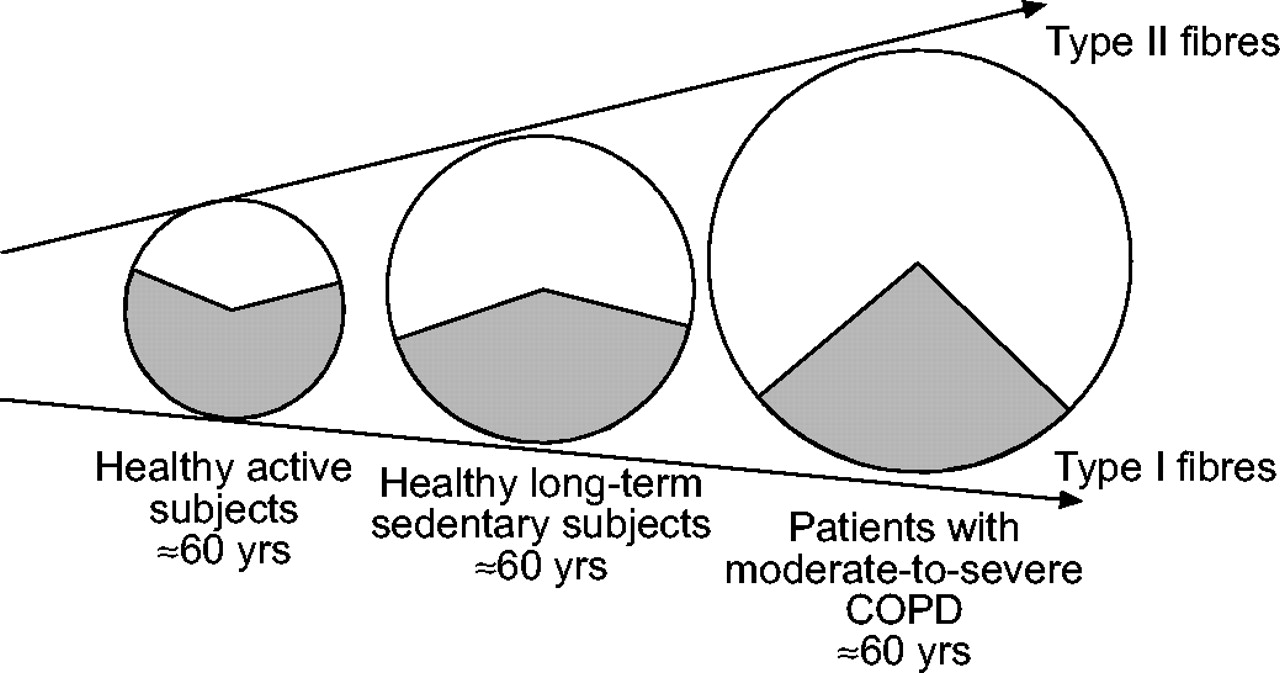
Histochemical proportion of type I ( ) and type II (□) fibres observed in the vastus lateralis of healthy active and long-term sedentary subjects as compared with patients with moderate-to-severe chronic obstructive pulmonary disease (COPD). The proportion of type I fibres is almost 50% lower in COPD as compared with long-term sedentary healthy subjects, indicating that disuse is unlikely to be the only mechanism explaining the peripheral muscle abnormalities in these patients.
) and type II (□) fibres observed in the vastus lateralis of healthy active and long-term sedentary subjects as compared with patients with moderate-to-severe chronic obstructive pulmonary disease (COPD). The proportion of type I fibres is almost 50% lower in COPD as compared with long-term sedentary healthy subjects, indicating that disuse is unlikely to be the only mechanism explaining the peripheral muscle abnormalities in these patients.
A large number of studies have evaluated the morphological properties of the vastus lateralis in moderate-to-severe COPD patients in comparison with age-matched healthy long-term sedentary subjects and have reported striking differences. Indeed, all have reported that the oxidative fibres averaged 23% (even down to 19%) in the patients, whereas they averaged, as expected, 44% in the healthy sedentary subjects 26, 34–36, 38. These results clearly indicate a two-thirds reduction in the proportion of type I fibres in the peripheral muscles of COPD patients, which is unlikely to be accounted for by muscle disuse alone (fig. 2⇑). Furthermore, the only studies of the effect of physical training on peripheral muscle typology in COPD reported that the proportion of type I fibres remains unchanged after 12 weeks of endurance training 39–41. In addition, the reduced oxidative activity of citrate synthase observed in the vastus lateralis of COPD patients (38% lower than in healthy subjects) was partially reversed by an endurance training programme (an increase of 16%), but the patient values nevertheless failed to return to the normal values of healthy subjects 8.
Finally, recent studies strongly suggest that the quadriceps endurance capacity of COPD patients only weakly correlates with daily physical activity scores. Serres et al. 29 reported a moderate, but significant correlation coefficient between physical inactivity as quantified with a questionnaire and quadriceps endurance capacity in COPD, while two other studies found no relationship between these two parameters 25, 42. In addition, a significant difference in quadriceps muscle endurance has been reported between COPD patients and healthy subjects, despite similar levels of physical activity 14.
Disuse is almost certainly an important factor in the skeletal muscle dysfunction of COPD. However, the most recent data indicate that disuse only partly explains the severe morphological, metabolic and functional abnormalities observed in COPD peripheral muscle; instead, the data support the concept of an intrinsic muscle disease, that is to say, a myopathy.
Therefore, it seems very likely that both muscle disuse and myopathy contribute to peripheral muscle dysfunction, at least in patients with moderate-to-severe COPD. The relative contribution of each process, however, needs to be determined.
POTENTIAL FACTORS OF MYOPATHY IN COPD
Once the contribution of myopathy to the peripheral muscle dysfunction of COPD has been convincingly demonstrated, it becomes necessary to elucidate its underlying mechanisms. This is fundamental to identifying the form of the myopathy (toxic, inflammatory, metabolic, endocrine and/or other?) and will then guide the development of appropriate therapeutic strategies. Several studies have already reported the potential contributions of different mechanisms, these being principally steroid use, inflammation and hypoxaemia. The objective of this part of the review is to consider the latest advances in understanding these mechanisms and then to discuss whether they adequately account for the myopathy in patients with COPD.
Corticosteroid use
Since patients with COPD are usually treated with corticosteroids either as “short-burst” therapy for acute exacerbations or as long-term, low-dose “maintenance” therapy, several authors have investigated the possibility that COPD peripheral muscles experience a toxic myopathy: the so-called “steroid-induced myopathy”. Evidence was first shown that long-term high doses of steroids mediate detrimental effects on the structure and function of COPD peripheral muscle 43. Thus, the next logical step was to determine whether long-term low doses, as often prescribed in severe COPD, also induce peripheral muscle abnormalities. The literature data indicate that the answer is yes. A significant reduction in quadriceps strength, strongly associated with quadriceps atrophy, was reported in COPD patients who received chronically low doses of steroids (the average daily dose of prednisone amounted to 5 mg) in the 6-month period preceding the study as compared with patients unexposed to this medication 28.
Furthermore, Decramer et al. 44 reported severe quadriceps weakness that was significantly correlated with the average daily dose of steroids taken by patients (an average of 4 mg of methylprednisolone), mostly as a short-burst therapy implemented during acute exacerbations over the preceding 6 months. In their study, the loss in quadriceps strength was out of proportion to the loss in total body muscle mass, thus suggesting that the muscle weakness induced by repetitive bursts of steroid ingestion was unlikely to be explained by muscle atrophy alone. In line with this notion, the same group further demonstrated that long-term repetitive bursts of steroid ingestion (methylprednisolone) was able to induce a global picture of myopathy, comprising both functional and severe histological peripheral skeletal muscle abnormalities 12. These results suggest that the myopathic effects of repetitive bursts of steroid therapy might be greater than those of continuous low-dose therapy. A word of caution is necessary here; however, it is important to recall that methylprednisone has been reported as having a greater potential to alter skeletal muscle structure and function than prednisone 45. Furthermore, the above-mentioned studies were not able to distinguish the effects of steroid treatment from the effects of the frequent exacerbations for which the steroids had been administered.
In an attempt to resolve this ambiguity, Hopkinson et al. 46 very recently showed that a 2-week course of oral corticosteroids (o.d. 30 mg prednisolone) did not reduce quadriceps strength or impair metabolic parameters during exercise in stable outpatients with moderate-to-severe COPD. Since the corticosteroid dose used in their study was similar to that conventionally used to treat acute exacerbations, it appears that in the absence of particular aspects of exacerbation (enhanced systemic inflammation, immobility), an isolated short-term treatment with corticosteroids does not in itself cause peripheral muscle weakness in COPD.
Systemic inflammation
It is now clearly established that COPD is associated not only with an abnormal inflammatory response of the lung, but also with systemic inflammation characterised by enhanced activation of circulating inflammatory cells (neutrophils and lymphocytes), greater expression of surface adhesion molecules in circulating neutrophils, and increased plasma levels of cytokines (interleukin (IL)-6 and IL-8, tumour necrosis factor (TNF)-α and its receptors tumour necrosis factor receptor (TNFR)-55 and TNFR-75) and acute phase reactant proteins (C-reactive proteins, lipopolysaccharide-binding protein (LBP), FAS and FAS ligands) 1, 17, 18. Different studies have shown that systemic inflammation may trigger a catabolic/anabolic imbalance that results in skeletal muscle wasting and reduced muscle strength 15, 47. Indeed, inflammation can have a negative impact on muscle protein catabolism via different cytokine-mediated pathways, particularly TNF-α. First, inflammation increases the demand for amino acids to synthesise acute phase proteins in the liver and reduces muscle protein stores 48. Secondly, TNF-α activates the adenosine triphosphate (ATP)-ubiquitin-dependent proteolytic system, through which muscle proteins are degraded and repair systems are inhibited 49. Thirdly, TNF-α stimulates apoptosis via fragmentation of DNA and/or interaction with the TNF-α receptor present on muscle cells 50. Increased levels of plasma pro-inflammatory cytokines have also been reported to decrease anabolic factors by altering the bioavailability and biological effects of key hormones needed for protein synthesis, growth and maintenance of skeletal muscle such as testosterone, insulin growth factor-1 (IGF-1) and dehydroepiandrosterone (DHEA). It is further important to specify that TNF-α may also have a direct inhibitory effect on myofilaments and may alter muscle contractility, irrespective of changes in protein degradation or synthesis 51.
Although the involvement of systemic inflammation in the peripheral muscle wasting and weakness of COPD has never been directly demonstrated, many data in the literature strongly support a causal relationship. Indeed, peripheral muscle wasting and weakness have been convincingly associated with increased levels of serum acute phase reactant proteins (C-reactive proteins and LBP) and inflammatory cytokines (IL-8, and TNF-α receptors) in these patients 52–54. The levels of glutamic acid and glutamate in the vastus lateralis, as well as the sum of plasma amino acids (mainly alanine, glutamine and glutamic acid), have also been reported to be significantly decreased and inversely correlated with serum levels of LBP in COPD, suggesting that amino acids are indeed redirected from muscle to liver in these patients 55, 56. The levels of systemic testosterone, IGF-1 and DHEA have been further reported to be significantly reduced and inversely correlated with serum levels of IL-6 in COPD patients as compared with healthy subjects 57–58. In the same study, Debigare et al. 58 showed a progressive rise in the IL-6/DHEA ratio with worsening of peripheral muscle atrophy in these patients, which strongly suggested that systemic inflammation, especially catabolic/anabolic ratio disturbances, is at least partly implicated in the peripheral myopathy of COPD. Lastly, it has recently been shown that excessive apoptosis of peripheral skeletal muscles occurs in patients with COPD and weight loss 59. However, the role of increased levels of TNF-α in the pathogenesis of this apoptosis in COPD nevertheless needs to be elucidated.
Hypoxaemia
Several studies performed on healthy subjects exposed to high-altitude hypoxia over a few months have demonstrated the detrimental effects of hypoxaemia on specific skeletal muscle properties: functional (reduced strength and endurance) 60, 61; morphological (atrophy) 62; and metabolic (reduced Krebs cycle enzyme activity) 63. As patients with COPD may present either chronic moderate-to-severe hypoxaemia or repeated episodes of hypoxaemia due to exercise-induced desaturation or sleep apnoea, hypoxaemia has been investigated as a potential contributor to myopathy in these patients. The proportion of type I fibres within the peripheral muscles of COPD patients was reported to be significantly smaller in hypoxaemic as compared with nonhypoxaemic patients 64 and strongly correlated with their resting value of O2 tension in arterial blood (Pa,O2) 65. In agreement with this notion, Serres et al. 29 showed that the quadriceps endurance capacity was significantly correlated with the resting Pa,O2 in COPD patients. Lastly, oxygen supplementation improves aerobic metabolism and ATP production in hypoxaemic COPD patients 66, 67. Altogether, the literature data strongly indicate that hypoxaemia contributes to the peripheral myopathy of COPD. However, the molecular and cellular mechanisms underlying the deleterious effects of hypoxaemia on skeletal muscle still need to be elucidated.
Other mechanisms?
A regression analysis revealed that catabolism/anabolism imbalance explained 20% of the peripheral muscle atrophy observed in nonhypoxaemic COPD patients without long-term steroid treatment 58. It is possible that hypoxaemia 68 and corticosteroid use 69, in association with systemic inflammation, enhance peripheral muscle wasting and reduce muscle strength in COPD. However, many studies have reported a significant reduction in peripheral muscle strength 38, endurance 25, 35, proportion of type I fibres 41, expression of slow myosin heavy chain 9 and oxidative enzyme activity 70 in moderate-to-severe COPD patients who did not present chronic or intermittent hypoxaemia and had not been treated with steroids in the 6 months preceding the studies.
Altogether, the available data indicate that corticosteroid use, inflammation and hypoxaemia contribute to the peripheral myopathy of COPD, but it is unlikely that these are the only mechanisms involved. A quite recent notion is that muscle oxidative stress occurs and contributes to the peripheral muscle myopathy in these patients (fig. 3⇓). Indeed, as will be discussed further on, muscle oxidative stress, which is commonly observed in chronic inflammatory disease, is well known to induce deleterious effects on skeletal muscle cells.

Both muscle disuse and myopathy may contribute to peripheral muscle dysfunction in chronic obstructive pulmonary disease (COPD). In these patients, myopathy is likely to result from several interrelated pathophysiological mechanisms, possibly in association with corticosteroid use.
MUSCLE OXIDATIVE STRESS AND MUSCLE DYSFUNCTION IN COPD PATIENTS
The superoxide anion (O2·−) and nitric oxide (NO·) are the most relevant radicals in living organisms. The first is formed by the univalent reduction of triplet-state molecular oxygen, mainly mediated by enzymes such as the reduced nicotinamide adenine dinucleotide phosphate (NAD(P)H) oxidases and xanthine oxidase, or by redox-reactive compounds such as the semi-ubiquinone compound of the mitochondrial electron transport chain.
NO· is synthesised from the amino acid L-arginine by the NO synthases (NOS), which include constitutively expressed neuronal (nNOS) and endothelial (eNOS) isoforms and the inducible (iNOS) isoform. Both O2.− and NO· are readily converted by enzymes or nonenzymatic chemical reactions into other reactive oxygen or nitrogen species (ROS and RNS, respectively), which can in turn give rise to new radicals. Many researchers have demonstrated that moderate ROS and RNS generation serves important, distinct and specific physiological functions, whereas excessive generation can overwhelm antioxidant capacity and lead to oxidative stress and damage to cell components 71. Briefly, the reaction of RONS with fatty acyl moieties in membrane phospholipids causes a chain reaction of peroxidation, the so-called lipid peroxidation that leads to severe alterations in both structural and functional membrane properties. Furthermore, protein oxidation or nitration by RONS can modify the structure or chemical properties of the affected proteins, thereby leading to a decline in protein function or even complete protein unfolding. Finally, RONS can interact with DNA, causing DNA damage and subsequent alterations including deletions, mutations, DNA-protein cross-links, etc. 72–74. It is, however, important to note that ROS and RNS may selectively target molecular components, most likely depending on their nature, reactivity and site of production 71. Altogether oxidative stress-induced molecular damage has been shown to mediate morphological, metabolic and functional muscle abnormalities 71. This review will now focus on oxidative stress as a potential mechanism of myopathy in COPD.
Does oxidative stress occur in the peripheral muscles of COPD patients?
At rest
Over the past several years, specific needle biopsy techniques have been developed to enable researchers to determine whether oxidative damage occurs in the peripheral skeletal muscles of COPD patients. Levels of 4-hydroxyl-2-nonenal and lipofuscin, both products of membrane lipid peroxidation that accumulate in the cells in the vastus lateralis, were reported to be significantly higher in resting moderate-to-severe COPD patients than in healthy subjects 75, 76. Other studies assessed malondialdehyde (MDA), another marker of lipid peroxidation, and reported no significant difference between the two groups at rest 35, 77. These results are only apparently conflicting, since MDA is an acute marker that is rapidly eliminated by the cells. Similarly, the levels of protein tyrosine nitration were shown to be significantly greater in the resting leg muscles of COPD patients as compared with healthy subjects, whereas the levels of protein oxidation, detected as carbonyl groups (also an acute marker), were nearly the same 75. Taken together, these data indicate that both ROS and RNS selectively target molecular components in COPD and provide evidence of RONS-mediated cell damage, in particular enhanced lipid peroxidation and protein nitration, in the resting leg muscles of these patients. An obvious conclusion is that COPD peripheral muscle is exposed to repeated episodes of oxidative stress.
After exercise
Interest has recently focused on the effects of exercise on systemic oxidative stress in COPD patients 77–79. Among the studies, two have shown that strenuous incremental cycle exercise, and even light constant cycle exercise, induces systemic oxidative stress in patients with moderate-to-severe COPD, strongly suggesting that these patients are frequently exposed to oxidative damage as they perform their daily living activities 78, 79. However, because these studies focused on whole body exercise and did not identify the specific source of the exercise-induced stress, the question remains: did it originate from lung, peripheral muscles, heart, liver or other organs?
Testing the hypothesis that exercise-induced oxidative stress originates, at least in part, from exercising peripheral muscles requires the design of specific research protocols. In fact, highly localised limb muscle exercise is essential to reduce, as much as possible, the sources of oxidative stress that are intrinsic to whole body exercise. Thus, Serres et al. 7 developed a model, also used by several groups 25, 27, 35, of peripheral muscle exercise based on one-leg repeated knee extensions performed at an individualised constant workload until exhaustion. This localised limb exercise, which has been reported to induce only moderate cardiac and ventilatory responses 25, is a reliable clinical method to assess peripheral muscle endurance in patients with COPD.
Following this quadriceps exercise, plasma lipid peroxidation was significantly increased in the patients, whereas no changes occurred in the healthy age-matched sedentary subjects 14. The current authors hypothesised that contracting muscle was the source of the exercise-induced systemic oxidative stress in the patients, and this was confirmed by needle biopsies of the vastus lateralis that revealed significantly increased lipid peroxides and oxidised proteins in the patients but not in the healthy sedentary subjects 35.
Here, the current authors would like to remind readers of the recent data indicating an increased susceptibility to oxidative stress in association with other chronic diseases. Chronic skeletal muscle oxidative stress and more numerous systemic markers of oxidative damage after exercise have been reported in patients with chronic heart failure 80, 81, chronic uremia 82, 83 and diabetes 84–86 as compared with healthy subjects. This may suggest the existence of a common scenario in all these diseases.
Does oxidative stress contribute to the peripheral muscle dysfunction in COPD?
From a clinical perspective, it is obviously important to determine whether exercise-induced oxidative stress is implicated in the peripheral muscle dysfunction of patients with COPD. The current authors' group recently established an inverse relationship between the magnitude of exercise-induced muscle oxidative stress and quadriceps endurance in COPD patients, and suggested that cytotoxic oxidants have a role in the reduced quadriceps endurance capacity observed in these patients 35 (fig. 4⇓). The authors' group also reported that short-term, high-dose N-acetylcysteine (NAC, an antioxidant) treatment prevented exercise-induced oxidative stress and significantly improved COPD quadriceps endurance, which further supports the concept that muscle oxidative stress is at least partly implicated in the peripheral muscle dysfunction of these patients 87.

An inverse and significant correlation exists between quadriceps endurance time and the exercise-induced increase in muscle lipids peroxidation in patients with chronic obstructive pulmonary disease (r = -0,66; p<0.05). Reproduced from 35.
These results agree with both animal and human studies showing that intracellular oxidant concentrations modulate the contractile function of mammalian skeletal muscles in a dose-dependent manner 88–91. Reid et al. 90, for example, showed ∼15% increase in sustainable force during fatiguing exercise in the anterior tibialis of healthy subjects after they were intravenously pre-treated with NAC. This effect was evident after 3 min of repetitive contractions and persisted throughout the 30-min protocol. Furthermore, Travaline et al. 92 showed that pre-administration of intravenous NAC may attenuate the development of fatigue and significantly increase diaphragm endurance in healthy subjects breathing against high inspiratory loads. Taken together, these data provide strong evidence of oxidative stress as a mediator of reduced endurance capacity and acute fatigue in skeletal muscle.
How does muscle oxidative stress mediate muscle dysfunction?
The excessive ROS and/or NOS produced within muscle fibre mainly target mitochondria and myofilaments, which can lead to apoptosis, mitochondrial respiratory chain dysfunction, and/or an alteration in myofilament contractile properties. Thus, these are three molecular mechanisms that may underlie oxidative stress-induced peripheral muscle dysfunction (fig. 5⇓).

Oxidative stress within muscle fibre can lead to apoptosis, mitochondrial respiratory chain dysfunction and/or alteration in myofilament contractile properties resulting in skeletal muscle dysfunction. ATP: adenosine triphosphate.
Mitochondria
Muscle apoptosis
A large number of studies have reported that massive formation of RNS and/or ROS is closely associated with increased apoptotic processes in the myocytes of animals and humans 93. In such multinucleated cells, this may lead to atrophy and reduced muscle strength 94.
Other data suggest that oxidative stress-induced apoptosis is mainly mediated via ROS and RNS-induced release of caspase-activating proteins such as cytochrome c and apoptotic-inducible factor. High levels of ROS and RNS have been reported to change cellular redox potentials, deplete reduced glutathione, and decrease ATP levels and such reducing equivalents as reduced nicotinamide adenine dinucleotide (NADH) and NADPH. These changes can facilitate the formation of permeability transition pores, which leads to the release of cytochrome c 95. The reaction of ROS and RNS with membrane phospholipids can also increase membrane permeability, so that caspase-activating proteins are released from mitochondria into the cytosol. Furthermore, excessive concentrations of ROS or RNS have been shown to significantly reduce the anti-apoptotic/pro-apoptotic protein ratio 96, thereby promoting the mitochondrial release of pro-apoptotic factors.
To the authors' best knowledge, the possibility that apoptosis presents a fibre-type specificity has never been investigated. However, type I fibres, which are predominantly oxidative, may be overwhelmed by the increased amount of mitochondrial RONS produced during exercise and/or under pathophysiological conditions, thereby enhancing the susceptibility to apoptosis. In this regard, it is interesting to note that Allaire et al. 76 determined that lipofuscin accumulation, as a marker for oxidative damage, was significantly greater in fibres with high (type I) as compared with low (type II) oxidative potential in the vastus lateralis of patients with COPD. Furthermore, as mentioned earlier, it was recently shown that apoptosis does not occur in the vastus lateralis of normal weight-stable COPD patients 59, although it occurs in more severe and underweight patients 97. It should be noted that the relationship between oxidative stress, apoptosis and the proportion of type I fibres in COPD peripheral muscles has never been clearly established. Due to of the drastic reduction of oxidative fibres observed in severe COPD, the current authors believe that this is an obvious perspective for research.
Altered oxidative capacity
Oxidative stress causes mitochondrial respiratory chain dysfunction and alters oxidative phosphorylation 98, 99, which leads to reduced ATP concentrations and greater intracellular acidosis. Studies on animal models have indicated that ROS and RNS may directly react with lipids and/or proteins to significantly alter the activity of the mitochondrial respiratory chain complex (I, II, IV) 100, 101 and reduce the activity of mitochondria 102 or Krebs cycle oxidative enzymes 103–105. Interestingly, antioxidant treatment increases the activity of the oxidative enzymes and respiratory complex in mice exposed to oxidative stress, thus supporting the cause and effect relationship between oxidative stress and altered oxidative phosphorylation 106. Moreover, increased mitochondrial permeability due to membrane lipid peroxidation can trigger ion leakage, i.e. a reduced proton gradient over the inner membrane of the mitochondrion and highly uncoupled oxidative phosphorylation. This results in respiratory chain dysfunction 107. Lastly, it has been shown that oxidative/nitrosative modification and mutation of mitochondrial DNA can cause defects in the respiratory chain due to impairment of the gene expression and the structure of the respiratory chain polypeptides that are encoded by the mitochondrial genome 108. Significantly reduced oxidative enzyme activity during exercise, associated with premature lactic acidosis, has been clearly demonstrated in the lower limb muscles of patients with COPD 70 and is generally attributed to the reduced proportion of oxidative fibres observed in these patients 64. These findings emphasise the need to understand the mechanisms underlying the reduced proportion of type I fibre in COPD muscle. However, it cannot be ruled out that the reduced oxidative capacity and the concomitant increase in lactic acid production may be independently attributed, at least in part, to muscle oxidative stress.
This research perspective requires an experimental design that can determine whether short-term antioxidant treatment improves oxidative phosphorylation and reduces exercise-induced intracellular acidosis, without concomitant changes in fibre-type distribution, in patients with COPD.
Myofilaments
In vitro studies on animal models have shown that, in a concentration- and time-dependent manner, intracellular ROS and RNS significantly reduce the activities of the Na+/K+ pump, which is mainly implicated in membrane depolarisation and the transmission of action potentials throughout the cell 109. Moreover, high intracellular concentrations of RONS have been shown to reduce the activities of the specific pumps responsible for the entry of Ca2+ from the cytosol back into the sarcoplasmic reticulum 110. The ryanodine receptor channels (RyR) (responsible for the release of Ca2+ from sarcoplasmic reticulum) are also known to be redox responsive because they contain a large number of sulhydryls whose oxidation and/or nitration modulate the function. However, it is important to note that ROS and NO appear to have respectively deleterious and regulatory effects on RyR activity. Indeed, ROS-induced oxidative modification may lead to intersubunit disulphide bond formation and cross-links of the RyR subunits that result in irreversible activation of RyR and subsequent sarcoplasmic reticulum (SR) leak and futile Ca2+ cycling. In contrast, NO-induced covalent modifications such as poly-S-nitrosylation have been reported to reversibly activate the channel, promote SR cycling, block intersubunit cross-links and prevent irreversible activation of the skeletal RyR channel by oxidants. Whether these RyR channels are modulated reversibly or irreversibly is determined by the relative balance between nitrosative and oxidative stress 111, 112. Taken together, the data indicate that oxidative stress significantly disrupts Ca2+ homeostasis in favour of intracellular cytosolic Ca2+ accumulation. These modifications may lead to a reduced rate of myofilament relaxation, thus limiting the amount of hydrolysed ATP and the force generated by the actin and myosin interactions, as well as the velocity of shortening.
An important point is that oxidative stress has been reported to significantly modify the conformation and increase the degradation of actin and myosin in rat skeletal muscle 113. These alterations may decrease the troponic-C affinity for calcium, so that a given increase in calcium in the vicinity of the troponin complex will have less of an activating effect on muscle contraction. A recent study reported a significant increase in muscle content of oxidised proteins after exercise in the vastus lateralis of COPD patients compared with healthy subjects, but it failed to distinguish what type of protein was oxidised 35. It would also be of interest to determine whether exercise-induced RONS generation preferentially targets actin and/or myosin in the peripheral muscle of these patients. If so, is there a causal relationship between contractile protein oxidation and the occurrence of early contractile fatigue during exercise in COPD?
MOLECULAR MECHANISMS UNDERLYING MUSCLE OXIDATIVE STRESS IN COPD
From a therapeutic perspective, the mechanisms underlying muscle oxidative stress in COPD need to be elucidated. Increased muscle oxidant production and/or reduced muscle antioxidant capacity can be hypothesised.
An increase in muscle ROS and/or NOS production
In the pathophysiological context of COPD, the principal sources of excessive RONS generation in the peripheral muscles would likely be the mitochondrial respiratory chain, xanthine oxidase, macrophage and neutrophil NADPH oxidase, or NOS.
Mitochondrial respiratory chain
Mitochondria are one of the main sources of ROS production because 2–5% of the oxygen consumed is not fully reduced in the electron transport chain and, thus, may leak away as superoxide radicals 114. Several studies have shown that ROS production is a function of the mitochondrial membrane potential. Indeed, ROS production is increased during state 4 respiration, when the inner membrane mitochondrial potential is at its peak (fig. 6⇓). An increase in mitochondrial membrane potential slows electron transport through the respiratory chain, resulting in an increase in the ubiquinone free-radical half-life. As a result, electrons have an increased probability of interacting with oxygen to form ROS 115. It has recently been shown that mitochondria possess a mechanism called “mild” uncoupling, which promotes proton leakage and prevents large increases in the proton electrochemical gradient, thereby reducing ROS production by the respiratory chain 116, 117. This is important in skeletal muscle, given that it accounts for an important fraction of the total oxygen consumption at rest and that this fraction is even higher when skeletal muscle switches to maximal activity. This “mild” uncoupling has been shown to be mediated by a specific uncoupling protein of the inner mitochondrial membrane called uncoupling protein 3 (UCP3). Mice lacking UCP3 were found to produce more ROS than wild-type mice 118, thus indicating that one of the functions of UCP3 is to prevent excessive oxidative stress in skeletal muscle (fig. 6⇓).

Mitochondrial reactive oxygen species (ROS) production is dependent on the membrane potential. A high mitochondrial membrane potential leads to inhibition of electron transport by the respiratory chain, resulting in increased half-life of intermediates, capable of reducing O2 to superoxide anion (O2·−), such as the semiquinone radical (Q·). Uncoupling proteins (UCP) have been reported to decrease the membrane potential, particularly under conditions of low adenosine diphosphate (ADP) availability, thus reducing mitochondrial ROS generation. ▵μH+ : electrochemical gradient; ATP: adenosine triphosphate; Cyt c: cytochrome c; NAD+: oxidised nicotinamide adenine dinucleotide; NADH: reduced nicotinamide adenine dinucleotide.
In regard to this, the activity of cytochrome c oxidase (COX), which is the final mitochondrial enzyme of the respiratory chain (complex IV), was noted to be significantly increased in the vastus lateralis of hypoxaemic COPD patients as compared with healthy subjects 119. In addition, a severe reduction in the expression of UCP3 was found in the vastus lateralis of these patients as compared with healthy subjects 120, 121. Such increased COX and reduced UCP3 activities may abnormally increase the mitochondrial membrane potential and slow electron transport through the respiratory chain, thereby promoting ROS generation in COPD peripheral muscles. Studies are still needed, however, to determine whether the electron transport chain is indeed altered and, if so, whether this contributes to the genesis of muscle oxidative stress in COPD patients.
Xanthine oxidase
Xanthine oxidase, in the muscle or in its capillary endothelium, is also an important source of oxidant production during exercise when in the presence of increased intracellular calcium concentration and ATP breakdown products, including hypoxanthine and its precursor inosine monophosphate (IMP). Heunks et al. 78 demonstrated that exercise-induced systemic oxidative stress during cycling exercise in COPD patients can be prevented by pre-treating the patients with allopurinol, a potent xanthine oxidase inhibitor. The potential role of this enzyme in generating oxidative stress was further substantiated by Pouw et al. 122, who reported elevated IMP levels in the quadriceps of COPD patients at rest compared with healthy subjects. Furthermore, Vina et al. 79 found that oxygen supplementation attenuates cycle exercise-induced systemic oxidative stress in these patients. This result supports the concept of xanthine oxidase as a source of free radical generation during exercise in COPD, because oxygen supplementation is likely to reduce metabolic stress, thereby limiting ATP degradation and IMP accumulation in exercising muscles. However, these studies focused on whole body exercise and did not localise the activated xanthine oxidase: hepatocytes, macrophages, vascular smooth muscle, cardiac muscles, etc. 123. Therefore, further studies are required to specifically isolate muscle xanthine oxidase as the source of excessive free radical generation in the exercising muscle of patients with COPD.
NOS
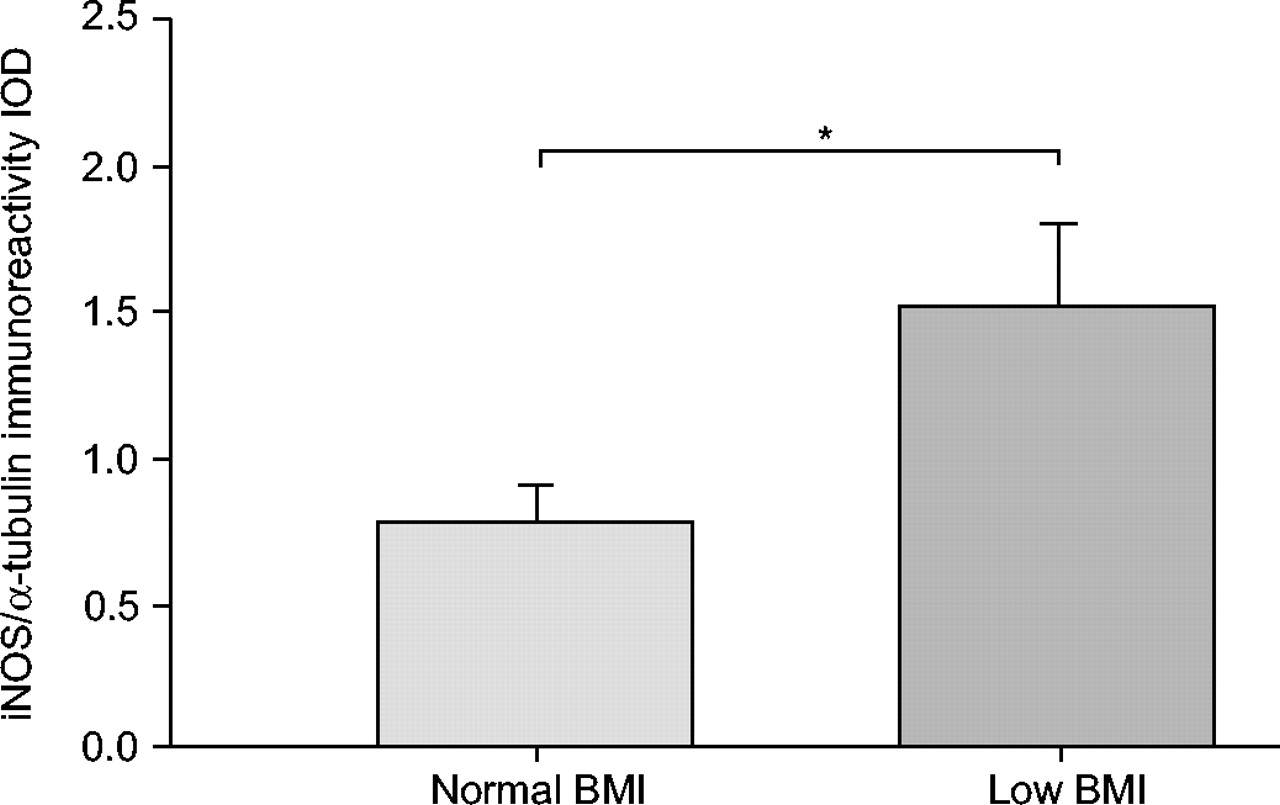
Over the past few years, there has been increasing evidence that muscle redox status is strongly influenced by NO, as well as by ROS 124. Indeed, an increase in NO production mediated by NOS in combination with excessive superoxide anion generation can lead to the formation of the highly reactive oxidant, peroxinitrite, resulting in nitrosative stress and inactivation of the enzymes involved in the defences against oxidative stress 88. Barreiro et al. 75 recently investigated NOS expression in the vastus lateralis of patients with moderate COPD in comparison with healthy subjects and reported that mean iNOS and eNOS expression did not differ between groups. Although no difference in muscle nNOS was detected between controls and the entire COPD group, those patients with forced expiratory volume in one second (FEV1 <50% of predicted showed a strong tendency toward higher expression of nNOS as compared with controls, which inversely correlated with the relative FEV1 values. Furthermore, the mean values of total muscle 3-nitrotyrosine level, as an index of nitrosative stress (protein tyrosine nitration), were ∼200% higher in the patients with COPD than in the healthy subjects and significantly correlated with nNOS expression. No such correlation was detected between muscle tyrosine nitration and eNOS expression, suggesting that enhanced nNOS expression could be one of the mechanisms involved in the oxidative damage observed in the peripheral muscles of patients with COPD. It is further important to note that Agusti et al. 125 recently showed increased activation of nuclear transcription factor (NF-κβ) and up-regulation of iNOS in the vastus lateralis of patients with severe COPD and low body mass index (BMI) as compared with patients with moderate-to-severe COPD and normal BMI. These results suggest that iNOS activation in the peripheral muscles of COPD is strongly dependent on the degree of disease severity, at least as expressed by FEV1 and BMI (fig. 7⇓).

Mean±se inducible nitric oxide synthase (iNOS) expression (normalised by Ñ-tubulin content) within vastus lateralis of moderate-to-severe chronic obstructive pulmonary disease patients with normal body mass index (BMI; 27.1±1 kg·m−2) as compared with low BMI (18.5±1 kg·m−2). Reproduced from 125. *: p<0.05.
NADPH oxidase
Activated macrophages and neutrophils can produce large amounts of superoxide and its derivatives via the phagocytic isoform, NADPH oxidase. The activation of NADPH oxidase is mainly induced by cytokines such as interferon-γ, IL-1β, IL-6 and IL-8 and TNF-α 126. One might expect that circulating inflammatory cells and cytokines would infiltrate the leg muscles of COPD patients, similar to bronchial inflammatory cell infiltration, thereby increasing ROS production. Yet Gosker et al. 97 did not find abnormal numbers of macrophages and leukocytes in the vastus lateralis of moderate COPD as compared with healthy subjects at rest. Furthermore, the leg muscles of moderate patients did not express significantly more iNOS protein than the muscles of healthy subjects, thus suggesting that the levels of inflammatory cells and cytokines inside these muscles were not abnormally elevated 75. However, Rabinovich et al. 127 recently showed that TNF-α mRNA expression was significantly greater in the vastus lateralis of moderate-to-severe COPD patients than of healthy subjects. The authors also reported a significant rise in plasma TNF-α after a moderate-intensity constant work-rate cycling exercise in the patients, which did not occur in the healthy controls. Once again, these results were obtained after whole body exercise and, thus, the working muscles could not be isolated as the source of cytokine release in the patients. However, it has recently been reported that localised quadriceps exercise-induced systemic oxidative stress occurs in patients with COPD, without a concomitant increase in blood levels of inflammatory cytokines 128. Altogether, these data seem to indicate that intracellular inflammation is an unlikely occurrence in the peripheral muscles of moderate COPD and, thus, activated NADPH oxidase is not the primary mechanism involved in the pathogenesis of muscle oxidative stress in these patients. However, as will be discussed further on, the possibility cannot be excluded that the marked systemic inflammation observed at rest in these patients may act directly or indirectly to trigger oxidative stress within contracting peripheral muscles.
Furthermore, it is important to bear in mind that apoptosis and upregulation of NF-κβ and iNOS have been noted in the peripheral muscles of severe COPD patients with low BMI 59, 125, which suggests that an inflammatory process and increased NADPH oxidase activity are more likely to occur in this subgroup of patients. This hypothesis, however, needs to be confirmed.
A reduction in antioxidant capacity
Evidence is growing that oxidant production may be used by cells to signal an adaptive enzymatic antioxidant response to prevent oxidative stress 129. The expression of an important mitochondrial enzyme responsible for the dismutation of superoxide anion, mitochondrial superoxide dismutase (Mn-SOD), was significantly elevated (≈200%) in the resting leg muscles of COPD compared with healthy controls 75. Furthermore, the current authors' group reported that glutathion peroxidase (GPX) activity was similar in the resting vastus lateralis of COPD and healthy subjects 35, despite the acknowledged histological changes in the COPD muscle in favour of type II fibres 36. This result is in fact somewhat surprising because type II muscle fibres have lower levels of antioxidant enzyme activity than type I 130. Thus, increased Mn-SOD and normal (instead of reduced) resting GPX activities may reflect a mechanism of adaptation to the increased free radical generation inside COPD skeletal muscle. This notion is supported by the depletion of muscle glutathion (GSH), a substrate required for GPX-catalysed reactions, reported in the resting vastus lateralis of these patients 131.
Moreover, a one-leg endurance exercise was reported to increase GPX activity in the vastus lateralis of healthy sedentary subjects, while no changes occurred in patients with COPD 35. This alteration in the GPX response of the patients may be attributed to muscle GSH depletion and may contribute to the greater susceptibility to exercise-induced muscle oxidative stress observed in COPD. This hypothesis fits well with the observation that supplementation with NAC, a precursor for GSH synthesis, significantly reduces quadriceps exercise-induced oxidative stress in patients with COPD 87. It is further important to note that the plasma reduction of nonenzymatic antioxidant levels observed in severe COPD 14, 132 can add to the altered muscle enzymatic antioxidant activity to increase the risk for exercise-induced oxidative stress in these patients (table 1⇓).
Antioxidant status in plasma and vastus lateralis of patients with severe chronic obstructive pulmonary disease(COPD) and normal body mass index (BMI) as compared with healthy age-matched subjects at rest and after exercise
HOW MIGHT LUNG DISEASE TRIGGER PERIPHERAL MUSCLE OXIDATIVE STRESS?
Several experimental studies in animals have evidenced inflammation and hypoxaemia as enhancers of RONS production in skeletal muscles 133, 134. In patients with COPD, systemic inflammation was strongly associated with an enhanced chronic and systemic oxidant burden 17. Moreover, it is well known that hypoxaemia contributes to the development of systemic inflammation in these patients 135. The current objective here is to discuss how inflammation and hypoxaemia, whether associated or not, might trigger oxidative stress in the peripheral muscles of patients with COPD.
Inflammation
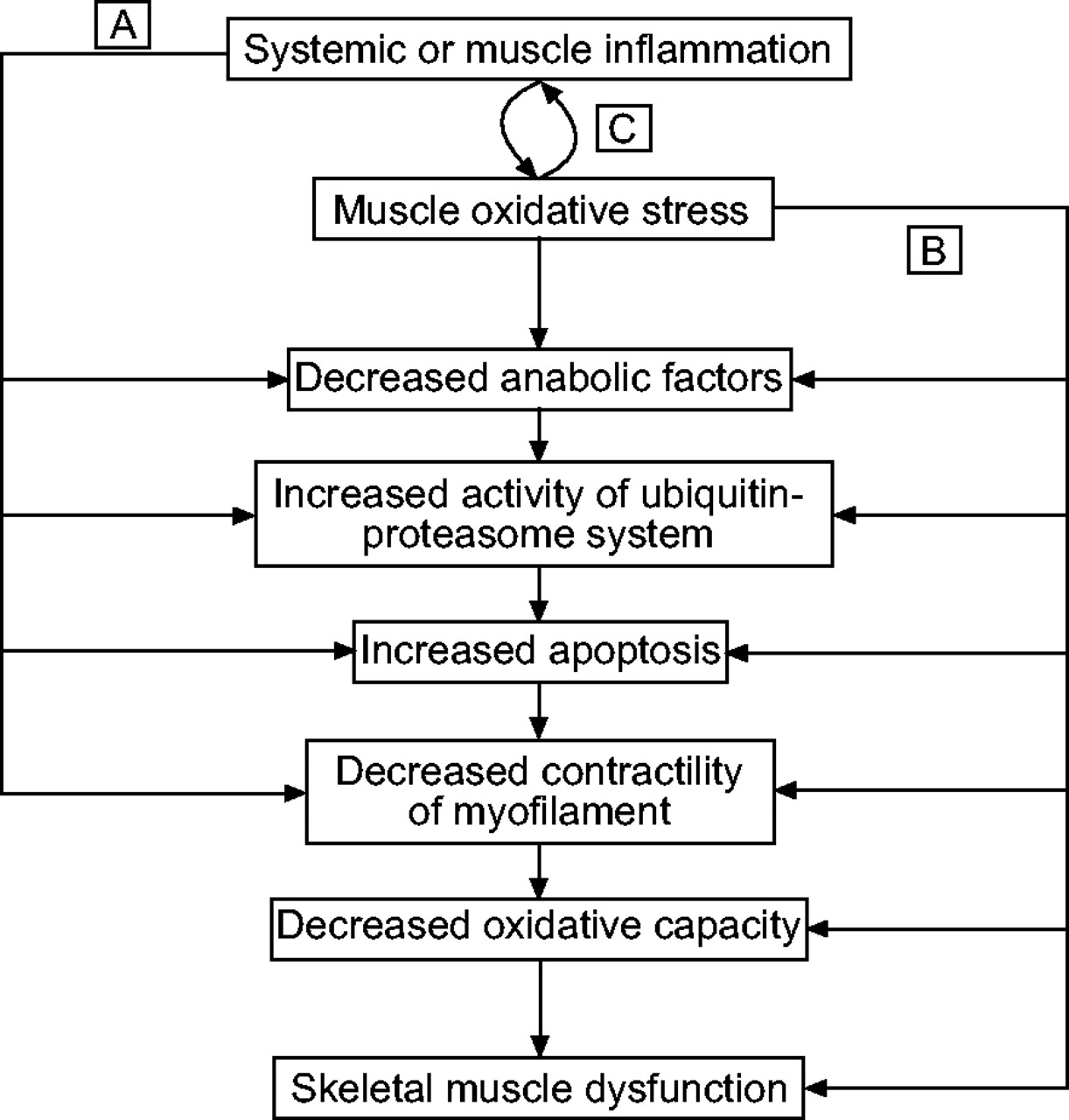
A number of studies have shown evidence that oxidants influence inflammation and that inflammation plays a role in inducing oxidative stress 17. The inflammatory response can be initiated and mediated by increased levels of ROS, which activate the transcription of pro-inflammatory cytokine and chemokine genes, upregulate adhesion molecules and increase the release of pro-inflammatory mediators. Conversely, several studies have indicated that inflammatory cytokines are capable of stimulating xanthine oxidase, NOS and NADPH oxidase, thereby increasing RONS generation and the risk of oxidative stress in various cells and tissues. In patients with COPD, systemic inflammation was shown to be strongly associated with increased ROS release by stimulated phagocytes and reduced plasma antioxidant capacity 136. A reduced basal antioxidant status due to systemic inflammation associated with a physiological increase in RONS production during exercise is able to act as a trigger for muscle oxidative stress in these patients (fig. 8⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
a) and b) Inflammation and oxidative stress can independently trigger muscle dysfunction by acting on mitochondria and myofilament properties. c) Inflammation and oxidative stress are interrelated mechanisms that would create a closed loop of persistence and amplification of the skeletal muscle abnormalities in patients with chronic obstructive pulmonary disease.
Can systemic inflammation directly promote excessive RONS generation in peripheral muscles? The answer is yes. A rise in circulating TNF-α levels leads to increased ligand binding to TNF-α receptors on the skeletal muscle whose activation stimulates intracellular ROS production 137. Yet it is important to bear in mind that the exact role of systemic inflammation in the pathogenesis of muscle oxidative stress in patients with COPD has never been fully elucidated. If this link can be clearly demonstrated, it will imply that muscle oxidative stress and the subsequent oxidative damage are considerably increased during acute episodes of exacerbation.
Hypoxaemia
Several in vitro and in vivo studies have reported that prolonged or acute periods of hypoxia increase RONS formation and promote oxidative stress in different tissues 138. Although not fully understood, this phenomenon seems likely to be mediated by hypoxia-induced upregulation in NOS and xanthine oxidase activities, as well as respiratory chain dysfunction 139–141. In patients with COPD, hypoxaemia is unlikely to be the only mechanism of muscle oxidative stress, since a significant increase in the markers of oxidative damage has been reported at rest and after localised exercise in patients with normal resting arterial oxygen tension (Pa,O2) and no exercise-induced oxygen desaturation 35, 76. However, recent data indicate that hypoxaemia can act in concert with other mechanisms to markedly increase the susceptibility to muscle oxidative stress in these patients. The current authors' group, for example, demonstrated that oxidative damage at rest and after localised exercise was significantly greater in hypoxaemic (Pa,O2 <9.3 kPa (<70 mmHg)) as compared with nonhypoxaemic patients 142.
Lastly, it is important to recall that hypoxaemia, inflammation and oxidative stress are strongly associated. In vitro studies have shown that hypoxia markedly induces the expression of inflammatory cytokines via transcriptional mechanisms 143, and Pa,O2 levels are inversely correlated with the TNF-α system (levels in circulating TNF-α and its receptors), in patients with COPD 135. As both hypoxaemia and inflammation can increase RONS production, it is difficult to determine whether they are interrelated or independent mechanisms in the pathogenesis of muscle oxidative stress in these patients. Yet an understanding of the relationship between the pathophysiology of COPD and muscle oxidative stress is crucial for the development of therapeutic strategies for different degrees of disease severity.
THERAPEUTIC PERSPECTIVES
The current data indicate that both physical training and antioxidant supplementation, perhaps in combination, can be useful to prevent exercise-induced muscle oxidative stress and improve muscle function.
Physical training
Many studies on animal models have shown that an exercise-induced increase in lipid peroxidation and acute fatigue in respiratory muscle are both significantly reduced after short-term endurance exercise training in rats 144. Physical training has been further reported to prevent exercise-induced systemic oxidative stress and to increase exercise tolerance in patients with chronic heart failure 81, 145 and healthy young and elderly subjects 146, 147. How can physical training prevent exercise-induced oxidative stress? This benefit is attributable to training-induced reduction in RONS generation and/or an increase in enzymatic antioxidant capacity. Indeed, physical training has been reported to improve mitochondrial membrane potential, respiratory chain function and oxidative metabolism, thereby reducing RONS generation by activated xanthine oxidase or altered mitochondria within contracting muscles 148, 149. Miyazaki et al. 147 reported that the magnitude of an exercise-induced increase in free radical generation in neutrophils was significantly reduced after 12 weeks of strenuous endurance training in healthy subjects. Moreover, physical training-induced muscle changes in thiol redox status can influence the intracellular transduction processes that trigger enzymatic antioxidant expression and activity. In this regard, evidence is growing that exercise training results in elevated activities in the enzymatic antioxidant systems of animal skeletal muscle 150, 151, whereas the results are less clear-cut in humans. One study showed that a 6- or 8-week programme of endurance training enhanced muscle oxidative power and the flux through the citric acid cycle in human skeletal muscle, without altering muscle antioxidative enzyme activities 152. In contrast, Ennezat et al. 153 reported that the transcript levels for SOD and GPX were significantly increased after 12 weeks of endurance training in patients with chronic heart failure. An examination of the literature suggests that the discrepancy between these results could be explained by differences in training modality, as well as in the populations studied 154.
Antioxidant supplementation
Nonenzymatic antioxidants such as vitamins A, E, C, selenium, thiols, glutathione, ubiquinone, flavonoids and others can interact with enzymatic antioxidants to reduce the risk of oxidative stress 150. Nonenzymatic antioxidant deficiency is known to enhance tissue lipid peroxidation, increase both skeletal muscle susceptibility to damage and the fragility of lysosomal membranes, compromise the respiratory control ratio in skeletal muscle, increase endoplasmic and sarcoplasmic reticulum lesions, and decrease exercise performance in animals 155. Interestingly, antioxidant supplementation has been reported to prevent exercise-induced muscle oxidative stress and increase muscle function in humans or animals with antioxidant deficiency 156–158. However, such benefits are strongly dependent on the type of association between antioxidants, as well as the dose and duration of the prescribed supplementation. In contrast, antioxidant supplementation of any type has no functional effect in subjects without antioxidant deficiency 159, 160.
Physical training combined with antioxidant supplementation
As physical training is based on repeated bouts of aerobic endurance exercise with consequent increases in oxygen utilisation, systemic and muscular RONS generation is physiologically increased. Although physical training has been reported to increase enzymatic antioxidant activity, it has also been shown to significantly reduce the systemic or tissue levels of nonenzymatic antioxidants 161. In line with this finding, a recent study reported that over-training significantly increased plasma GPX activity, but reduced nonenzymatic antioxidant capacity and promoted exercise-induced oxidative stress in triathletes 162. Thus, it is important to note that physical training may have no effect on oxidative stress, it may protect against it, or it may have an adverse effect, depending on its frequency and/or intensity; training that is too low may be insufficient to provide significant benefits, while training that is too high can be deleterious. This argues in favour of the “individualised” prescription of training and points to an interesting clinical question: how can we obtain a physical training-induced increase in enzymatic antioxidant capacity, without a concomitant decrease in nonenzymatic capacity? One of the possibilities is to associate physical training with antioxidant supplementation. Palazzetti et al. 163 reported that physical over-training-induced nonenzymatic antioxidant downregulation and increased susceptibility to oxidative damage can be avoided by antioxidant supplementation during the training period.
Perspectives for improving COPD peripheral muscle dysfunction
Information is currently lacking about the effects of physical training on the muscle oxidative stress in patients with COPD. To the current authors' knowledge, only Rabinovich et al. 77 evaluated the effects of exercise training (5 days·week−1 for 8 weeks) on limb muscle redox status in patients with COPD as compared with healthy subjects. Although the healthy control subjects markedly increased muscle-reduced GSH levels after training (100%), the GSH homeostasis in patient skeletal muscle did not undergo significant change except for the oxidised glutathion (GSSG) increase detected after the training period. In their study, the training sessions were split into small blocks of 2–5 min of high-intensity continuous cycling (at ∼90% of the peak work rate at the end of the training programme) for an effective period of ≥30 min. As mentioned previously, the training intensity chosen for this study may have been too high for COPD patients and, thus, could have generated oxidative stress at each training session, which would explain the increased GSSG. Further studies are needed to determine whether a lower intensity of exercise training is better adapted to prevent exercise-induced muscle oxidative stress and improve muscle function in patients with COPD. Finally, because plasma and muscle antioxidants are already reduced at rest in severe COPD 14, 136, the potential benefits of physical training associated with antioxidant supplementation in these patients should be considered.
CONCLUSION
After revisiting the key definitions and comparing the data obtained in chronic obstructive pulmonary disease patients and appropriately matched long-term sedentary subjects, this review presents evidence that the peripheral muscle dysfunction of chronic obstructive pulmonary disease is due not only to disuse, but also to a form of myopathy. This new concept implies pathophysiological mechanisms, whose identification should help in developing therapeutic strategies to improve muscle function, exercise intolerance and quality of life in these patients. Recent data indicate that oxidative stress occurs in chronic obstructive pulmonary disease peripheral muscle and that it contributes, at least in part, to the peripheral muscle dysfunction of these patients. Further studies are needed to elucidate the molecular mechanisms underlying the oxidative stress in chronic obstructive pulmonary disease muscle, to determine its relationship with systemic inflammation and/or hypoxaemia, and to develop appropriate therapeutic approaches to prevent it. From this perspective, physical training, perhaps in association with antioxidant supplementation, may be useful. This possibility, as well as the best training modality for these patients, needs to be investigated.
- Received December 8, 2004.
- Accepted June 6, 2005.
- © ERS Journals Ltd
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵










Jump To
- Article
- Abstract
- GENERAL DEFINITIONS
- DISUSE AND/OR A FORM OF MYOPATHY IN COPD
- POTENTIAL FACTORS OF MYOPATHY IN COPD
- MUSCLE OXIDATIVE STRESS AND MUSCLE DYSFUNCTION IN COPD PATIENTS
- MOLECULAR MECHANISMS UNDERLYING MUSCLE OXIDATIVE STRESS IN COPD
- HOW MIGHT LUNG DISEASE TRIGGER PERIPHERAL MUSCLE OXIDATIVE STRESS?
- THERAPEUTIC PERSPECTIVES
- CONCLUSION
- References
- Figures & Data
- Info & Metrics