





Abstract
This study was designed to investigate the possible effect of injurious mechanical ventilation on peripheral immune function of healthy rats. Three ventilation strategies were compared: 1) low peak inspiratory pressure (PIP)/positive end-expiratory pressure (PEEP); 2) high PIP/PEEP; and 3) high PIP/zero PEEP (ZEEP). As a reference group, healthy, nonventilated, sham-operated, anaesthetised rats were used.
After 4 h, rats were sacrificed and macrophage inflammatory protein (MIP)‐2 levels in lung and plasma were determined. Peripheral immune function was determined by measurement of splenic natural killer (NK) activity, mitogen-induced splenocyte proliferation and in vitro cytokine production. All immune measurements in the low PIP/PEEP group did not differ from the immune measurements in the reference group. High PIP strategies, irrespective of applied PEEP, enhanced MIP‐2 levels in lung and plasma.
NK cell activity, mitogen-induced splenocyte proliferation and MIP‐2 and interleukin (IL)‐10 production significantly decreased after high PIP/PEEP ventilation. In the high PIP/ZEEP‐ventilated group, the decrease in splenocyte proliferation, MIP‐2 and IL‐10 production and NK cell activity was more pronounced and interferon‐γ production was also significantly lower than in the low PIP/PEEP group.
These data show that high positive inspiratory pressure ventilation induces an inflammatory response in the lung, whereas at the same time the peripheral immune response is downregulated. Ventilator-induced peripheral immune suppression may contribute to poor outcome in acute respiratory distress syndrome patients.
- cytokine production
- mechanical ventilation
- mitogen-induced splenocyte proliferation
- natural killer cell activity
- rat
- ventilator-induced lung injury
This study was financially supported by the “Catharȳne Stichting Utrecht”.
Ventilator-induced lung injury (VILI) is characterised by increased alveolar-capillary membrane permeability, protein leakage, production of inflammatory mediators and ultimately, impaired gas exchange 1. The degree of injury depends on the strategy and mode of ventilation. Positive end-expiratory pressure (PEEP) may prevent lung injury and inflammation, whereas strategies with high peak pressures, high tidal volumes and low PEEP enhance VILI 2–4.
Recently, it has been shown in vivo that the inflammatory mediator macrophage inflammatory protein (MIP)‐2, a very potent neutrophil chemoattractant, is a critical mediator in the development of VILI. Strategies with high peak pressures and no PEEP induced MIP‐2 expression in the lung 5. Moreover, in MIP‐2 receptor (CXCR2)-/- animals, the development of VILI is profoundly reduced 5. Previous studies in rats, which had been pretreated with HCl or lipopolysaccharide intratracheally, have shown that injurious ventilation strategies do not only enhance inflammation in the lung, but also increase the level of tumour necrosis factor (TNF)‐α and MIP‐2 in the systemic circulation 6, 7.
Systemic effects of mechanical ventilation (MV), including increased proinflammatory cytokine levels, have been suggested to contribute to the development of multiple system organ failure (MSOF) 8, 9. In acute respiratory distress syndrome (ARDS) patients, not only intra-alveolar levels but also plasma levels of the inflammatory mediators TNF‐α, interleukin (IL)‐6 and IL‐1β were higher after conventional ventilation than after lung-protective ventilation 10. Moreover, changes in cytokine levels and MSOF scores were positively correlated in ARDS patients 11.
Many critically ill, ventilated intensive care unit (ICU) patients suffer from unexplained immunosuppression and an associated increased risk for infections and MSOF 12–16. The immunosuppression in these patients is reflected by reduced peripheral blood lymphocyte proliferation and cytokine production 12, 13. There are indications that MV as such may contribute to suppression of the peripheral immune response. The authors previously described changes in the peripheral immune response in children without lung pathology after 2 h of MV and anaesthesia. In these children, both peripheral blood leukocyte production of proinflammatory cytokines and natural killer (NK) activity were decreased 17. However, in this human study, a possible effect of anaesthesia on the immune response could not be ruled out.
The present study was designed to compare the effects of different MV strategies on the peripheral immune system in rats without pre-existing lung pathology. Therefore, three different ventilation strategies were applied in the healthy rat: one with high peak pressures and PEEP, one with high peak pressure without PEEP and one protective strategy with low peak pressures and PEEP. Healthy, nonventilated, sham-operated, anaesthetised rats were included as a reference group. As a marker for inflammatory activity, MIP‐2 was measured in lung and plasma. To study peripheral immune function, i.e. the activity of the immune system outside the lung, mitogen-induced splenocyte proliferation, production of proinflammatory cytokines (interferon (IFN)‐γ and MIP‐2) and the anti-inflammatory cytokine IL‐10 were measured, as well as the NK cell activity of rat splenocytes.
Methods
Experimental protocol
A total of 28 Sprague-Dawley rats (250–300 g) were obtained from Harlan CPB (Zeist, the Netherlands). All animals were handled in accordance with European Community Guidelines and the study was approved by the experimental animal committee of the Erasmus MC‐Faculty (Rotterdam, the Netherlands). The rats were tracheotomised under inhalation anaesthesia (65% nitrous oxide/33% oxygen/2% isoflurane (isoflurane from Pharmachemie BV, Haarlem, the Netherlands). A sterile metal cannula was inserted into the trachea and a polyethylene catheter was inserted into the carotid artery to draw arterial blood gases and monitor blood pressures. Subsequently, inhalation anaesthesia was discontinued and anaesthesia was continued with 60 mg·kg−1·h−1 pentobarbital sodium (Nembutal; Algin BV, Maassluis, the Netherlands) given intraperitoneally. Muscle relaxation was attained with 2 mg·kg−1·h−1 pancuronium bromide (Pavulon; Organon Technika, Boxtel, the Netherlands) intramuscularly.
The rats were connected to a ventilator (Servo Ventilator 900 C; Siemens-Elema, Solna, Sweden) and ventilated for 4 h in a pressure-controlled, time-cycled mode, at an inspiratory oxygen fraction (FI,O2) concentration of 1.0, inspiratory:expiratory ratio of 1:2 and a frequency of 20–30·min−1 to maintain normocapnia, as described earlier 18. The rats were randomised to one of three ventilator strategies and ventilated for 4 h: 1) high peak inspiratory pressure (PIP)/zero PEEP (ZEEP) ventilator group, where PIP=32 cmH2O and PEEP=0 cmH2O (n=6); 2) high PIP/PEEP ventilator group, where PIP=32 cmH2O and PEEP=6 cmH2O (n=6); and 3) low PIP/PEEP group, where PIP=14 cmH2O and PEEP 6 cmH2O (n=6).
Six additional rats were ventilated according to the high PIP/PEEP group with reduced FI,O2 levels to study the effect of low (Pa,O2) levels on the immune system (hypoxic PEEP group). FI,O2 levels in this group started at 1.0 and were gradually decreased to 0.15 by increasing the amount of nitrogen. Four rats were sham-operated and anaesthetised during 4 h, and served as nonventilated controls (reference group). Blood-gas determinations were performed every hour using a pH/blood-gas analyser (ABL 505; Radiometer, Copenhagen, Denmark). In order to stabilise the high PIP/ZEEP rats haemodynamically, rats received 10 mL·kg−1·h−1 Hemohes 6% (B. Braun Melsungen AG, Melsungen, Germany). Body temperature was maintained at 37°C with heating lamps. All ventilated animals were sacrificed with an overdose of intra-arterially administered pentobarbital sodium.
Pressure/volume curves
After the animals were sacrificed the thorax was opened and the pressure/volume characteristics of the lungs in the opened chest were determined using conventional techniques 19. Maximal compliance (Cmax) was defined as the steepest part of the pressure/volume deflation curve, and was determined separately for each animal. From the pressure/volume deflation curve, the lung stability index was calculated according to Gruenwald 20.
Bronchoalveolar lavage protein and inflammatory markers in lung and plasma
Bronchoalveolar lavage (BAL) was performed with 50 mL saline (five times, 35 mL·kg body weight−1 rat, heated to 37°C) at the end of the ventilation period. BAL samples were centrifuged at 400×g for 10 min to remove cells and cellular debris, snap frozen in liquid nitrogen and stored at −80°C. Total protein concentration was measured in 1‐mL aliquots of the BAL (Bio-Rad, München, Germany). Total lungs were homogenised in radioimmunoprecipitation assay buffer with protease inhibitors (1 mM phenylmethylsulphonyl fluoride, 20 µg·mL−1 leupeptin, 200 µg·mL−1 benzamidin, 5 µg·mL−1 pepstatin) and cleaved caspase‐3 levels were determined by Western blotting. Total lung protein (50 µg) was separated by polyacrylamide gel electrophoresis and transferred to nitrocellulose. Cleaved caspase‐3 was labelled with a rabbit polyclonal antibody (Cell signaling, Beverly, MA, USA). Blots were developed using horseradish peroxidase-conjugated secondary antibody and chemiluminescence (electrochemoluminescence).
Total lung homogenates in Tris buffer (50 mM Tris, 150 mM NaCl, pH 8) were prepared for determination of TNF‐α and MIP‐2 by enzyme-linked immunosorbent assay (ELISA). In plasma, MIP‐2 expression was analysed by ELISA.
Mitogen-induced splenocyte proliferation and cytokine production
Mitogen-induced splenocyte proliferation was measured by standard procedures 21. Briefly, spleens were collected and mashed through filter chambers. Splenocytes were cultured inRoswell Park Memorial Institute (RPMI)‐1640 (Gibco, Grand Island, NY, USA) supplemented with 5% foetal calf serum (Gibco), 50 mM 2‐mercaptoethanol and antibiotics. Splenocytes were cultured in quadruplicate in 150‐µL round-bottomed microtitre wells (Costar Corp., Cambridge, MA, USA) at 1.5×105 cells·well−1 with the mitogen phytohaemagglutinin (PHA; Murex Biotech, Kent, UK). After 48 h of incubation at 37°C in a humidified atmosphere containing 5% CO2, culture supernatants were harvested and remaining cells were pulsed with 1 µCi·well−1 of 3H‐thymidine (Amersham, Buckinghamshire, UK). After 24 h, 3H‐thymidine uptake was measured using a liquid scintillation β‐counter. Supernatants were assayed for IFN‐γ (U‐Cytech, Utrecht, the Netherlands), MIP‐2 (Biosource, Camarillo, CA, USA) and IL‐10 (Pharmingen, Heidelberg, Germany), by ELISA according to the manufacturer's instructions.
Natural killer cell activity
Splenocytes were incubated for 4 h in round-bottom microtitre plates (Costar Corp.) with radiolabelled NK‐sensitive 51Cr‐YAC‐1 target cells (1×104 YAC‐1 cells·well−1). Maximal release (MR) and spontaneous release (SR) of 51Cr were determined by incubating labelled YAC‐1 cells with 1% Triton and medium, respectively. Specific killing was calculated using the formula  where X is the release of 51Cr in the experimental sample. All measurements were performed in triplicate.
where X is the release of 51Cr in the experimental sample. All measurements were performed in triplicate.
Statistical analysis
One-way analysis of variance (ANOVA) with Bonferroni post-test was used to analyse data from NK cell activity assays. Proliferative responses, BAL protein, caspase‐3 and cytokine levels were analysed after logarithmic transformation by one-way ANOVA and Bonferroni post-test. Two-way ANOVA with Bonferroni post-test was used to analyse blood gases and blood pressures. All data are expressed as mean±sem. p‐values<0.05 were considered statistically significant.
Results
Blood-gas analysis and blood pressures
Pa,O2 values at the start of MV were comparable in all groups (fig. 1⇓). During the first hour, arterial carbon dioxide tension (Pa,CO2) levels in the high PIP/ZEEP group were notably lower, due to overinflation, though not statistically different from the other groups. After 60 mins, Pa,CO2 levels were comparable in all groups. At time points 180 and 240 mins, Pa,CO2 levels in the high PIP/ZEEP group were significantly lower (p<0.05) than in the low PIP/PEEP‐ventilated group. Ventilation with high and low PIP/PEEP during 4 h did not impair mean arterial pressure (MAP) and Pa,O2 levels (except at time point 120 mins), but MAP and Pa,O2 levels in the high PIP/ZEEP‐ventilated rats significantly deteriorated over time (p<0.001 and p<0.0001, respectively). Between the three ventilated groups, pH values were not significantly different (table 1⇓).

Arterial oxygen tension (Pa,O2) levels in the low peak inspiratory pressure (PIP)/ positive end-expiratory pressure (PEEP) (▪), high PIP/PEEP (□), high PIP/zero PEEP (•) and hypoxic PEEP (○) group during 4 h of mechanical ventilation. 1 mmHg=0.133 kPa.
Blood-gas analysis and mean arterial pressures (MAP) at several time points during 4 h of mechanical ventilation with three different ventilation strategies
Pressure/volume curves and protein in bronchoalveolar lavage
In the high PIP/ZEEP group, both Cmax and the Gruenwald index were significantly lower (p<0.001) compared with the high PIP/PEEP group and low PIP/PEEP group, respectively. Protein levels in the BAL of the high PIP/ZEEP group were 4–6‐fold higher than in the high PIP/PEEP and low PIP/PEEP group (p<0.001). In addition, BAL protein levels in the high PIP/PEEP‐ventilated rats were slightly but significantly higher than protein levels in BAL from the low PIP/PEEP‐ventilated rats (p<0.05). BAL protein levels in the low PIP/PEEP group did not differ from the protein levels in the reference group (table 2⇓).
Maximal compliance (Cmax) and Gruenwald indexes and protein concentrations in bronchoalveolar lavage (BAL) fluid after 4 h of mechanical ventilation with three different ventilation strategies
Inflammatory markers in the lung
To study whether MV of healthy rats could evoke an inflammatory response in the lung, the expression of the proinflammatory cytokine TNF‐α and the chemokine MIP‐2 were analysed after 4 h of ventilation. TNF‐α was not detectable (<15 pg·mg protein−1) in total lung homogenates in any group. The expression of MIP‐2 in total lung homogenates from both high PIP‐ventilated groups was significantly higher than in lung homogenates from low PIP/PEEP‐ventilated animals (p<0.001). However, a difference in the expression of MIP‐2 was not observed between lungs from high PIP/PEEP and high PIP/ZEEP‐ventilated animals. MIP‐2 expression in the low PIP/PEEP group was not different from MIP‐2 expression in the reference group (fig. 2a⇓).

Effect of three different ventilation strategies in the healthy rat on the level of a) macrophage inflammatory protein (MIP)‐2 in total lung homogenates and b) in the plasma, and c) cleaved caspase‐3 in the lung. Rats were ventilated with low peak inspiratory pressure (PIP)/positive end-expiratory pressure (PEEP), high PIP/PEEP or high PIP/zero PEEP (ZEEP) for 4 h. Nonventilated, anaesthetised, sham-operated animals were used as a reference group. MIP‐2 expression was analysed by enzyme-linked immunosorbent assay and expression of cleaved caspase‐3 in total lung homogenates was analysed by Western blotting (50 µg protein). A positive trend was observed in the high PIP‐ventilated groups (p=0.18) for caspase‐3. ***: p<0.001.
To study whether the increase in MIP‐2 levels in the high PIP/PEEP and high PIP/ZEEP group was restricted to the lung, MIP‐2 plasma levels were also measured. In a similar manner to what was observed in the lung, MIP‐2 levels in plasma were significantly increased in both high PIP‐ventilated groups compared with the low PIP/PEEP‐ventilated group (p<0.001), and the level of plasma MIP‐2 did not differ between both high PIP‐ventilated groups. MIP‐2 was not detectable in plasma of both the low PIP/PEEP‐ventilated group as well as the reference group (fig. 2b⇑).
In addition, the level of cleaved caspase‐3 in the lung was determined as an indicator of activation of the apoptotic cascade. Surprisingly, there was no significant difference in cleaved caspase‐3 levels in lung homogenates from the four experimental groups of rats (fig. 2c⇑).
Natural killer cell activity
To investigate the effect of different ventilation strategies on the functioning of the immune system outside the lung, NK cell activity in splenocytes was determined. Splenocytes from high PIP/PEEP‐ventilated animals showed a significantly lower NK activity than splenocytes from low PIP/PEEP‐ventilated animals (p<0.001). Moreover, high PIP/ZEEP ventilation resulted in further suppression of splenic NK cell function, compared with high PIP/PEEP ventilation (p<0.01). NK activity did not differ between the low PIP/PEEP‐ventilated group and the reference group (fig. 3⇓).

Effect of three different ventilation strategies on splenocyte natural killer (NK) activity. Rats were ventilated with low peak inspiratory pressure (PIP) positive end-expiratory pressure (PEEP), high PIP/PEEP or high PIP/zero PEEP (ZEEP) for 4 h. Nonventilated, anaesthetised, sham-operated animals were used as a reference group. Moreover, the effect of hypoxia was measured in the hypoxic PEEP group. Splenocytes were collected and NK cell activity was determined using a standard 51Cr-release assay with YAC‐1 cells as target. Nonsignificant values were gained for reference versus low PIP/PEEP and for hypoxic PEEP versus high PIP/PEEP. **: p<0.01; ***: p<0.001.
Mitogen-induced splenocyte proliferation
To answer the question whether the immune system outside the lung was affected by MV, mitogen-induced proliferative responses of splenocytes were measured in vitro. After 4 h of ventilation, splenocytes from high PIP/PEEP‐ventilated animals showed a significantly lower proliferative capacity than splenocytes from low PIP/PEEP‐ventilated animals (p<0.01; fig. 4⇓). The proliferative response of the splenocytes of the high PIP/ZEEP‐ventilated group was significantly lower than in the high PIP/PEEP‐ventilated group (p<0.05) and in the low PIP/PEEP‐ventilated group (p<0.001). The mitogen-induced proliferative responses in the low PIP/PEEP group were not different from the responses in the reference group (fig. 4⇓).
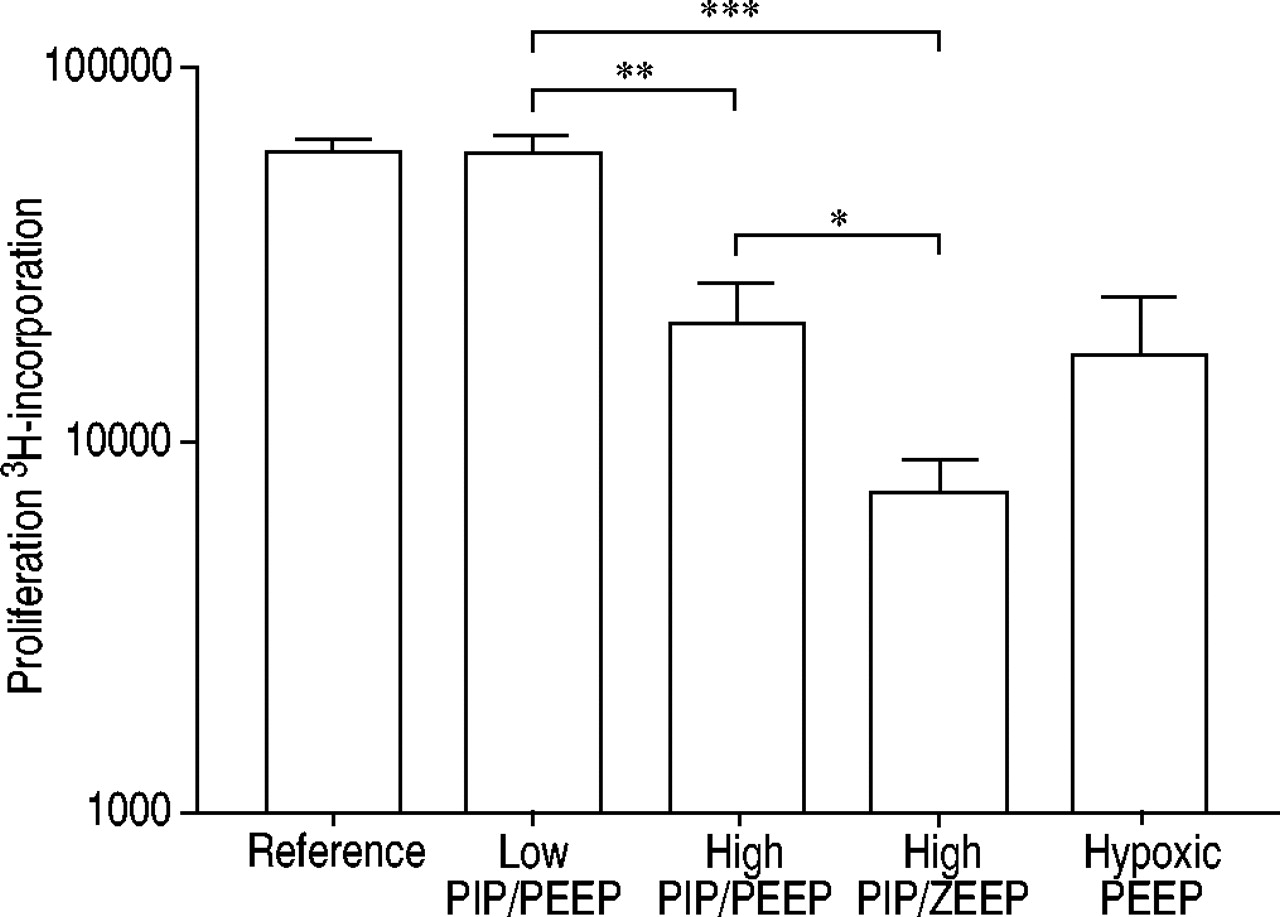
Effect of three different ventilation strategies on in vitro mitogen-induced splenocyte proliferation. Nonventilated, anaesthetised, sham-operated control animals were used as a reference group. In addition, the effect of hypoxia was determined in the hypoxic positive end-expiratory pressure (PEEP) group. Splenocytes from low peak inspiratory pressure (PIP)/PEEP, high PIP/PEEP or high PIP/zero PEEP (ZEEP)‐ventilated rats were stimulated in vitro with phytohaemagglutinin (200 µg·mL−1). After 48 h, 3H‐thymidine was added and 24 h later incorporation of 3H‐thymidine was determined. Nonsignificant values were gained for reference versus low PIP/PEEP and hypoxic PEEP versus high PIP/PEEP. *: p<0.05; **: p<0.01; ***: p<0.001.
Production of pro- and anti-inflammatory cytokines in vitro
Splenocytes from all groups were stimulated with PHA, and the production of the proinflammatory cytokine IFN‐γ, the chemokine MIP‐2, and of the anti-inflammatory cytokine IL‐10 were determined. Production of IFN‐γ, MIP‐2 and IL‐10 in the low PIP/PEEP‐ventilated group was not different from the reference group. Splenocyte MIP‐2 and IL‐10 production were significantly lower in the high PIP/PEEP group than in the low PIP/PEEP group (p<0.05 and p<0.001, respectively). Production of IFN‐γ did not differ between the high PIP/PEEP group and the low PIP/PEEP group. MIP‐2 and IL‐10 production in the high PIP/ZEEP group were significantly lower than in the high PIP/PEEP and low PIP/PEEP group (p<0.01 and p<0.001, respectively). Moreover, IFN‐γ production in the high PIP/ZEEP‐ventilated animals was significantly lower than in the low PIP/PEEP animals (p<0.05; fig. 5⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of three different ventilation strategies on in vitro mitogen-induced cytokine production. Nonventilated, anaesthesised, sham-operated animals were used as a reference group. The effect of hypoxia was determined in the hypoxic positive end-expiratory pressure (PEEP) group. Splenocytes from low peak inspiratory pressure (PIP)/PEEP, high PIP/PEEP or high PIP/zero PEEP (ZEEP)‐ventilated rats were stimulated with phytohaemagglutinin. After 48 h, supernatants were harvested. The supernatants were assayed for a) interferon (IFN)‐γ, b) macrophage inflammatory protein (MIP)‐2 and c) interleukin (IL)‐10 levels. Nonsignificant values were gained for reference versus low PIP/PEEP and hypoxic PEEP versus high PIP/PEEP. *: p<0.05; **: p<0.01; ***: p<0.001.
The effect of low arterial oxygen tension on peripheral immune function
It is possible that the more pronounced suppression of peripheral immune acitivity in the high PIP/ZEEP group compared with the high PIP/PEEP group results from the low Pa,O2 levels in the ZEEP‐ventilated rats. Therefore, the low Pa,O2 levels of the ZEEP‐ventilated rats were mimicked by reducing FI,O2 levels in a group of high PIP/PEEP rats. Pa,O2/FI,O2 ratios in the high PIP/PEEP plus low FI,O2 group (hypoxic PEEP group) were not different from Pa,O2/FI,O2 ratios in the high PIP/PEEP group, which implies that only the reduction of FI,O2, and not lung injury, was responsible for the low Pa,O2 levels in the hypoxic PEEP group. FI,O2 levels in the hypoxic PEEP group were 1.0 at the start of the experiment and were gradually decreased to 0.15 at the end of the experiment in order to induce the same Pa,O2 levels as in the high PIP/ZEEP group (fig. 1⇑). Pa,CO2 was kept constant by adjusting the frequency of ventilator (data not shown).
Comparing NK cell activity, splenocyte proliferation and cytokine production in splenocytes from hypoxic PEEP‐ventilated rats and high PIP/PEEP‐ventilated rats revealed that the reduced Pa,O2 levels in the hypoxic PEEP group did not result in reduced NK cell activity (fig. 3⇑), splenocyte proliferation (fig. 4⇑) or cytokine production (fig. 5⇑) (unpaired t‐test p>0.05).
Discussion
The major finding of this study is that 4 h of high peak pressure ventilation resulted in a marked reduction in peripheral immune functioning; NK cell activity, mitogen-induced splenocyte proliferation and production of MIP‐2 and IL‐10 decreased significantly after high peak pressure ventilation with or without application of PEEP. Although application of PEEP completely prevented deterioration of lung function, it only partially attenuated the observed peripheral immunosuppression. Lung and plasma MIP‐2 levels increased after 4 h of high PIP ventilation.
The increased BAL protein and the decreasing Pa,O2 levels in the high PIP/ZEEP group suggest pulmonary oedema, enhanced alveolar-capillary membrane permeability and hence VILI. It has been shown repeatedly that ventilating healthy animals, with a large tidal volume or high PIP without PEEP damages the lung and causes VILI 3, 4. In addition, it has been shown that application PEEP may prevent such damage 2. Indeed, the present data confirm that high PIP/PEEP ventilation results in maintenance of lung function (Pa,O2 and compliance) similarly to low PIP/PEEP‐ventilated animals.
In humans with acute lung injury the apoptotic cascade in the lung is activated 22. Therefore, it is expected that the apoptotic marker cleaved caspase‐3 to be upregulated in the high PIP/ZEEP‐ventilated group. However, a significant difference was not observed in cleaved caspase‐3 levels in the lung between the ventilated groups.
Previous ex vivo and in vivo studies in healthy animals and animals with pre-existing lung injury showed that especially injurious ventilator strategies can induce an inflammatory reaction in the lung, characterised by sequestration of neutrophils and local production of cytokines 5, 23, 24. In line with recent data of Verbrugge et al. 18 and Ricard et al. 25, the present authors have shown that injurious ventilation strategies do not necessarily evoke TNF‐α expression in the rat lung.
However, the present data show that the expression of MIP‐2 in the lung and plasma was elevated after 4 h of MV with high PIP, irrespective of PEEP application, which is in contrast to the data of Ricard et al. 25 who found no differences in MIP‐2 expression in the lung and no expression of MIP‐2 in plasma. Apparently, high peak ventilation can evoke an inflammatory response in the lung and plasma, even when shear forces in the lung are minimised. Since MIP‐2 levels were similar in both high PIP‐ventilated groups, there isno evidence for a direct relationship between injurious ventilation-induced lung dysfunction and MIP‐2 levels in lung tissue.
Recently, the authors described that in children without lung pathology, 2 h of MV with mean tidal volumes of 10 mL·kg−1 and 4 cmH2O of PEEP induces an inflammatory state in the lung characterised by TNF‐α in BAL 17. Interestingly, in these children, a marked decrease in peripheral blood NK cell activity and a decrease in in vitro IFN‐γ production were observed, whereas IL‐10 production did not change. In this experimental rat study, a decrease in NK cell activity in the high PIP‐ventilated groups was observed compared with the group that was ventilated with a low PIP strategy. In addition, mitogen-induced splenocyte proliferation and IL‐10 production were reduced in both the high PIP/PEEP and the high PIP/ZEEP group. In the human study, aneffect of anaesthesia could not be excluded as such. In this rat study, however, all animals received the same anaesthesia. Therefore, the difference in peripheral immune function between the high PIP/PEEP and low PIP/PEEP group cannot be attributed to anaesthesia.
In the present study, the findings of the previous clinical study 17 were specified and extended, by demonstrating that ventilation strategies leading to increased inflammation in the lung was accompanied by a reduced function of the immune system outside the lung. These data are in line with studies that demonstrate that the peripheral immune response can be reduced in ventilated intensive care patients, which may contribute to increased morbidity and mortality 12, 13. A recent experimental study by Lin et al. 26 demonstrated that 1 h of injurious ventilation in rats, before instillation of Pseudomonas aeruginosa, resulted in an enhanced susceptibility to bacteraemia. The authors suggested that apart from local epithelial damage and subsequent bacterial translocation, decreased systemic cytokine levels rendered the animals more susceptible to bacteraemia 26. Based on these results, the present authors suggest that peripheral immunosuppression may have contributed to the increased vulnerability to bacteraemia, possibly by a decreased clearance of translocated bacteria.
The suppression of the peripheral immune response in highPIP/ZEEP‐ventilated rats was more pronounced than in the high PIP/PEEP‐ventilated rats. Hypoxia per se is not responsible for the more pronounced reduction in immune function in the high PIP/ZEEP‐ventilated rats. In fact Pa,O2 levels deteriorated during 4 h of high PIP/ZEEP ventilation, but similar low Pa,O2 levels in the hypoxic PEEP group, obtained by reducing the FI,O2, did not result in further reduction of immune activity. Although it cannot be excluded that Hemohes contributed to the more severe immunosuppression in the high PIP/ZEEP group, it is important to realise that a significant decrease in NK activity and splenocyte proliferation was also found in the high PIP/PEEP‐ventilated group. This high PIP/PEEP‐ventilated group had normal blood pressures and did not receive Hemohes suppletion. In addition, it cannot be excluded that low blood pressure, due to injurious ventilation in the high PIP/ZEEP‐ventilated group, may have affected the results.
Some experimental and human studies have suggested that MV can induce systemic inflammation by loss of compartmentalisation between lung and systemic circulation and spill-over of inflammatory mediators 7, 10, 27. In ARDS patients, lung protective strategies appeared to reduce plasma TNF‐α, IL‐β and IL‐6 levels 10. The spill-over of cytokines has been implicated in the fact that most ARDS patients do not die from lung injury but from MSOF 8–11. An increase in MIP‐2 expression was found in the plasma of both high PIP‐ventilated groups, but reduced splenic in vitro production of MIP‐2. Therefore, these data suggest that the increased plasma MIP‐2 levels may result from spill-over of cytokines from the lung and could contribute to the peripheral immune suppression. In line with this hypothesis, Suter 28 suggested that peripheral immunosuppression or “icy peripheral leukocytes” may be the result of inflammation or “fire” in the lung as a consequence of MV.
To conclude, the authors have demonstrated that 4 h of mechanical ventilation of healthy rats does evoke an inflammatory response in the lung. More importantly, they have shown that mechanical ventilation can lead to impairment of peripheral immune function even when normal lung function is maintained. Since many critically ill patients require mechanical ventilation, it is important to realise that mechanical ventilation cannot only enhance inflammation in the lung, but may impair the functioning of peripheral lymphocytes, which may affect the susceptibility to infections and may contribute to multiple system organ failure.
Acknowledgments
The authors would like to thank S. Krabbendam, J. Goorman and F. Delreu for expert technical assistance, and L. Visser-Isles for English-language editing (all Erasmus MC‐Faculty).
- Received March 28, 2003.
- Accepted July 21, 2003.
- © ERS Journals Ltd
References









