





Abstract
Exposure in swine confinement buildings causes intense airway inflammation and lymphocyte activation, as assessed by bronchoalveolar lavage. To further clarify the T-cell activation, the present in vitro study focused on intracellular cytokine production following exposure to organic dust from swine houses.
Whole blood from healthy donors was incubated with swine dust, phytohaemagglutinin (positive control) or Roswell Park Memorial Institute 1640 medium (negative control), and the production of intracellular interferon (IFN)-γ, tumour necrosis factor (TNF)-α, interleukin (IL)-4 and IL-12 was analysed by flow cytometry. Cells were double stained for specific cell surface markers on T-cells (CD3+, CD4+, CD8+), natural killer (NK) cells (CD56+CD16+) and monocytes (defined as CD14+ cells).
Following 1 h of incubation of whole blood with swine dust, CD14+ cells produced high levels of TNF-α and IL-12, whereas CD3+, CD4+ and CD8+ T-cells and CD56+CD16+ NK cells required a longer incubation time (22 h) to produce IFN-γ and TNF-α. When antibodies that block the IL-12 receptor were added to whole blood incubated with swine dust, NK cell production of IFN-γ was attenuated and CD69 expression on CD3+ cells decreased.
In conclusion, this study indicates that swine dust can, at least in part, stimulate phagocytic cells to activate natural killer cells and T-lymphocytes through the production of interleukin-12.
The present authors have previously demonstrated that exposure to dust in a swine confinement facility induces activation of T-cells in vivo, as assessed by analyses of cells obtained through bronchoalveolar lavage 1. The authors have also shown that T-cells in whole blood exposed to dust in vitro express the activation markers CD69, CD25 and human leucocyte antigen-DR in a dose- and time-dependent manner 2. Plasma obtained from blood that had been incubated with dust for 24 h, i.e. “conditioned plasma”, induced a high CD69 expression on T-cells (CD3+) in whole blood 3. Since no increased CD69 expression on T-cells was observed in blood depleted from phagocytic cells, and since an increased production of interleukin (IL)-12 and interferon interferon (IFN)-γ was detected in whole blood stimulated with swine dust, the authors suggested that soluble mediators derived from phagocytic cells may be responsible for the lymphocyte activation.
It is known that cytokines, such as tumour necrosis factor (TNF)-α and IL-12, are capable of inducing T-cell activation 4, 5. IL-12 is produced by activated monocytes but also by other antigen-presenting cells and neutrophils, and it has pleiotropic effects on T-cells and natural killer (NK) cells 6. The aim of the present study was to investigate whether swine dust could induce phagocytic cells (CD14+ monocytes) to produce the cytokines TNF-α and IL-12 and/or T- and NK cells to produce TNF-α, IFN-γ and IL-4. In addition, the authors wanted to clarify the role of IL-12 in lymphocyte activation by using of IL-12 receptor blocking antibodies.
Materials and methods
Subjects
Venous blood was collected from 10 (two males) nonallergic, nonsmoking, healthy subjects, with a mean age of 44 yrs (range 27–60). All subjects gave their informed consent and the study was approved by the Ethics Committee of Karolinska Institutet, Stockholm, Sweden.
Stimulation with dust
Dust was collected in a swine confinement building ∼1.5 m above the ground and dissolved in Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco Laboratories, Paisley, UK), sonicated for 10 min and diluted in whole blood to a final concentration of 100 µg·mL−1. The polyclonal activator, phytohaemagglutinin (PHA), used as positive control in T- and NK cell analysis, was also dissolved in RPMI 1640, sonicated for 10 min and diluted in whole blood to a final concentration of 10 µg·mL−1. Unstimulated whole blood containing RPMI 1640 was used as negative control.
Staining of surface markers and intracellular cytokines
For analyses of monocytes (CD14+ cells) (n=6), whole blood was incubated for 1 h with dust or RPMI 1640 in the presence of brefeldin A (Golgi PlugTM; PharMingen, San José, CA, USA). Brefeldin A inhibits the intracellular transport, implying that cytokines produced during the activation are intracellularly retained (protocol according to the manufacturer).
For lymphocyte analyses (n=10), whole blood was diluted 1:1 with RPMI containing L-glutamine and incubated for 12 h with swine dust or PHA followed by an additional 10-h incubation in the presence of 1 µg brefeldin A·mL blood−1. All incubations were performed at 37°C in the presence of 5% carbon dioxide.
After stimulation, red blood cells were lysed (PharmLyse; PharMingen) and washed with staining buffer (phosphate buffered saline (PBS) containing 1% foetal calf serum). The cells were centrifugated to a pellet at 500×g for 5 min, the supernatant was aspirated and cells (∼200×103) were diluted in 100 µL staining buffer. The respective cell subset and/or activation marker-specific monoclonal antibody (Mab) was added and incubated for 30 min, washed in staining buffer and then fixed and permeabilised by adding Cytofix/Cytoperm solution (PharMingen) for 20 min. Cells were then washed with Perm/Wash solution (PharMingen) and incubated for 10 min at room temperature (RT) in the dark. They were then centrifuged for 5 min at 500×g. The supernatant was aspirated and the cell pellet was resuspended in Perm/Wash solution containing cytokine-specific Mab for intracellular staining and incubated for 30 min at RT in the dark. Cells were washed with Perm/Wash solution (without Mab) and thereafter centrifuged for 5 min at 500×g. The supernatant was aspirated and the cells were resuspended in PBS containing 4% formaldehyde (Cellfix; Becton Dickinson, San José, CA, USA). The cells were then analysed by flow cytometry.
In a second experimental set up (n=6), a combination of the Mabs 2.4E6 and 2B10 (PharMingen) directed to the IL-12 receptor β1 subunit, were used to block the human IL-12 receptor 7. Blocking antibodies or isotype controls (mouse immunoglobulin (Ig)G1 and rat IgG2a; PharMingen) were added 1 h prior to dust incubation. Cells were then incubated with dust, PHA or RPMI. Cytokine production by T- and NK cells as well as T-cell CD69 expression were investigated with and without IL-12 receptor blocking Mab.
Monoclonal antibodies and flow cytometric analysis
Cells were characterised by subgroup specific fluorochrome conjugated Mab. For identification of T-cells, CD3, CD4, CD8-peridinin clorophyll protein (PerCP) conjugated Mab (Becton Dickinson) were used, while NK cells were identified using CD56+CD16+−phycoerythrin (PE) conjugated Mab (Becton Dickinson). Monocytes were identified by forward and sidescatter parameters and by staining with CD14-PerCP (Becton Dickinson). T-cell activation was measured by double staining CD3 and the early activation marker CD69, using flourescein isothiocyanate (FITC)-conjugated anti-CD3 Mab and PE-conjugated anti-CD69 Mab (Becton Dickinson).
IFN-γ, IL-4 and TNF-α specific Mabs were obtained from Becton Dickinson and IL-12 specific Mab were obtained from PharMingen. All intracellular cytokine specific Mab were FITC conjugated. Cells were analysed by flow cytometry using fluorescence-activated cell sorter Calibur (Becton Dickinson).
Positive cells were expressed as percentages among the different subsets (CD3+, CD4+, CD8+ T-cells, NK cells and monocytes).
Statistics
Results are presented as medians (25th–75th percentiles). Comparisons were performed by analysis of variance (ANOVA). All p-values of <0.05 were considered as statistically significant.
Results
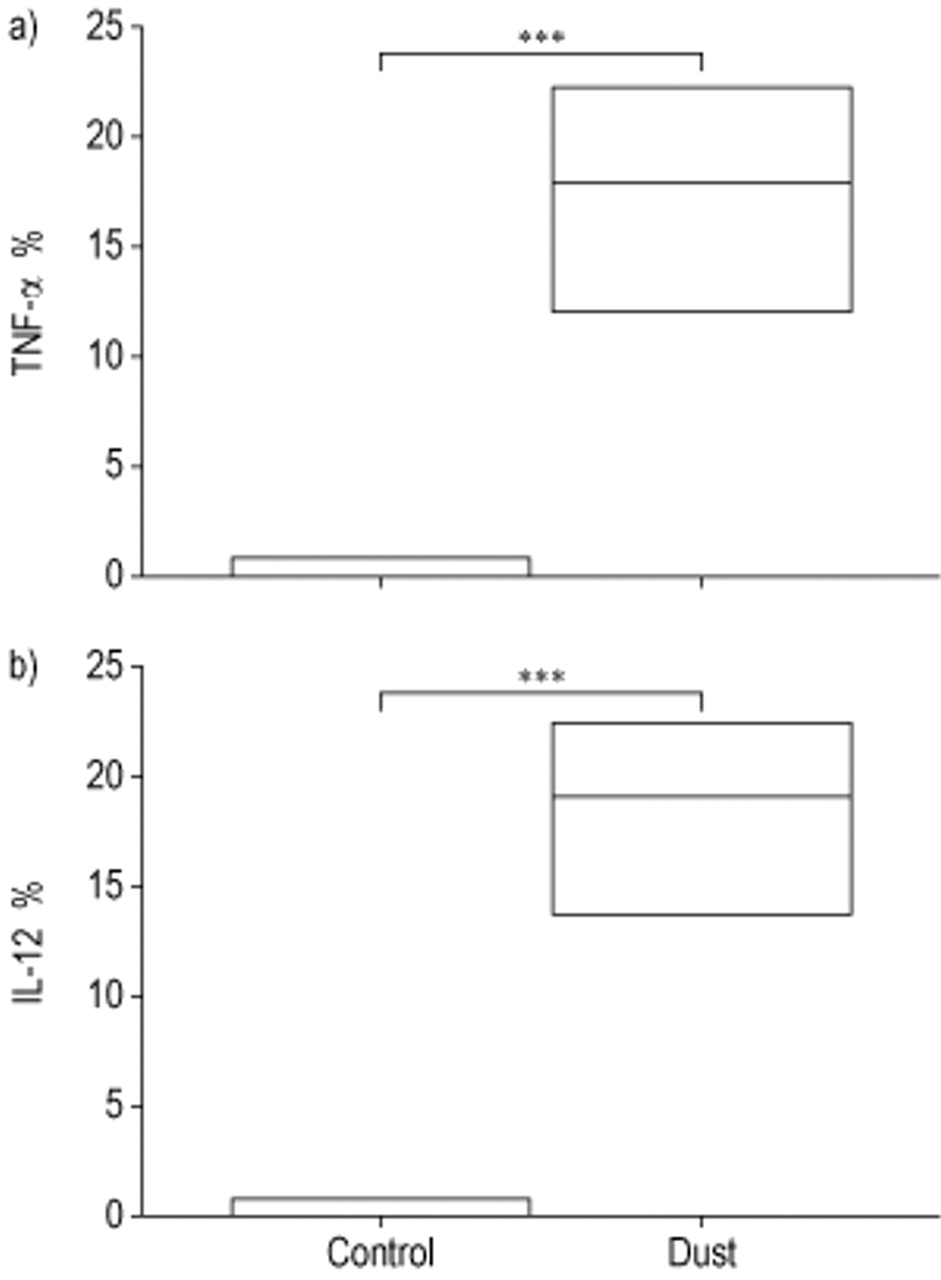
Incubation with dust for 1 h induced a significant production of TNF-α in the monocytes, i.e. CD14+ peripheral blood cells (n=6), compared to nonstimulated control cells (p<0.001, fig. 1a⇓). Similarly, dust induced a significant IL-12 production in the monocytes compared to the negative control (p<0.001, fig. 1b⇓).

Intracellular a) tumour necrosis factor (TNF)-α and b) interleukin (IL)-12 production by CD14+ cells (monocytes) under control conditions (control) or following 1-h incubation with swine dust (dust) (n=6). Data are presented as median and 25th–75th percentiles. ***: p<0.001.
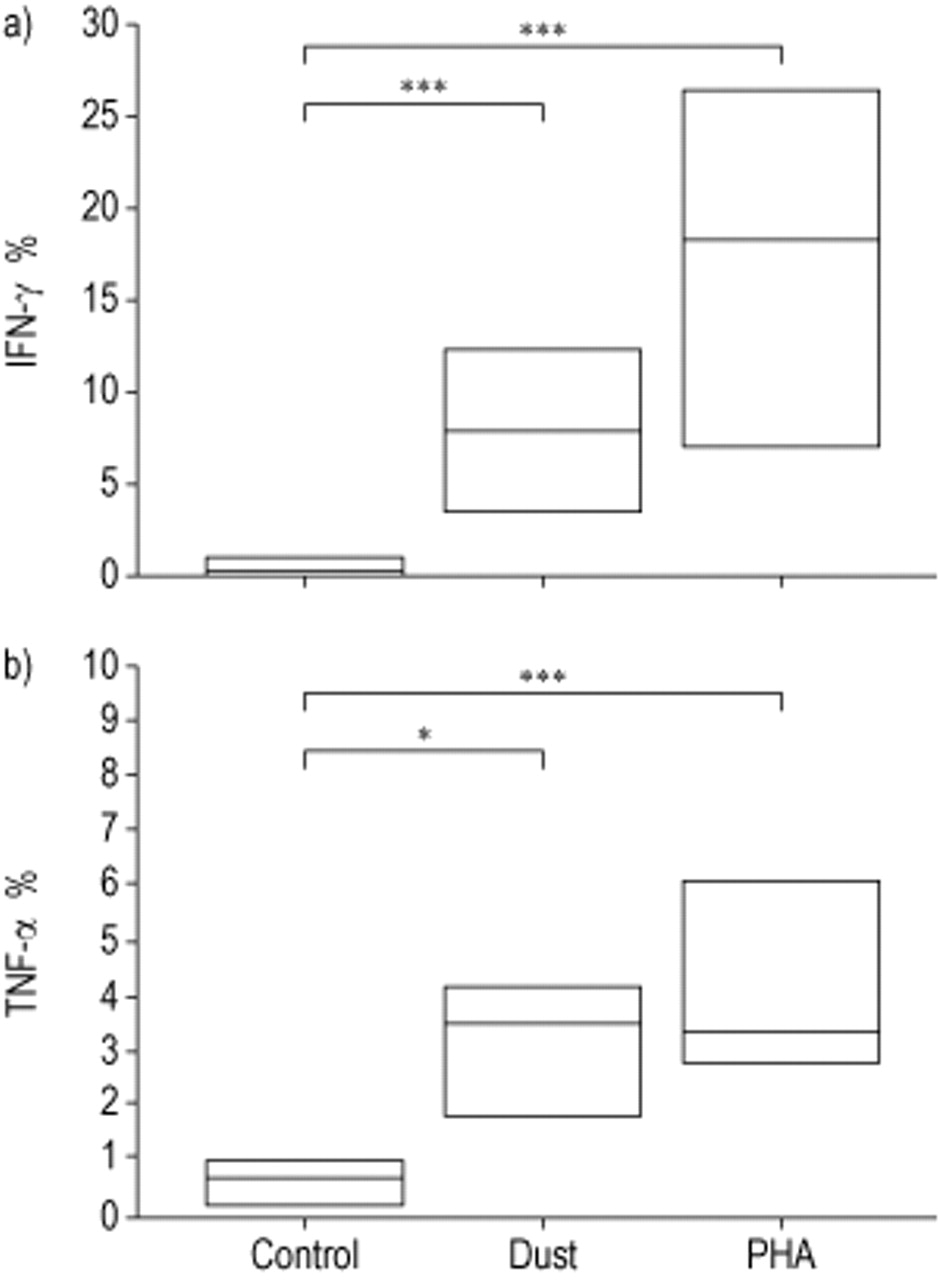
No intracellular cytokine production was observed in lymphocytes after 1 h of dust incubation. After time kinetic pilot experiments, 22-h incubation, including brefeldin A for the last 10 h, was used. The dust induced significantly higher IFN-γ levels in NK cells after 22-h incubation compared to negative controls (p=0.001, fig. 2a⇓). Similarly, NK cells produced significantly more TNF-α compared to negative controls (p<0.05, fig. 2b⇓). PHA (positive control) also induced high levels of IFN-γ and TNF-α in NK cells (p<0.001, fig. 2a and b⇓).

Intracellular a) interferon (IFN)-γ and b) tumour necrosis factor (TNF)-α production by natural killer cells under control conditions (control), following 22-h incubation with swine dust (dust) and phytohaemagglutinin (PHA). Data are presented as median and 25th–75th percentiles (n=10). *: p<0.05; ***: p<0.001.
CD3+ T-cells, CD4+ T-cells and CD8+ cells only produced low levels of IFN-γ, TNF-α and IL-4 after dust stimulations (table 1⇓), although the increased production of IFN-γ by CD4+ and CD8+ T-cells was significant (p<0.05 and p=0.05, respectively).
Cytokine production by T-cell subsets after stimulation with organic dust
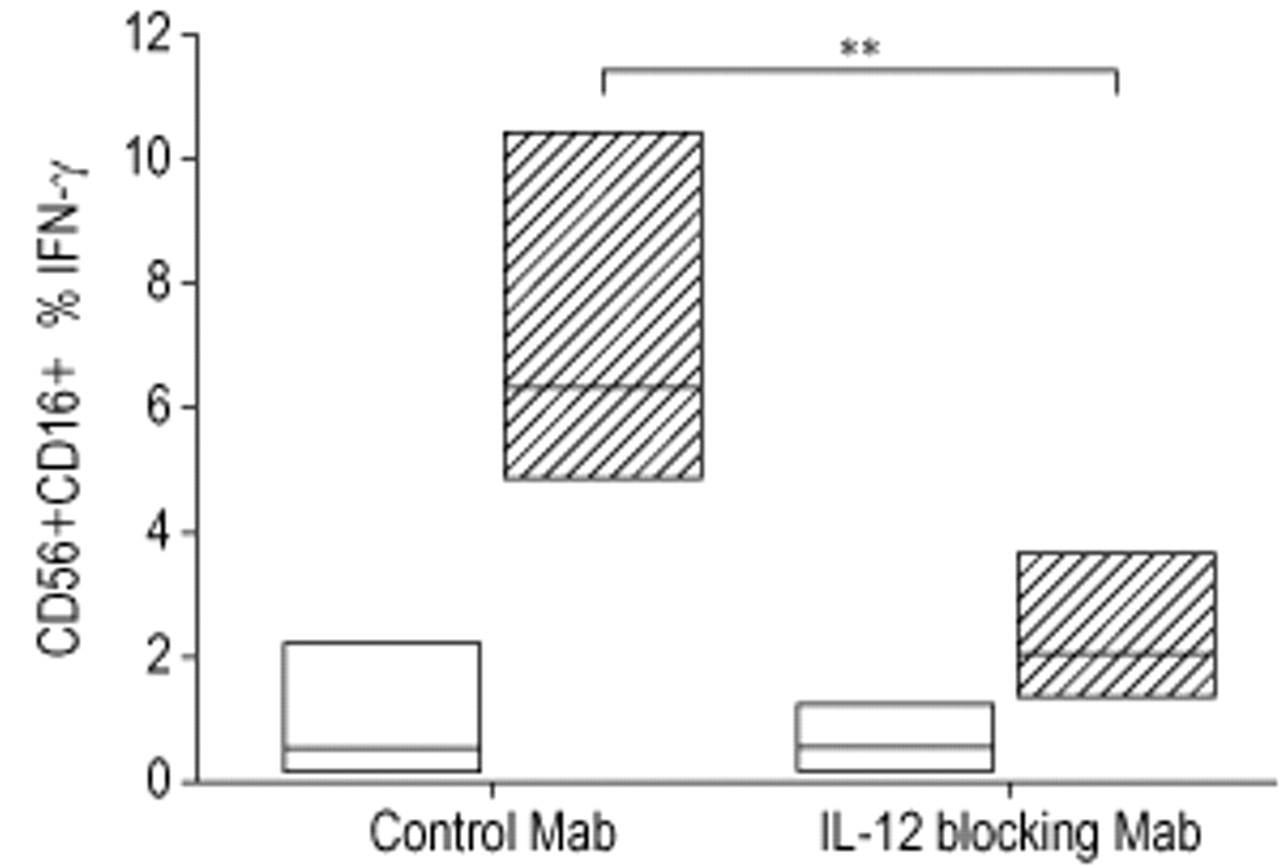
A reduction in dust-induced IFN-γ production by NK cells from 6.3–1.8% was observed in the presence of IL-12 receptor blocking antibodies (n=6, p=0.017), while no difference in the NK cell production of IFN-γ was found in the presence of control antibodies (fig. 3⇓). CD69 expression on T-cells was reduced from 19.1% (17.1–28.2) to 12.2% (11.5–20.2) when whole blood was stimulated by dust in the presence of IL-12 receptor blocking antibodies (fig. 4⇓).

Intracellular interferon (IFN)-γ production in natural killer cells in blood preincubated with control antibodies (control Mab) or blocking antibodies to the interleukin-12 receptor (IL-12 blocking Mab), following 22-h incubation with (┘) or without (□) organic dust. Data are presented as median and 25th–75th percentiles (n=6). **: p=0.01.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
CD69 expression on T-cells in blood following 22-h incubation with Roswell Park Memorial Institute (RPMI) medium only (control), with organic dust only (dust), with organic dust and control antibodies (dust+control Mab), or with organic dust and blocking antibodies to the interleukin-12 receptor (dust+blocking Mab). Data are presented as individual CD69 expressions on CD3+ T-cells.
Discussion
In the present study, it was shown that monocytes in peripheral whole blood could be stimulated to produce TNF-α and IL-12 after 1-h incubation with dust from a swine confinement facility. No intracellular cytokine production was observed in lymphocytes after incubation with dust at this early time point. However, as seen in the findings of Lertmemongkolchai et al. 8, after 22-h incubation with dust, both T-lymphocytes and NK cells produced IFN-γ and low levels of TNF-α and IL-4. Furthermore, it was shown that blocking of the IL-12 receptor inhibited the dust-induced IFN-γ production by NK cells as well as T-cell activation.
In time kinetic studies of whole blood stimulated with dust, it was found that 1-h incubation was enough to induce monocytes to produce TNF-α and IL-12. A previous report 9 also showed a rapid IL-12 production by monocytes after stimulation with Trypanosoma cruzi. The present authors previously found 24 h of incubation to be optimal with regard to T-cell activation by swine dust. In the present study, the incubation was reduced to 22 h, with only 10 h brefeldin A, due to the toxicity of this substance.
The present authors have previously reported that dust failed to activate T-cells in peripheral blood depleted from phagocytes 3. However, plasma separated from dust-incubated whole blood, i.e. conditioned plasma, and added to peripheral blood mononuclear cells over night, was highly capably of inducing CD69 expression on T-cells (CD3+). Such conditioned plasma contained high levels of IL-12 and IFN-γ, probably released from phagocytic cells upon stimulation. Thus, IL-12 may be one of the components required for lymphocyte activation by the organic dust. This suggestion is supported by the finding that IL-12 is required for lipopolysaccharide (LPS)-induced activation of NK cells, and that no IL-12 was found in monocyte-depleted cell cultures where the cells were stimulated with LPS 10.
In the present study, whole blood was preincubated for 1 h with IL-12 receptor blocking Mabs. Used in combination, these two Mabs have an inhibitory effect on the IL-12-mediated functions 7. A significantly decreased production of IFN-γ by NK cells was found when blood was preincubated with IL-12 receptor blocking antibodies. Any influence on T-cell IFN-γ production was difficult to evaluate due to the low T-cell IFN-γ levels. However, preincubated blood with IL-12 receptor blocking antibodies clearly decreased the activation of T-cells, as judged from the reduced CD69 expression.
One mechanism through which monocytes recognise swine dust antigens could be via the CD14 receptor to LPS 11, and the dust is known to contain small amount of LPS 12. Interestingly, human T-lymphocyte activation by LPS has been shown to be dependent on monocytes, and IL-12 to be one of the important costimulatory factors 13. In the present study, monocytes rapidly produced TNF-α and IL-12 after incubation with swine dust but when stimulated with LPS only, no distinct IL-12 production was found (data not shown). Thus, other components than LPS are also important for the organic dust stimulation of monocytes.
In conclusion, it has been shown that organic dust can stimulate interleukin-12 production in CD14+ cells and interferon-γ production in T- and natural killer cells. In the presence of dust and interleukin-12 blocking antibodies, the interferon-γ production was reduced in natural killer cells as was the CD69 expression on T-cells. In order to clarify the mechanism for in vivo activation of human T- and natural killer cells further studies will be necessary, including cytokine analyses in bronchoalveolar lavage fluid cells and peripheral blood cells before and after exposure of healthy volunteers to swine dust.
- Received March 12, 2002.
- Accepted April 23, 2002.
- © ERS Journals Ltd
References









