





Abstract
Exercise-induced bronchoconstriction is associated with heat and water loss from the airways. It is not known whether these conditions can influence the response to bronchoactive agonists. The effects of different degrees of alveolar ventilation on the pulmonary response to methacholine and the role of humidity and temperature in this response were evaluated.
Wistar rats were anaesthetized, tracheostomized and mechanically ventilated. Increasing doses of methacholine were infused intravenously and respiratory system resistance (Rrs) and elastance (Ers) were measured. The rats were ventilated with dry air at 13°C, dry air at 37°C, humid air at 13°C and humid air at 37°C. These four groups were further divided into three subgroups with a respiratory frequency adjusted to reach a carbon dioxide tension in arterial blood of 30, 40 and 50 mmHg.
Temperature, humidity and level of alveolar ventilation did not influence the position of the dose/response curve to methacholine. However, the maximal changes in Ers were significantly lower in the rats ventilated with humid air. In addition, maximal changes in Ers were significantly higher in the rats with lower alveolar ventilation. These differences were not observed for maximal values of Rrs.
The pulmonary response to methacholine in normal rats is significantly affected by the humidity of inspired air and the level of alveolar ventilation. This influence is more intense in the small airways and/or distal airspaces. This suggests that exercise or hyperventilation can change the behaviour of airway smooth muscle.
- airway smooth muscle
- alveolar ventilation
- experimental asthma
- hyperventilation-induced bronchospasm
- pulmonary responsiveness
This study was supported by the following Brazilian Scientific Agencies: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and Programa de Núcleos de Excel̂ncia (PRONEX-MCT).
Many inflammatory mediators, neurotransmitters and drugs can induce contraction of airway smooth muscle (ASM). The role of ASM in normal respiration is still the subject of debate, but it is unlikely that baseline airway tone is physiologically important, since bronchodilation induced by muscarinic antagonists or β2-agonists does not appear to have adverse effects on airway function 1. Most of the interest in ASM response to contractile agonists lies in the role of ASM in airway diseases, particularly asthma.
Airway hyperresponsiveness to nonspecific stimuli, such as methacholine, histamine, cold air or exercise, is an important feature of asthma. Airway hyperresponsiveness has been defined as an exaggerated response of the airways to nonspecific stimuli, resulting in airway obstruction, mainly due to excessive ASM contraction 2.
ASM can be constricted directly by agonists, such as methacholine or histamine, which activate receptors on the smooth-muscle cells, or by indirect mechanisms, such as cold air or exercise, which, at least in part, induce the release of bronchoactive mediators from mast cells 1, 2.
Exercise and hyperventilation result in similar transient increases in airway resistance in patients with asthma. It is generally accepted that the pathogenesis of exercise-induced bronchoconstriction (EIB) in asthma is associated with the fluxes of heat and water that develop in the airways during the warming and humidification of large volumes of air. The loss of both heat and water play a pivotal role in EIB and bronchospasm induced by hyperventilation. In addition, the inspiration of low-temperature air and dry air result in an increased prevalence of EIB compared to warm and humid air 3–5. However, the relative effects of the degrees of ventilation, humidity and temperature on the pulmonary responsiveness to a bronchoactive agonist has not yet been studied. The authors reasoned that this kind of study could add relevant information to understanding of airway hyperresponsiveness and exercise-induced bronchospasm.
Therefore, the aims of the present study were: 1) to study the effects of different degrees of alveolar ventilation on the pulmonary response to methacholine; and 2) to evaluate the effects of dry and humid air and of different temperatures of inspired air on these responses.
Materials and methods
Study animals
Male Wistar rats (250–350 g), obtained from the School of Medicine of the University of Sao Paulo (Sao Paulo, Brazil) were used. The rats were housed in conditions of a constant temperature and relative humidity and were fed a standard rat diet. Animals were kept free from all evidence of infectious diseases. All animals received humane care in compliance with the Helsinki convention for the use and care of animals. The study was approved by the Institutional Review Board of the School of Medicine of the University of Sao Paulo.
Study design
The rats were anesthetized, tracheostomized and mechanically ventilated and randomly assigned to 12 groups (n=6 for each group). The rats were ventilated with either dry air at 13°C, dry air at 37°C, humid air at 13°C and humid air at 37°C. These groups were further divided into three subgroups with a respiratory frequency adjusted to reach a carbon dioxide tension in arterial blood (Pa,CO2) of 30, 40 and 50 mmHg. Increasing doses of methacholine were infused intravenously, tracheal pressure, airflow and lung volume changes were obtained and respiratory system resistance (Rrs) and elastance (Ers) were calculated.
Methods
The rats were anaesthetized with pentobarbital sodium (50 mg·kg−1 i.p.). Once surgical-level anaesthesia was achieved, the animals were secured supine, a tracheotomy was created and a polyethylene tube (internal diameter=1.7 mm) was inserted into the trachea. A jugular vein was cannulated to infuse methacholine solutions and a carotid artery was cannulated to drain arterial blood for measurements of pH, oxygen tension in arterial blood (Pa,O2) and Pa,CO2, using a Stat Profile 10 (Nova Biomedical Inc., Waltham, MA, USA). Both procedures were performed using a polyethylene tube (internal diameter=1.1 mm) and the arterial line was kept free from blood clotting with heparin solution at 10 U·mL−1.
The animals were ventilated with a small animal ventilator (Harvard 683; Harvard Apparatus, South Natick, MA, USA) with a tidal volume (VT) of 10 mL·kg−1. The respiratory rate was adjusted to achieve the Pa,CO2 previously assigned (30, 40 or 50 mmHg ±10%). The range was 60–90 breaths·min−1 (table 1⇓). In order to wet the ventilation gas, a bypass system was used where the gas passed under a water column in a vaporization camera, set to 13 or 37°C. Humidity was measured at the vaporization camera and temperature was monitored just before the entry of the tracheal tube.
Values of respiratory frequency (RF) carbon dioxide tension in arterial blood (Pa,CO2) oxygen tension in arterial blood (Pa,O2) and pH in the 12 groups of rats studied
Rats were randomly assigned to 12 groups. Animals were ventilated with: 1) dry air (3 parts per million (ppm) of H2O) at 13°C (D13); 2) dry air at 37°C (D37); 3) humid air (100% saturation) at 13°C (H13); 4) humid air at 37°C (H37). Each one of these groups were further divided into three subgroups that had the frequency of the ventilator adjusted to reach a Pa,CO2 of 30, 40 and 50 mmHg (n=6 for each final group).
Measurements of Rrs and Ers were performed as previously described 6, 7. A pneumotachograph (Fleish-4.0; OEM Medical Inc., Richmond, VI, USA) was connected to the tracheal tube and to a differential pressure transducer (Honeywell 163PC01D36; Freeport, IL, USA) to measure airflow (V′). Tracheal pressure (Ptr) was measured with a pressure transducer (Honeywell 142PC05D). V′ and Ptr signals were measured and conditioned during 10 s at 200 Hz with a 12-bit analog-to-digital converter (DT 01-EZ; Data Translation, Marlboro, MA, USA) and stored in a microcomputer. In each measurement, 10, 13 or 15 cycles, according to the respiratory frequency setup, were analysed for each data point. Lung volume changes (V) were obtained by electronic integration of the V′ signal.
Rrs and Ers were obtained using the equation of motion of the respiratory system: where (t) is time. After the rats had achieved the desired Pa,CO2, the groups of animals designed to proceed under dry ventilation had their Ptr and V′ measured (baseline). V′ was also measured directly after achieving a maximal increase in Ptr that followed each intravenous infusion of increasing concentrations of methacholine chloride (Sigma Chemical Co., Saint Louis, MI, USA) in normal saline (0.1–300 µg·kg−1). The groups of animals designed to proceed under humid ventilation were kept under those conditions for an additional 10 min. Ptr and V′ were measured (baseline) and the same protocol of methacholine infusion described earlier was followed. All rats received intermittent deep inflations (3×VT) every 15 min from the start of mechanical ventilation. The last deep inflation was performed immediately before the beginning of methacholine challenge. The methacholine dose/response curve lasted 30 min. The authors previously observed that 30 min of mechanical ventilation with either dry or humid air and without deep inflations did not change Rrs and Ers.
where (t) is time. After the rats had achieved the desired Pa,CO2, the groups of animals designed to proceed under dry ventilation had their Ptr and V′ measured (baseline). V′ was also measured directly after achieving a maximal increase in Ptr that followed each intravenous infusion of increasing concentrations of methacholine chloride (Sigma Chemical Co., Saint Louis, MI, USA) in normal saline (0.1–300 µg·kg−1). The groups of animals designed to proceed under humid ventilation were kept under those conditions for an additional 10 min. Ptr and V′ were measured (baseline) and the same protocol of methacholine infusion described earlier was followed. All rats received intermittent deep inflations (3×VT) every 15 min from the start of mechanical ventilation. The last deep inflation was performed immediately before the beginning of methacholine challenge. The methacholine dose/response curve lasted 30 min. The authors previously observed that 30 min of mechanical ventilation with either dry or humid air and without deep inflations did not change Rrs and Ers.
Data analysis
To compare the dose/response curves to methacholine, the doses of methacholine that caused 50% of maximal changes in Rrs and Ers (termed KxRrs and KxErs, respectively) were calculated for each animal, according to Hulbert et al. 8. The maximal changes in Rrs and Ers (termed ΔmaxRrs and ΔmaxErs, respectively) were calculated. Kx and Δmax values for both Ers and Rrs were compared using a three-way analysis of variance (factors Pa,CO2, temperature and humidity) followed by the Tukey Test for multiple comparisons 9.
Baseline values of Rrs and Ers, respiratory frequency, pH, Pa,O2 and Pa,CO2 were also examined using a three-way analysis of variance. A p-value <0.05 was considered significant.
Results
Table 1⇑ shows the respiratory frequency, Pa,CO2, Pa,O2 and pH (means±sem) values obtained in each of the 12 groups studied. At each set Pa,CO2 level (30, 40 or 50 mmHg), no significant differences in respiratory frequency, pH, Pa,CO2 and Pa,O2 values were observed when groups D13, D37, H13 and H37 were compared. In the ventilated groups that had lower respiratory frequencies there was a significant decrease in pH due to higher values of Pa,CO2 as well as significantly lower values of Pa,O2, since all animals were ventilated with a concentration of oxygen of 21% in the inspired air.
Table 2⇓ shows baseline values of Rrs and Ers. These values were obtained immediately before the beginning of methacholine challenges. There was no significant influence of temperature or humidity of the inspired air or the level of alveolar ventilation on basal values.
Values of respiratory system resistance (Rrs) and elastance (Ers) in the 12 groups of rats studied
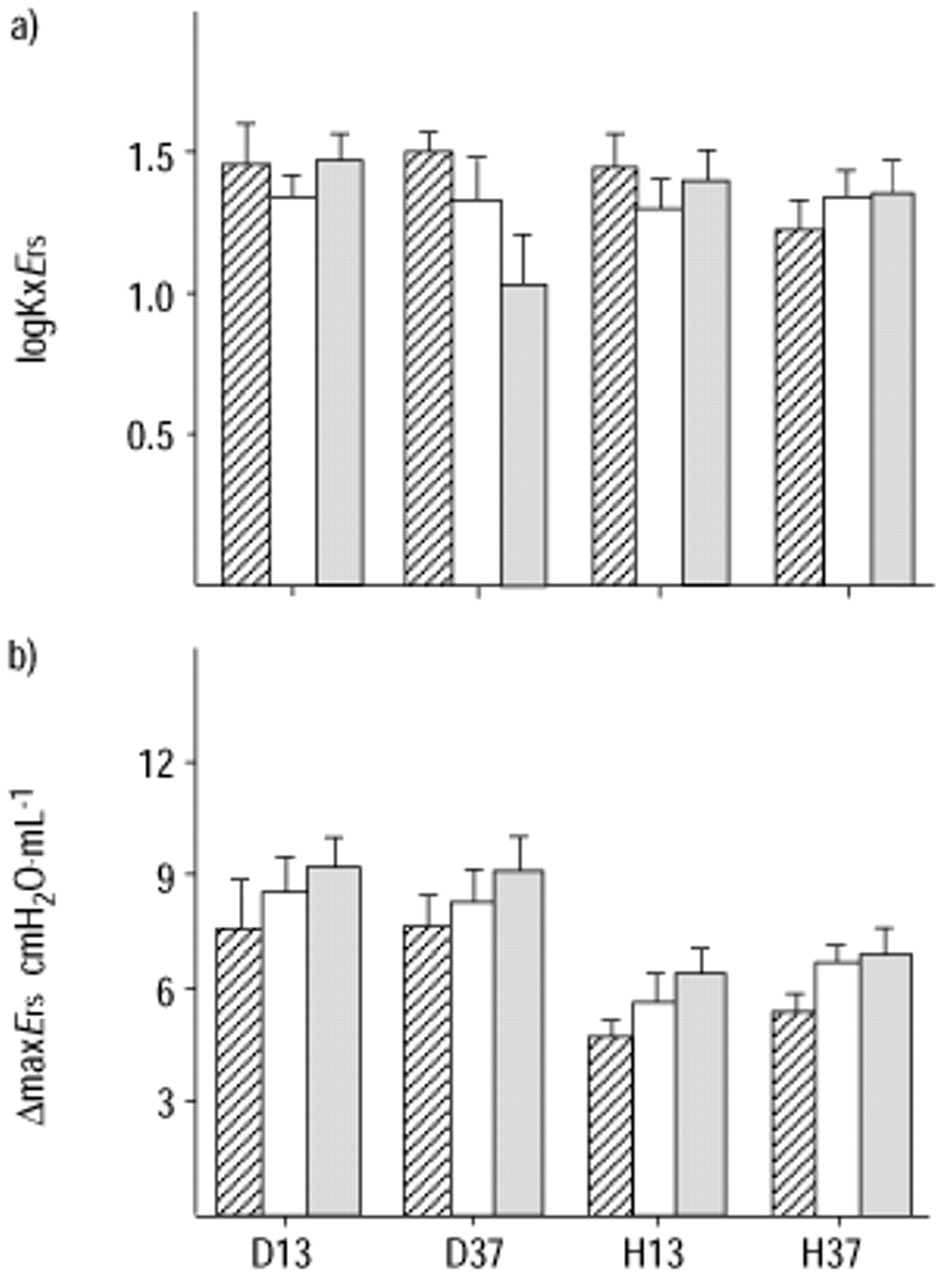
No significant influence of the temperature of inspired air on Δmax values for either Rrs or Ers was observed (figs. 1 and 2⇓⇓). In contrast, values of ΔmaxErs in the groups ventilated with humid air were significantly lower than in the groups ventilated with dry air (p<0.001). Moreover, the decrease in alveolar ventilation resulted in significantly greater values of ΔmaxErs (p<0.001). Statistically significant differences in ΔmaxRrs values were not observed. Finally, there were no differences in Kx values among the groups studied, for both Ers and Rrs (figs. 3⇓ and 4⇓).

Dose/response curves of intravenous methacholine and respiratory system resistance (Rrs) in rats ventilated with a) dry air at 13°C, b) dry air at 37°C, c) humid air at 13°C and d) humid air at 37°C, and with different respiratory frequencies to reach carbon dioxide tension in arterial blood values of 30 (○), 40 (□) or 50 (▵) mmHg. Data are presented as mean±sem.

Dose/response curves of intravenous methacholine and respiratory system elastance (Ers) in rats ventilated with a) dry air at 13°C, b) dry air at 37°C, c) humid air at 13°C and d) humid air at 37°C, and with different respiratory frequencies to reach carbon dioxide tension in arterial blood values of 30 (○), 40 (□) or 50 (▵), mmHg. Data are presented as mean±sem.

a) Values of the logarithm of the dose of intravenous methacholine that caused a 50% maximal increase in respiratory system elastance (logKxErs). b) Values of the maximal increase in respiratory elastance (ΔmaxErs). Animals were ventilated with dry air at 13°C (D13), dry air at 37°C (D37), humid air at 13°C (H13) and humid air at 37°C (H37). Data are presented as mean±sem. └: 30 mmHg; □: 40 mmHg; ▓: 50 mmHg.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
a) Values of the logarithm of the dose of intravenous methacholine that caused a 50% maximal increase in respiratory system resistance (logKxRrs) and of the maximal increase in respiratory resistance (ΔmaxRrs). Animals were ventilated with dry air at 13°C (D13), dry air at 37°C (D37), humid air at 13°C (H13) and humid air at 37°C (H37). Data are presented as mean±sem. └: 30 mmHg; □: 40 mmHg; ▓: 50 mmHg.
Discussion
The influence of alveolar ventilation degree, temperature and humidity on pulmonary responsiveness to a bronchoactive agonist has been studied here for the first time in a normal animal. The maximal response to methacholine was significantly reduced in the presence of humid air, particularly in small airways, as suggested by significant differences in elastance but not in resistance. Higher ventilation was also associated with a lower maximal elastance response.
Exercise, hyperventilation and ventilation with cold air cause a transient increase in pulmonary resistance in a substantial number of patients with asthma (40–90%) and in some subjects without history of asthma 10–13. At a fixed level of ventilation, colder and dryer inspired air exacerbates this response. In contrast, ventilation with warmer and more humidified air reduces its severity. There are numerous similarities between exercise- and hyperventilation-induced bronchospasm, such as the time-course of airway narrowing, the degree of evaporative loss of both heat and water and the response to medication 3, 14. Because of these similarities, experimental models of hyperventilation have been used to study the mechanisms of exercise-induced bronchospasm 4, 5, 14. EIB is probably an amplification of the normal response of the airways, since normal subjects show a reduction in forced expiratory volume in one second following high levels of hyperpnoea with cold, dry air 15, 16. It has been shown that various animal species, such as guinea pigs, dogs, rabbits and monkeys, present airway obstruction in several experimental models of hyperpnoea, mainly those using dry air 14, 17. These animal models have been used to study the mechanisms of EIB. However, to the authors' knowledge, previous studies have not investigated the influence of the degree of alveolar ventilation, temperature and humidity on the responsiveness to a bronchoactive agonist, such as methacholine, in a normal animal. These relationships in normal animals are important in experimental situations where there is no change in baseline respiratory mechanics before the infusion of methacholine, as they clarify the influence of temperature, humidity and ventilation on the behaviour of normal airway smooth muscle.
To evaluate the response to methacholine, the dose that caused 50% of the maximal change in both Ers and Rrs was determined as an index of the position of the dose/response curve (sensitivity). The maximal change (reactivity) was also compared 8. It was observed that temperature, humidity and degree of alveolar ventilation did not significantly influence the position of the curve (fig. 3 and 4⇑⇑). However, the maximal response to i.v. methacholine was significantly affected by ventilation and humidity. These effects were more intense in small airways and/or distal airspaces, as the differences in Ers but not Rrs were significant. It has been shown previously that administration of contractile agonists, such as methacholine or acetylcholine, not only results in an increase in airway resistance but also in substantial changes in the mechanical properties of pulmonary parenchyma 18–20. These observations suggest that dryness has a greater effect than low temperature on the pulmonary response to a bronchoconstrictor. Very low temperatures, however, were not used in this study.
A significant influence of the level of alveolar ventilation on the response to methacholine was also observed, with a higher Pa,CO2 associated with a greater maximal response to methacholine. This difference was significant for Ers (fig. 4⇑). This finding is somewhat surprising, since EIB is related to hyperventilation. In contrast with this observation, Stephens and Mitchell 21, studying canine isolated tracheal strips, observed a decrease in the isometric maximum tetanic tension in the presence of high Pa,CO2. However, Stephens and Mitchell 21 exposed the airway smooth muscle to a much higher Pa,CO2 than in this study (110 versus 50 mmHg). In addition, it can be speculated that there are differences between the in vitro and in vivo contractile response of airway smooth muscle due to haemodynamic, humoral and/or nervous influences.
The mechanisms of exercise- and hyperventilation-induced bronchospasm are not completely determined, but are thought to be related to the cooling and/or drying of the airways resulting from hyperpnoea. Following these stimuli, airway obstruction is due to mediator release, sensory nerve activation and/or reactive hyperaemia of the bronchial circulation 16, 22. Large as well as small airways are involved in this response 16, 23.
In the present study alveolar ventilation was changed by increasing or decreasing respiratory frequency, without modifying VT or flow profile of the ventilator, in order to obtain comparable values of Ers and Rrs in all groups of rats. With this approach, a group with a lower Pa,CO2 (20 mmHg) could not be studied, since the higher respiratory frequency used to reach this value resulted in an increase in baseline tracheal pressure (auto-positive end-expiratory pressure). In addition, if exposed to dry air, rats with higher alveolar ventilation presented higher values of basal (premethacholine) Ers and Rrs, perhaps as a result of hyperventilation-induced bronchoconstriction.
Methacholine was infused i.v. to give the same amount of this agonist to all groups of rats. This would not have been possible by aerosol because of the different respiratory frequency of the groups. However, the i.v. infusion of high doses of methacholine has substantial systemic and pulmonary vascular effects, which could theoretically be influenced by pH or Pa,CO2.
There are other limitations to this study. Different rodents, such as guinea pigs, rats and mice, and even different strains of rats or mice have shown substantially different responses to bronchoactive agonists, such as methacholine. However, only Wistar rats were used in this study. Isocapnic conditions in the lung were not maintained and supplemental oxygen was not administered to the groups of rats that were ventilated with low respiratory frequencies. The rats in the group administered 50 mmHg of Pa,CO2 had significantly lower values of Pa,O2, although these values were not low enough to induce substantial changes in pulmonary haemodynamics. The purpose of this study was to reproduce situations of hyperpnoea and hypopnoea, without correction for blood gases. It has been shown previously that carbon dioxide might reduce pulmonary peripheral resistance 24. However, this was probably not the case in this study, since Pa,CO2 levels did not influence baseline resistance and elastance values.
In conclusion, it has been demonstrated in this study that the pulmonary response to methacholine in normal rats is significantly affected by the humidity of inspired air and the level of alveolar ventilation. Ventilation with dry air and lower respiratory frequency resulted in significantly greater maximal values of respiratory system elastance, but not resistance, suggesting that this effect is more intense in the small airways and/or distal airspaces than in larger airways.
Acknowledgments
The authors would like to thank R.W. Mitchell for his critical review of the manuscript.
- Received March 29, 2001.
- Accepted January 16, 2002.
- © ERS Journals Ltd
References









