





Abstract
It has recently been suggested that proteinase inhibitors modulate the fibrotic response in the lung. This study investigated the development of bleomycin-induced pulmonary changes in pallid mice, deficient in serum α1‐proteinase inhibitor, and with a lower elastase inhibitory capacity, and in congenic C57Bl/6J mice.
Male pallid and C57Bl/6J mice received a single intratracheal instillation of either saline or bleomycin. The investigation was carried out by means of biochemical, morphological and morphometrical methods.
In both strains, 21 and 72 h after bleomycin, the lungs showed foci of inflammatory cell infiltration associated with emphysema. Fibrosis developed with time after bleomycin. At 14 days fibrosis affected 23.46±9.48% (mean± sd) and 40.62±13.34% (p<0.01) of the lungs of C57Bl/6J and pallid mice, respectively. Emphysema affected 3.68±3.11% and 12.57±4.13% (p<0.01) of lung in C57Bl/6J and pallid mice, respectively. In C57Bl/6J mice bleomycin increased lung hydroxyproline content by 34% and desmosine content by 44% (p<0.01 for both). In pallid mice these increases were only 21% (p<0.01) and 6%, which may reflect parenchymal loss.
Thus, the lung destructive response (emphysema) and the subsequent proliferative reaction (fibrosis) to bleomycin are potentiated in α1‐proteinase inhibitor deficiency.
This work was supported by a grant from Ministero dell'Universita' e della Ricerca Scientifica e Tecnologica.
Bleomycin, an antineoplastic agent, induces pulmonary fibrosis in man and experimental animals 1–3. In rodents, bleomycin administration induces a strain-dependent pulmonary inflammatory response characterized by leukocyte infiltration, fibroblast proliferation and increase in collagen content 4, 5. This picture is generally considered a representative model of human idiopathic pulmonary fibrosis 6.
Recently it has been reported that serine proteinase inhibitors such as α1‐proteinase inhibitor (α1‐PI) and secretory leukocyte protease inhibitor (SLPI), as well as the synthetic inhibitor of leukocyte elastase ONO-5046, significantly attenuate the fibrotic response to bleomycin in rodents 7–9. In man, inactivation of proteolytic enzymes may also be a critical event in normal repair and, as demonstrated in infants with respiratory distress syndrome, lack of antiprotease activity is associated with chronicity and development of fibrosis 10. Consequently, these studies suggest a significant role for the antiprotease screen not only, as well documented, in the development of pulmonary emphysema but also in themodulation of fibrotic lesions 11, 12. Thus, the investigation into the role of the antiprotease-protease system in models of fibrosis may contribute to the understanding of the mechanism(s) of this disease.
The pallid mouse has recently been described as a model of genetic α1‐PI deficiency 13. The pallid (pa) mutation was first discovered in a wild mouse 14 and was placed on a C57Bl/6J background by repeated crossing. The only difference between the congenic C57Bl/6J pa/pa and a normal C57Bl/6J +/+ is in the pa gene on chromosome 2, which is inherited in an autosomal recessive fashion 15. Serum α1‐PI levels of the pa mice are significantly lower than in C57Bl/6J mice (2.7±0.4 mg·mL−1 versus 4.4±0.3 mg·mL−1, respectively) 16. Histological evidence of emphysema can be detected only in pa mice from 12 months of age onwards 13, 17.
It was thus of interest to investigate the response to bleomycin in 3–4 month-old pa mice in comparison with the congenic C57Bl/6J mice. The latter mice are known to be sensitive to bleomycin, and also to have relatively low values of serum α1‐PI (4.4±0.3 mg·mL−1 compared with 5.9±0.5 mg·mL−1 in Balb/c mice and 5.7±0.5 mg·mL−1 in NMRI mice) 16, 17.
This study was carried out by means of biochemical, morphological, and morphometrical methods. Additionally, since recent work in vitro and/or in vivo indicates a modulating role for the antiprotease-protease system on the activity of some cytokines 18–20, the presence of tumour necrosis factor‐α (TNF‐α), interleukin 1β (IL‐1β), and transforming growth factor β1 (TGF β1) were also investigated using immunohistochemical methods in lung parenchyma of these animals. The importance of these cytokines for the fibrotic process has recently been stressed 21.
Materials and methods
Bleomycin treatment
Male pallid (pa/pa) mice with a C57Bl/6J background and male C57Bl/6J mice of 3–4 months of age were used in this study. Pallid mice were originally obtained from Jackson Laboratory (Bar Harbor, ME, USA). C57Bl/6J mice were originally supplied by Charles River (Calco, Italy) and used as controls. All animal experimentation was conducted in conformity with the “Guiding Principles for Research involving Animals and Human Beings” and was approved by the Local Ethical Committee of the University of Siena.
For each strain of mice, different groups of animals were used. A group received a single intratracheal instillation of 0.1 μg bleomycin (Rhone-Poulenc Rorer, Milano, Italy) in saline solution (50 μL). Another group was instilled intratracheally with the same amount of saline. All intratracheal instillations were performed under ether anaesthesia. Fourteen days later, the animals from all groups were injected with an overdose of pentobarbital sodium and exsanguinated by cutting the abdominal aorta. The lungs were then excised and processed for biochemical analysis and histological examination. An additional three animals from each strain and group were sacrificed at 14 and 21 h, and 3 and 7 days after the treatment, and the lungs processed for conventional light microscopy and immunohistochemistry.
Bronchoalveolar lavage elastase inhibitory capacity
To analyse the elastase inhibitory capacity (EIC) levels in bronchoalveolar lavage (BAL) fluids, ten animals of each strain were used. Mice were anaesthetized with pentobarbital given intraperitoneally and exsanguinated by severing the abdominal aorta. The trachea was isolated and then cannulated with a 20 gauge blunt needle. With the aid of a peristaltic pump (P‐1 Pharmacia Uppsala, Sweden) the lungs were lavaged in situ 3 times with 0.6 mL normal saline. The average fluid recovery was greater than 95%. Cell-free BAL fluids obtained after centrifugation at 600×g for 15 min, were tested for EIC against homologous mouse leukocyte elastase (MLE) 16. MLE was purified as previously described 22 and active-site titrated using Z-Ala-Ala-Pro-azaAla-ONp 23. It was found to be 96% active. EIC was expressed as μg enzyme inhibited/mL BAL.
Lung biochemical assays
To quantify lung collagen and elastin, animals fromeach group were anaesthetized with sodium pentobarbital and sacrificed by severing the abdominal aorta. After thoracotomy, the thoracic viscera were immediately removed. The lungs were weighed, homogenised (1:5, weight:volume (w:v)) and hydrolysed in 6 N HCl before biochemical determinations. Hydroxyproline (HO‐proline) was determined in lung hydrolysates according to the method of Kivirikko et al. 24 and data were expressed as μg·lung−1.
Desmosine was analysed on hydrolysates by means of an enzyme-linked immunosorbent assay (ELISA) essentially according to Cocci et al. 25. Briefly, rabbit antiserum to desmosine-haemocyanin conjugate (AbI) (Elastin Product Company, Inc, Owensville, MO, USA) was incubated with desmosine standard (0–30 ng) (Elastin Product Company, Inc) or with adequately diluted hydrolysates for 16 h at room temperature. At the same time, microtitre plates (Sigma, St Louis, MO, USA) were incubated with 0.5 μg of desmosine-albumin conjugate (Elastin Product Company, Inc) in 0.05 M sodium carbonate buffer pH 9.6 at 4°C. After incubation, wells were washed five times with 0.05% Tween 20 in 0.15 M phosphate buffered saline (PBS), pH 7.2 and saturated with 0.05% Tween 20 in 0.15 M PBS, 1% bovine serum albumin (BSA) pH 7.2 for 1 h at room temperature. Eight fold aliquots of AbI-standard or AbI-sample solutions were then added tothe wells for a 2 h incubation at room temperature. Wells were then incubated in succession with anti-rabbit immunoglobulin G (IgG) (1:2000) (Sigma) for2 h at room temperature and with peroxidase-antiperoxidase complex (1:200) (Sigma) for 1 h at room temperature. Then, 2,2′-Azino-bis (3‐ethyl-benz-thiazoline‐6‐sulphonic acid) solution (Sigma) was added to the wells. After incubation for 1h at room temperature, absorbance was read at 405 nm. Data were expressed as μg·lung−1.
Morphology and morphometry
The lungs from the different groups of mice were fixed intratracheally with buffered formalin (5%) at a constant pressure of 20 cm H2O at least for 24 h. All lungs were then dehydrated, cleared in toluene, and embedded in paraffin. Transversal sections (7 μm) of each pair of lungs were cut and stained with haematoxylin and eosin. Morphometric assessment consisted of the determination by point counting, of the per cent volume densities of fibrosis Vv(f) and of theemphysematous changes Vv(e) according to the stereological principle of Glagoleff and Weibel 26: Vv=Pp, where Vv is the volume density and Pp the fraction of points superimposed a defined structural change. Point counting was performed at a magnification of ×100, by determining 20 random fields per slide and using a multipurpose grid to count 45 points per field for a total of 900 points per slide. Fibrosis was defined as inflammatory cell infiltration within the alveolar septa and alveolar spaces with deposition of extracellular matrix. Emphysematous changes were defined as abnormal enlargement of air spaces with loss of alveolar septa, with or without thickening of the alveolar walls. The reproducibility of the results was greater than 98%.
Immunohistochemistry
The lungs from the different groups of mice were fixed intratracheally with buffered formalin (5%) at a constant pressure of 20 cm H2O for 4 h, and then immersed in the same fixative for an additional hour. The samples were then dehydrated, cleared in toluene, and embedded in paraffin. Tissue sections (8 μm) were stained for TNF‐α, IL‐1β and TGF β1 by an immunoperoxidase method. The sections were pretreated with 0.3% hydrogen peroxide to inhibit the activity of the endogenous peroxidase. For TGF β1 detection, the sections were digested with 1 mg·mL−1 of hyaluronidase in 0.1 M sodium acetate in 0.15 M NaCl (pH 5.5) for 30 min at 37°C. For TNF‐α and IL‐1β detection, the sections were treated with 0.1% trypsin in 0.05 M tris-HCl buffer, pH 7.6 for 10 min at 37°C. All the sections were also incubated with 3% bovine serum albumin for 30 min at room temperature to block nonspecific antibody binding. They were then incubated overnight at 4°C with the primary antibodies. The primary antibodies used were: rabbit polyclonal antibodies to mouse TNF‐α diluted 1:20 (Immunogenetics, Ghent, Belgium), rabbit polyclonal antibodies to mouse IL‐1β, diluted 1:100 (Antigenix America Inc., New York, N.Y. USA), and rabbit polyclonal antibodies to mouse TGF β1 diluted 1:20 (Insight Biotechnology Ltd., Wembley, England). All the sections were rinsed and incubated with sheep anti-rabbit IgG antibodies for 30 min at room temperature. The staining was revealed by adding peroxidase-antiperoxidase complex prepared from rabbit serum. Detection was accomplished by incubating in diamino-benzidine freshly dissolved in 0.03% hydrogen peroxide in 100 mM Tris-HCl pH 7.4. As negative controls for the immunostaining, the primary antibodies were replaced by nonimmunized rabbit serum.
Statistical analysis
For each parameter, both measured and calculated, mean±sd values for individual animals were calculated. The significance of the differences between groupswas calculated using one-way analysis of variance (ANOVA) (F‐test) 27. Ap‐value of ≤0.05 was considered significant.
Results
Mortality
No animal treated with saline died. Following bleomycin, 25% of the C57Bl/6J mice and 31% of pallid mice died. This difference was not statistically significant. Death occurred 7–14 days after the treatment and appeared to be due to respiratory failure.
Elastase inhibitory capacity in bronchoalveolar lavage
The EIC determined as μg of MLE inhibited by 1 mL of BAL was 7.48±1.48 (mean±sd) in C57Bl/6J mice (n=10). The EIC in pallid mice (n=10) was significantly lower (4.91±0.96, p<0.01).
Lung biochemical assays
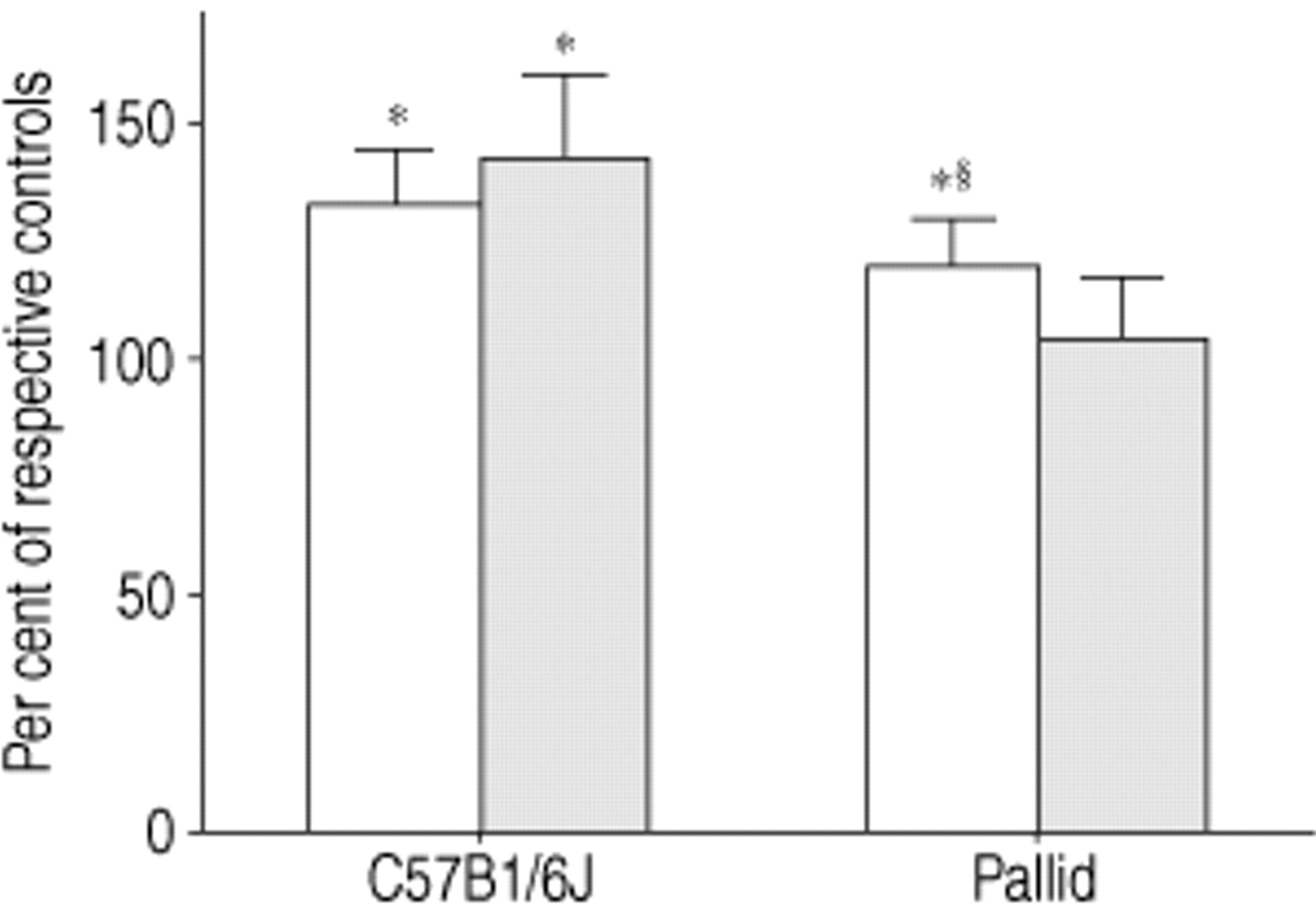
The results of the HO‐proline assessment are shown in figure 1⇓. Fourteen days after bleomycin challenge, the lungs of the C57Bl/6J mice (n=7) showed on average a 34% (p<0.01) higher content in HO‐proline than the lungs of the saline-treated mice (n=7) (203.88±16.26 μg·lung−1versus152.23±12.18 μg·lung−1). Similarly, the lungs of the pallid mice (n=10) showed after bleomycin a mean content in HO‐proline which was 21% (p<0.01) greater than that of the saline-treated mice (n=10) (180.36±14.46 μg·lung−1 versus 149.01±12.02 μg·lung−1).

Lung HO‐proline (□) and desmosine (▒) content in C57Bl/6J (n=7) and pallid (n=10) mice 14 days after bleomycin challenge. Data are given as mean± s.d. and represent per cent change over respective saline-treated controls. *: p<0.01 versus respective saline-treated group. §: p<0.01 versus C57Bl/6J plus bleomycin group.
Figure 1⇑ also shows the results of the lung desmosine content. Following bleomycin, the lungs of the C57Bl/6J mice (n=7) showed a mean desmosine content which was 44% higher than that of the corresponding saline-treated mice (n=7) (3.62±0.52 μg·lung−1 versus 2.52±0.29 μg·lung−1).
The lung desmosine content of the pallid mice had similar values in the bleomycin (2.61±0.41 μg·lung−1) (n=10) and in the saline (2.46±0.31 μg·lung−1) (n=10) groups.
Morphology and morphometry
Eight out of ten C57BL/6J control mice showed a normal, well fixed lung parenchyma (fig. 2⇓), while 2/10 mice had a few patchy areas of trivial airspace enlargement with a mean Vv(e) group value of 0.39% (table 1⇓). Fourteen days after bleomycin, the lungs of all C57BL/6J mice (n=9) showed large areas of cellular infiltration and fibrosis which involved 23.46% of the lungs. Additionally, in 8/9 mice some emphysematous changes were also seen. They were situated predominantly adjacent to the fibrotic lesions (fig. 2⇓), rarely within the areas of fibrosis. These lesions affected, on average, 3.68% of the lungs (table 1⇓).

A) Histologic section from the lung of a C57Bl/6J mouse treated with saline showing a normal parenchyma. B) Lung of a C57Bl/6J mouse treated with bleomycin showing intraseptal and intra-alveolar infiltration of inflammatory cells with deposition of extracellular matrix (arrowheads). Adjacent to these lesions, areas of emphysema-like changes can be seen (asterisks). C) Representative histologic section of a pallid mouse treated with saline showing a patchy area of trivial enlargement of alveolar ducts and sacs. D) Lung of a pallid mouse after bleomycin showing extensive areas of inflammation and fibrosis (arrowheads) intermixed with severe emphysematous changes (asterisks). Haematoxylin-eosin stain (internal scale bar=35 µm).
Lung volume density of fibrotic and emphysematous changes after either saline or bleomycin in congenic strains of mice
In the pallid control group, 4/7 mice showed a few trivial foci of fibrosis which involved 0.59% of the lungs (table 1⇑), while all animals had a few patchy areas ofairspace enlargement sometimes accompanied by flattening of the alveolar septa. These lesions involved usually only 2–3 alveolar sacs (fig. 2⇑) and affected 2.47% of the lungs (table 1⇑). Fourteen days after bleomycin, pallid mice (n=6) responded with the most severe fibrotic and emphysematous changes. Both lesions were seen in all animals, were widely spread and intermixed (fig. 2⇑), with mean values for Vv(f) of 40.62%, and for Vv(e) of 12.57% (table 1⇑).
Immunohistochemistry
Staining for TNF‐α showed, in control mice of both strains, a faint reaction on alveolar cells with morphological features of macrophages and type II pneumocytes. After bleomycin in both strains, a clear reaction was seen on the above mentioned alveolar cells as wellon parenchymal areas showing an inflammatory reaction, at 14 h, 21 h and 3 days. At 7 days, the positive staining was also distributed on extracellular matrix components associated with an inflammatory reaction (data not shown).
In control mice, reaction for IL‐1β revealed a faint and diffuse staining of all alveolar structures. In both mouse strains 14 h after bleomycin, there was a marked positive staining on alveolar cells in the subpleural regions. Subpleural inflammatory reaction and extracellular matrix components also stained positively at 21 h and 3 days. A weaker staining was seen on the above mentioned structures at 7 days (data not shown).
In control animals of both strains, there was practically no reaction to TGF β. Twenty-one hours after bleomycin, a first faint positive staining was seen on pleural cells of both strains. At 3 days, the positive reaction had spread throughout the parenchyma with more intensive staining of alveolar wall cells at 7 days (data not shown).
Thus, the present results do not reveal apparent differences in distribution and stain intensity for the three cytokines, in both strains of mice after bleomycin.
At 21 and 72 h after saline the lungs of both C57Bl/6J and pallid mice which were used for immunohistochemical studies were normal when viewed at light microscopy after haematoxylin and eosin staining. At 21 and 72 hours after bleomycin, the lungs of the C57Bl/6J and of pallid mice showed foci of inflammatory cells associated with appreciable emphysema (fig. 3⇓).

{kind=link}
{kind=link}
{kind=link}
A) Lung of a C57Bl/6J mouse 21 h after bleomycin showing inflammatory cells scattered through lung parenchyma with areas of appreciable emphysema (asterisks). B) Similar histological features were also seen in the lungs of pallid mice earlyafter bleomycin. Haematoxylin-eosin stain. (internal scale bar=50 µm).
Discussion
The main findings of this work are: (1) the influence of a deficiency in α1‐PI on the pulmonary response to bleomycin in 2 congenic strains of mice (2) and the fact that one noxa (bleomycin) can induce two different lung pathologies such as fibrosis and emphysema.
With regard to the first point, it is known that the response to bleomycin is strain-dependent 5, and thus may be related to many independent factors specific to a given strain. These include variations in collagen synthesis and deposition 5, cytokine production 28, H‐2 aplotype as well as non-H‐2 genetic factors 29. Consequently, it was important to investigate the outcome of the bleomycin-lesion within the same strain in two congenic mice that differ in their α1‐PI serum levels. The time point 14 days after bleomycin was chosen following the results of a pilot study in which we investigated the animals at 14 and 35 days after treatment. The 35 day period was then omitted due to the very high mortality observed at this time period (∼70% in both strains). Thus, the few surviving animals could not be considered a representative sample.
It has previously been reported that the pallid mice, compared to the mice of the parent C57Bl/6J strain, have a deficiency in serum α1‐PI levels which correlates with a deficit in serum EIC 16. The present study investigated the EIC in the BAL (i.e. in the target organ) of these 2 strains. The data obtained indicate that the values of the EIC in pallid mice were also significantly lower in BAL.
In both these strains, the morphological response to bleomycin in terms of fibrosis was very similar and consisted of intraseptal and intra-alveolar leukocyte infiltration and mesenchymal cell proliferation with deposition of connective tissue components. Additionally, foci of mononuclear cells were seen. However, morphometrically, the extent of the lesion was almost twice as great in the pallid than in the C57Bl/6J mice. Similarly, the extent of the emphysematous changes was also more than three times greater in the pallid than in the C57Bl/6J mice. Thus, in the absence of differences in genetic background between these two strains, a greater deficit in EIC corresponded to a potentiation of the responses to bleomycin.
The present results, coupled with the observations that exogenous administration of α1‐PI, SLPI, and ofthe compound ONO-5046, a specific inhibitor of neutrophil elastase, significantly attenuates the response to bleomycin 7–9, indicate a role for the ratio of α1‐PI to neutrophil elastase in the development of the bleomycin-fibrosis.
In the clinics, an imbalance in the ratio of α1‐PI to neutrophil elastase is known to play a major role in the development of pulmonary emphysema 11, 12. Its rolein the development of fibrosis is the subject of investigation. Recently, in some patients with idiopathic pulmonary fibrosis, the ratio in blood of neutrophil elastase to α1‐PI was significantly greater than that seen in patients with emphysema and in normal individuals 30.
The mechanism of action of neutrophil elastase in the development of fibrosis could rest on its capability ofattacking a number of connective tissue proteins leading to severe tissue changes and chronic inflammation 31, 32. Alternatively, or in addition to these direct effects, neutrophil elastase may exert some indirect effects consisting of the modulation of certain cytokines. In particular, elastase has been considered to modulate the in vivo effects of TNF‐α, and pretreatment with α1‐PI protects mice against a lethal dose of this cytokine 18, 19. Also, it has been recently reported that the mouse elastase inhibitor ONO-5046 33 significantly decreased messenger ribonucleic acid (mRNA) levels of inflammatory and matrix producing cytokines in BAL cells after bleomycin 9. Indeed, under the present study's experimental conditions, no difference could be detected either in distribution or in intensity for TNF‐α, IL‐1β and TGF‐β, at various time-intervals after bleomycin, between the 2 strains of mice. The overall results of the present study, coupled with data in the literature, indicate that a deficiency in α1‐PI also plays a role in the pathogenesis of fibrosis.
With regard to the second point (one noxa, two pathologies), emphysematous lesions adjacent to the area of fibrosis were found 14 days after bleomycin in C57Bl/6J mice which, in comparison to mice of other strains, also have a mild deficiency (approximately -25%) in serum EIC 17. These emphysema-like lesions have been described previously in C57Bl/6J mice following bleomycin 34, 35, but have received little, if any, attention. They consist of enlargement of air spaces accompanied by flattening and/or disappearance of the alveolar septa. At 14 days after bleomycin, alveolar walls are sometimes thickened, thus these lesions do not fully meet the official definition of emphysema that excludes “obvious fibrosis” 36. They are thus called here “emphysematous”. Untreated, normal C57Bl/6J mice do not develop spontaneous emphysema. However, intratracheal administration of either N‐formyl‐L‐methionyl‐L‐leucyl‐L‐phenylalanine (FMLP) or endotoxin, results in a rapid influx of neutrophils into the lungs and emphysema 37, 38. Strains of mice such as the NMRI, which do not have a deficiency in α1‐PI, respond to FMLP with a similar influx of neutrophils into the lungs, but not with emphysema 37. Thus, the emphysematous changes observed here in the C57Bl/6J mice, probably represent a primary lesion, i.e. the result of a proteolytic attack from infiltrating inflammatory cells, which follow the instillation of bleomycin. This assumption is based on the observation of areas of “true” emphysema associated with foci of inflammation early after bleomycin (at 21 and 72 h) and thus, long before the development of fibrosis.
Biochemically, in the C57Bl/6J mice, there was an increase in lung content of HO‐proline, which reflects the fibrotic changes 5, accompanied by an increase in lung content of desmosine, which is consistent with the increase in lung elastin mRNA expression found at various time-intervals after bleomycin 34.
In pallid mice, 21 and 72 h after bleomycin, appreciable emphysema was seen associated with spotty areas of inflammatory cells infiltration. At 14 days, both the fibrotic and the emphysematous lesions were more severe than in the C57Bl/6J mice and appeared intermixed. Thus, although at this later time the emphysematous lesions of the pallid mice appear morphologically to be of the paracicatricial type, the early observation indicates that their development preceded that of the fibrosis. A further deterioration of the air space enlargement due to the retraction forces of the fibrotic process however, cannot be excluded.
The large areas of parenchymal destruction seen in these mice resulted in a lower increase in lung HO‐proline than in the C57Bl/6J mice and in a trivial increase in desmosine.
All these data show that one noxa, bleomycin, can induce in the same lung two distinctly different types of responses: the destructive emphysematous type and theproliferative fibrotic type. The present results extend previous findings obtained with cadmium chloride, asubstance that when given alone intratracheally to hamsters induces a lung fibrotic lesion, and when given together with the lathyrogen, β‐aminoproprionitrile, induces emphysema 39. The hamster, relative to several species, also has an inherently low serum antiprotease activity 40. Taken all together, these results suggest that the outcome of the lung response in terms of fibrosis and emphysema, may be conditioned by the intrinsic levels of antiprotease activity and that the two lesions may coexist in presence of an α1‐PI deficiency.
- Received February 2, 2000.
- Accepted September 25, 2000.
- © ERS Journals Ltd
References









