





Abstract
We hypothesised that primary bronchial epithelial cells (PBECs) from subjects with chronic obstructive pulmonary disease (COPD) respond differently to Pseudomonas aeruginosa lipopolysaccharide (LPS) after cigarette smoke extract (CSE) exposure than PBECs obtained from smokers without airflow obstruction and nonsmokers.
PBECs from 16 COPD subjects, 10 smokers without airflow obstruction and nine nonsmokers were cultured at air–liquid interface. Cultures were incubated with CSE prior to stimulation with P. aeruginosa LPS. Interleukin (IL)-6 and IL-8 were measured by ELISA and Toll-like receptor (TLR)-4 expression by fluorescence-activated cell sorter. Activation of nuclear factor (NF)-κB was determined by Western blotting and ELISA, and MAPK and caspase-3 activity by Western blotting. Apoptosis was evaluated using Annexin-V staining and the terminal transferase-mediated dUTP nick end-labelling methods.
Constitutive release of IL-8 and IL-6 was greatest from the COPD cultures. However, CSE pretreatment followed by P. aeruginosa LPS stimulation reduced IL-8 release from COPD PBECs, but increased it from cells of smokers without airflow obstruction and nonsmokers. TLR-4 expression, MAPK and NF-κB activation in COPD cultures were reduced after CSE treatment, but not in the smokers without airflow obstruction or nonsmoker groups, which was associated with increased apoptosis.
CSE attenuates inflammatory responses to LPS in cells from people with COPD but not those from nonsmoking individuals and those who smoke without airflow obstruction.
- Airway inflammation
- cigarette smoking
- chronic obstructive pulmonary disease
- epithelial cell biology
- infections in COPD
- smoking and health
Chronic obstructive pulmonary disease (COPD) is a global health problem and a major cause of death [1]. Exacerbations of COPD secondary to infection are a significant cause of morbidity, mortality and burden on healthcare costs. The relationship between smoking, infection, inflammation and COPD is not well understood.
Interleukin (IL)-6 and IL-8 responses of primary bronchial epithelial cells (PBECs) to stimulation with cigarette smoke extract (CSE) have been of particular interest [2]. IL-8 is an important neutrophil chemoattractant and has been implicated in inflammatory lung disease [3], and is often used as a measure of inflammation in cell culture research [4]. Cigarette smoke induces IL-6 in lung tissue [5] and levels relate to COPD disease severity [6]. Systemically, IL-6 induces skeletal muscle wasting [7] and probably heightens exacerbation risk [8], both of which are major comorbidities in COPD.
Nuclear factor (NF)-κB is an important transcriptional factor in the regulation of inflammatory genes in the airways of people with COPD [9]. Previous investigators have reported a reduced activation of NF-κB in PBECs after exposure to CSE [10], while others have reported increased expression in bronchial biopsies obtained from smokers and COPD subjects [9]. Activated NF-κB can have pro-apoptotic effects in certain cells, yet be protective of apoptosis in others [11]. CSE has been reported to induce apoptosis in PBECs [12] and primary nasal epithelial cells by some [13], but not by other investigators [10]. Furthermore, recent research using PBECs indicated that CSE induces necrosis rather than apoptosis [14].
CSE provides a tool to explore the impact cigarette smoke has on PBEC cultures and facilitates our understanding of crucial intracellular signalling pathways. There is no consensus on the capacity for CSE to induce a pro-inflammatory response in epithelial cells. There is considerable diversity in the methods adopted to prepare CSE and no agreed gold standard. However, the weight of evidence would support that CSE has predominantly pro-inflammatory [2, 15, 16] as opposed to immunosuppressive effects [17, 18] on bronchial epithelial cells. Although there is evidence that nasal epithelial cells can act as a satisfactory substitute for bronchial epithelial cells for certain end-points, we have recently shown that bronchial epithelial cells respond differently to nasal epithelial cells to CSE treatment [19].
In this study we investigated the responses of PBECs obtained from subjects with COPD, smokers with no airflow obstruction and nonsmoking control subjects to CSE treatment. To address this, we cultured cells at the air–liquid interface (ALI), which most closely resembles the in vivo environment, and stimulated cells with Pseudomonas aeruginosa lipopolysaccharide (LPS) as a surrogate for infection, with or without pretreatment with CSE. We aimed to determine the effects of CSE on inflammatory responses to P. aeruginosa LPS and levels of apoptosis in COPD epithelial cells compared to smokers without airflow obstruction and nonsmokers.
METHODS
Study subjects
16 subjects with a diagnosis of COPD according to the British Thoracic Society guidelines (13 of whom were current smokers, three ex-smokers), 10 smokers without airflow obstruction and nine nonsmokers were included. Subject demographics are detailed in table 1. All study subjects provided written informed consent to have a fibre-optic bronchoscopy, during which between four and six bronchial brushings obtained under direct visual guidance by means of a protected brush were obtained from the third generation bronchus. Brushings were placed in bronchial epithelium growth medium and transported to the laboratory for culture. Exclusion criteria included other significant pulmonary pathologies, including lung carcinoma. None of the patients had taken oral corticosteroids for at least 8 weeks prior to the study and no individual had a bronchodilator reversibility of greater than 10%. Patients with COPD had no recent exacerbations and their current treatments are shown in table 1. The 10 smokers without airflow obstruction required a bronchoscopy for clinical requirements such as persisting haemoptysis, but no abnormalities were evident during the procedure. The nine nonsmokers took part on a voluntary basis. This study was approved by the office for research ethics committees, Northern Ireland (REC: 09/NIR03/42).
Cell culture
PBECs were initially expanded in bronchial epithelial growth medium (BEGM; Promocell, Heidelberg, Germany) in purified bovine collagen coated (PureCol; Advanced Biomatrix, San Diego, CA, USA) T10 flasks and then T75 flasks for expansion with penicillin streptomycin antibiotics (Invitrogen, Grand Island, NY, USA) and Primocin (Invivogen, San Diego, CA, USA). Cells were confirmed to be epithelial in origin by randomly staining cultures by immunocytochemical staining for cytokeratin expression (data not shown). All experiments were performed in ALI cultures by seeding cells onto collagen coated Transwells (Corning Inc., Corning, NY, USA) at a seeding density of 1.5×105 cells per well and grown until cultures became confluent and developed tight junctions. At this stage the apical media was removed and the cell cultures fed basolaterally only on alternate days for 28 days.
CSE
CSE was prepared by a modification of the method of Richter et al. [15]. One commercial Marlboro Red cigarette (0.8 mg nicotine; 10 mg tar; 10 mg carbon monoxide) was combusted with a modified syringe-driven apparatus. The smoke was bubbled through 25 mL of media over 5 min by drawing 35-mL volume of smoke every 15 s. The resulting suspension was filtered through a 0.2 μm pore-size filter to remove large particles and bacteria. This solution was regarded as “100% CSE” and was freshly generated for each experiment, and subsequently serially diluted with culture medium to obtain a final 5% working concentration.
Immunofluoresence
Selected cultures were stained with a rabbit anti-E-cadherin primary antibody to demonstrate the presence of tight junctions. Separate cultures were exposed to both a 1:200 dilution of rabbit anti-MUC5AC primary antibody and a mouse anti-acetylated α-tubulin antibody to demonstrate the presence of goblet cells and cilia, respectively. Images were captured and visualised using LAS AF (Leica, Wetzlar, Germany) acquisition software. Full details are provided in the online supplementary material.
FACS
Toll-like receptor (TLR)-4 was determined using fluorescence-activated cell sorter (FACS) by staining permeabilised cells with phycoerthrin-conjugated anti-TLR-4 monoclonal antibodies (eBioscience, San Diego, CA, USA). The results obtained with specific antibodies were compared with those using isotype-matched control antibodies. Analysis of 10 000 events was performed using an Epics XL flow cytometer (Beckman Coulter, UK Ltd, High Wycombe, UK).
Apoptosis was analysed using Annexin V and propidium iodide (PI) staining (eBioscience, UK). For experiments using CSE, cells were treated with 5% CSE for 24 h and the amount of apoptosis and/or necrosis was measured. Events which were positive for Annexin V, but negative for PI were considered to be early apoptotic, events positive for both Annexin V and PI late apoptotic, and events positive for PI alone necrotic. Full details are provided in the online supplementary material.
TUNEL assay
After treating PBECs with CSE for 24 h, cultures were analysed for apoptosis using the Click-It TUNEL assay (Invitrogen, UK). Cells, after being fixed and permeabilised, were exposed to a reaction cocktail overnight at room temperature. Cells were then treated with a reaction buffer additive mixture for 30 min, and cells mounted onto glass coverslips with mounting media and DAPI. 10 randomly selected high power fields at a final magnification of ×100 were counted and the results expressed as the total number of apoptotic cells divided by the total number of cells per field. Full details are provided in the online supplementary material.
Western blots
Western blotting was used to determine phosphorylation of p38, ERK and JNK1 MAPK, cleaved caspase 3, IκB-α and phospho-NF-κB after 4 h stimulation with P. aeruginosa LPS, with or without 5% CSE pretreatment for 24 h. Full details are provided in the online supplementary material.
ELISA
Cytokine concentrations were measured from aliquots of basolateral and apical culture medium after appropriate stimulation as outlined in the results section using commercial IL-6 and IL-8 ELISA kits (R&D Systems Europe, Abingdon, UK) according to the manufacturer's instructions. A chromogenic endotoxin quantification kit was used for the detection of bacterial endotoxins according to manufacturer's instructions (Thermoscientific, Cambridge, UK).
TransAM NF-κB assay
Nuclear extracts were prepared using a nuclear extraction kit from Active Motif (La Hulpe, Belgium) according to the manufacturer's instructions. Details are provided in the online supplementary material. The Active Motif Trans-AM NF-κB ELISA kit (Active Motif) was used to determine the levels of p65 in nuclear extracts. In brief, 2 μg of nuclear extract, diluted to 20 μL, was added to the wells coated with oligonucleotides containing the NF-κB consensus binding site. The primary antibodies used to detect NF-κB recognise an epitope on p65 that is accessible only when NF-κB is activated and bound to its target DNA. After the addition of secondary antibodies conjugated with HRP and substrate, absorbance was read at 450 nm (with a reference wavelength at 650 nm). In order to monitor for specificity, competitive binding assays were performed. Wild-type or mutated consensus oligonucleotides were added to the wells containing immobilised oligonucleotides before the addition of nuclear extracts.
Statistics
Statistical analysis was performed using SPSS version 17.0 (SPSS Inc., Chicago, IL, USA). Data are presented as median values with interquartile ranges. Comparisons between groups were performed using the nonparametric Kruskal–Wallis test for multiple comparisons and the Mann–Whitney test for two groups. A p-value of less than 0.05 was considered significant.
RESULTS
Cell culture
Cells were successfully cultured in Transwells, developed tight junctions and had a cobblestone appearance when examined using a phase contrast light microscope. Mucus secretion was apparent after approximately 7 days in culture. After 14 days, there was an increase in the number of ciliated cells and after 28 days in culture a marked increase in their number was apparent (supplementary figure S1).
CSE
The optical density of a 5% CSE concentration at 450 nm did not vary significantly when comparing a series of 5% CSE preparations (optical density ∼0.25). Preparing CSE using a single cigarette in 25 mL of media produced a sufficiently consistent preparation. Previous investigators have reported similar findings [17]. Endotoxin was not detected in our 5% CSE preparation.
Soluble mediator release
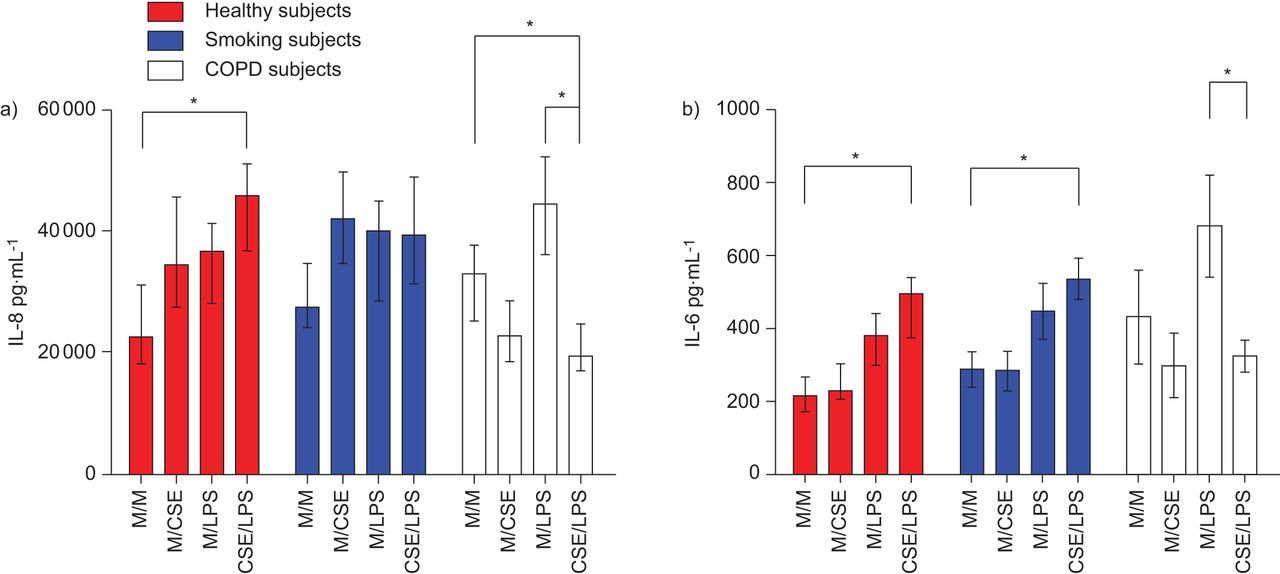
Constitutive and stimulated release of IL-8 and IL-6 was used to determine epithelial cell activation. Protein concentration was not significantly different from Transwells selected at random determined using a BCA assay (data not shown), and so soluble mediator release was expressed in pg·mL−1 rather than normalised to protein concentration. Concentrations of P. aeruginosa LPS (Sigma-Aldrich, Gillingham, UK) higher than 50 μg·mL−1 were cytotoxic and lower concentrations released less IL-8. The release of both IL-8 and IL-6 after stimulation with 50 μg·mL−1 P. aeruginosa LPS from cultures obtained from control subjects and smokers without airflow obstruction was heightened after pretreatment with CSE for 24 h. In the COPD cultures, 5% CSE pretreatment mitigated the stimulatory effects of p. aeruginosa LPS (fig. 1).

a) Interleukin (IL)-8 and b) IL-6 release from apical supernatants in primary bronchial epithelial cell (PBEC) air–liquid interface (ALI) cultures from chronic obstructive pulmonary disease (COPD) subjects, smoking subjects and healthy control subjects after treatment with Pseudomonas aeruginosa lipopolysaccharide (LPS) (50 μg·mL−1) for 24 h, with or without pretreatment with 5% cigarette smoke extract (CSE) for 24 h. Well differentiated PBEC ALI cultures were treated with P. aeruginosa LPS (50 μg·mL−1) for 24 h, with or without pretreatment with 5% CSE (or vehicle) for 24 h (n=7 for each group). Apical supernatants were collected and assessed for IL-8/IL-6 by ELISA. Data are displayed as medians and interquartile ranges. M/M: media (24 h) and media (24 h); M/CSE: media (24 h) and 5% CSE (24 h); M/LPS: media (24 h) and LPS 50 μg·ml−1 (24 h); CSE/LPS: 5% CSE (24 h) and LPS 50 μg·ml−1 (24 h). *: p<0.05.
In separate experiments, cells obtained from a nonsmoking subjects were incubated with various concentrations of CSE (1–50%) for 24 h. CSE stimulated the cultures to release IL-8 up to a 5% concentration, subsequently falling at higher concentrations due to cytotoxicity (fig. 2).

Interleukin (IL)-8 dose response from control primary bronchial epithelial cell (PBEC) air–liquid interface (ALI) cultures after prolonged treatment with cigarette smoke extract (CSE). Well differentiated PBEC ALI cultures from a) nonsmokers and b) chronic obstructive pulmonary disease subjects were treated with 1%, 2%, 5%,10%, 20% or 50% CSE (or vehicle) for 24 h. Supernatants from apical segment were collected and assessed for IL-8 by ELISA (n=5 for each group). Data are displayed as medians and interquartile ranges. *: p<0.05.
CSE induces apoptosis in primary bronchial epithelial cell cultures
Apoptosis was confirmed using the TUNEL assay. For the representative image (fig. 3), the number of apoptotic cells determined by this technique was 4% for the healthy subject and 19% for the corresponding patient with COPD after cells were exposed to CSE for 24 h.

Effect of 5% cigarette smoke extract (CSE) on apoptosis in primary bronchial epithelial cell cultures from healthy subjects and smokers from air–liquid interface cultures determined using the TUNEL assay. Representative images of bronchial epithelial cells from nonsmoking subjects or smokers without airflow obstruction were grown on coverslips and treated with a) PBS or b) DNase I solution for the negative and positive control, respectively. Cultures obtained from c) a nonsmoking subject and from d) a chronic obstructive pulmonary disease subject were treated with 5% CSE for 24 h and the number of apoptotic cells counted using the Click-iT reaction according to manufacturer's instructions.
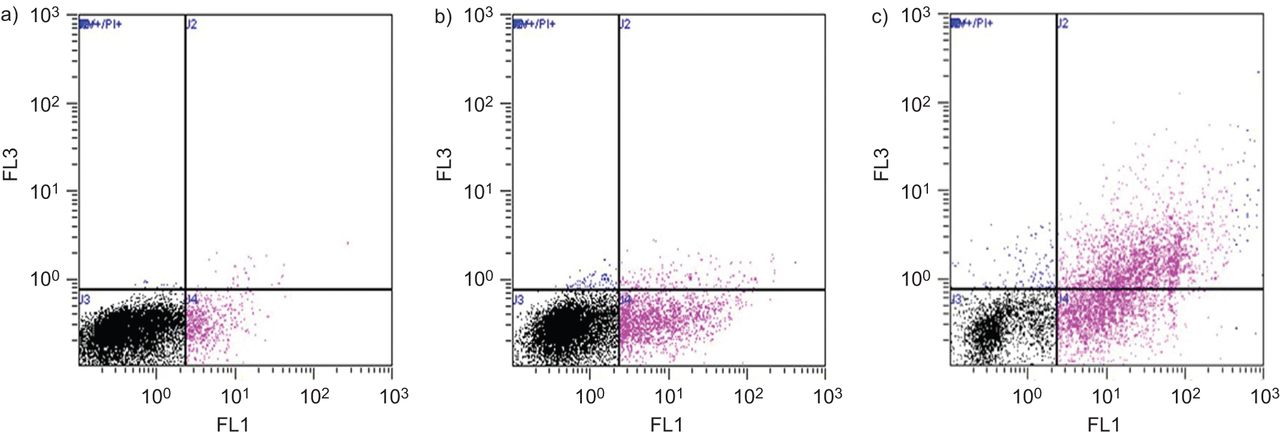
Using the FACS technique, also after treatment with 5% CSE for 24 h in the COPD cells: 51% of cells remained viable (Annexin V and PI negative); 19% were undergoing early apoptosis; 26% undergoing late apoptosis; and 4% were necrotic (fig. 4). In separate experiments, cells from each of the three study groups (again after treatment with CSE for 24 h) were stained with Annexin V to determine any differences in the levels of apoptosis between the groups. CSE induced the highest percentage of apoptosis from the COPD cultures, with the least amount from the nonsmokers. Representative dot plots are shown in figure 5.

Annexin V/propidium iodide (PI) analysis of 5% cigarette smoke extract (CSE) treatment in primary bronchial epithelial cell air–liquid interface chronic obstructive pulmonary disease cultures. In each plot, the horizontal axis represents intensity of staining for annexin V and the vertical axis intensity of staining for PI (determined in the FL1 and FL3 plot, respectively, both logarithmic scale). a) Untreated cells stained with isotope control, b) cells treated with staurosporin 2 μM for 8 h, c) cells treated with 0.1% Triton for 8 h, and d) cells treated with 5% CSE for 24 h. For cells treated with 5% CSE: 51% remained viable; 19% were undergoing early apoptosis; 26% undergoing late apoptosis; and 4% were necrotic.

Annexin V analysis in primary bronchial epithelial cell air–liquid interface chronic obstructive pulmonary disease cultures demonstrating the effects of 24 h incubation with 5% cigarette smoke extract on cell viability. Representative dot-plots demonstrating the percentage of cells staining positive for annexin V. Dot plots correspond to a) nonsmoker, b) smoker without airflow obstruction and c) a chronic obstructive pulmonary disease subject. The percentage of cells staining positive in each quadrant are indicated in table 2.
CSE cleaves full length caspase-3 in PBEC cultures
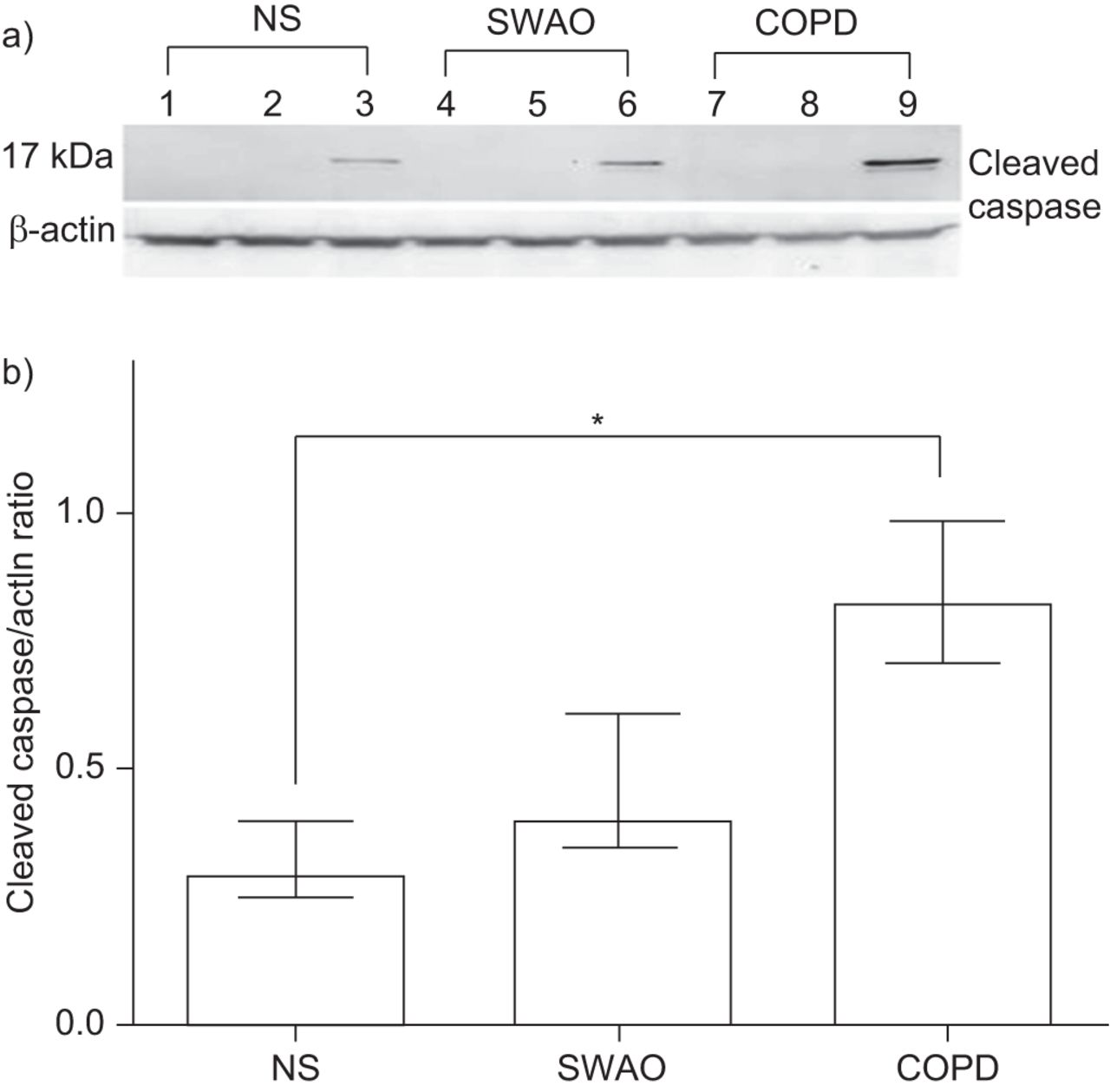
Using an antibody against the cleaved fragment of caspase-3 (17 kDa), we demonstrated by Western blotting that treatment with P. aeruginosa LPS and CSE increased the amount of active caspase-3, whereas P. aeruginosa LPS alone did not. Furthermore, the COPD cultures had the greatest amount of cleaved caspase-3 after P. aeruginosa LPS and CSE stimulation. Cleaved caspase to actin ratio, as determined by densitometry and a representative blot, are shown in figure 6.

Effect of Pseudomonas aeruginosa lipopolysaccharide (LPS) and 5% cigarette smoke extract (CSE) on caspase-3 activation in primary bronchial epithelial cell air–liquid interface (ALI) cultures determined by Western blotting. a) Lanes 1–3 represent treatment of ALI cultures from a nonsmoking (NS) subject with media, 50 μg·mL−1 P. aeruginosa LPS (4 h), and 5% CSE (24 h) followed by 50 μg·mL−1 P. aeruginosa LPS (4 h), respectively. Lanes 4–6 represent equivalent stimulation in ALI cultures obtained from a smoker without airflow obstruction (SWAO) and lanes 7–9 ALI cultures from a subject with chronic obstructive pulmonary disease. The upper blot represents the large fragment of caspase-3 resulting from cleavage (17 kDa) and the lower β-actin. b) Cleaved caspase:actin ratio following exposure to 5% CSE (n=5 for each group). Data are displayed as medians and interquartile ranges. *: p<0.05.
In separate experiments, COPD cultures were treated with increasing concentrations of P. aeruginosa LPS (0–50 μg·mL−1) and CSE (5–50%) and caspase-3 was determined using an antibody against full length caspase (35 kDa) and its cleaved fragment (17 kDa). P. aeruginosa LPS alone did not cleave full length caspase, but there was definite cleavage using a 5% CSE and increasingly so with higher CSE concentrations (supplementary figure S2).
CSE reduces TLR-4 expression in COPD cell cultures
To examine whether CSE regulates the expression of TLR-4, PBEC cultures were treated with P. aeruginosa LPS (50 μg·mL−1), with or without pretreatment with 5% CSE for 24 h. Although there was no change in the absolute values for mean fluorescence intensity for TLR-4 in the cultures from control subjects after CSE treatment, CSE treatment reduced the mean fluorescence intensity for TLR-4 in the COPD cultures. Representative histograms are shown in figure 7.

Representative histogram of amount of intracellular Toll-like receptor (TLR)-4 in primary bronchial epithelial cell (PBEC) air–liquid interface (ALI) cultures after stimulation with Pseudomonas aeruginosa lipopolysaccharide (LPS) with or without 5% cigarette smoke extract (CSE) pretreatment. Well differentiated PBEC ALI cultures from a) and b) a subject with chronic obstructive pulmonary disease (COPD), and cultures from c) and d) a smoker without airflow obstruction were incubated with 5% CSE for 24 h and then stimulated with P. aeruginosa LPS (50 μg·mL−1) for 4 h. e) and f) Separate PBEC ALI cultures from a COPD subject were treated either with or without a 5% CSE alone for 24 h. Cells were then fixed, permeabilised and stained with a phycoerthrin-conjugated antibody against TLR-4 or equivalent isotype-matched control. Mean fluorescence intensity (MFI) is shown (n=4 for each group). Data are displayed as medians and interquartile ranges. *: p<0.05.
CSE reduces MAPK activation and NF-κB in COPD cell cultures
Western blotting showed a reduced phosphorylation of p38, JNK and ERK MAPK in the COPD cultures after treatment with 5% CSE for 24 h. All MAPK were activated, even without stimulation, with no further significant increase after P. aeruginosa LPS stimulation (lanes 1–4 in all blots; supplementary figure S3–S5). However, levels of phosphorylated p38, ERK and JNK were reduced with 24 h 5% CSE pretreatment and P. aeruginosa LPS stimulation in the COPD cultures (lanes 5–8 in all blots; supplementary figure S3–S5). The reduced activation of MAPK was not evident in the control cultures. The reduced activation was not as pronounced for p38 compared with JNK or ERK, and so this was also measured using FACS which confirmed a reduced mean fluorescence intensity for phospho-p38 after 5% CSE treatment (supplementary figure S3).
There was a significant increase phospho-NF-κB protein level from whole cell lysates after stimulation with 50 μg·mL−1 P. aeruginosa LPS for all three groups. This was further heightened in the nonsmoker and smokers without airflow obstruction groups after pretreatment with 5% CSE. In contrast, in the COPD cultures, pretreatment with 5% CSE reduced P. aeruginosa LPS induced NF-κB activation. Phospho-NF-κB to actin ratio as determined by densitometry and a representative blot are shown in figure 8. Levels of IκB-α decreased as phospho-NF-κB increased. Repeating the experiments and determining levels of p65 NF-κB from nuclear extracts using an ELISA based method demonstrated similar results (fig. 9).

Effect of lipopolysaccharide (LPS) with or without 5% cigarette smoke extract (CSE) on nuclear factor (NF)-κB activation in primary bronchial epithelial cell (PBEC) air–liquid interface (ALI) cultures determined by Western blotting. a) Western blot of p65 NF-κB and IκB-α protein expression (with β-actin loading control) in PBEC ALI cultures of control nonsmokers (NS), smokers without airflow obstruction (SWAO) and chronic obstructive pulmonary disease (COPD) patients after treatment with 50 μg·mL−1 Pseudomonas aeruginosa LPS for 4 h or 5% CSE treatment for 24 h followed by treatment with 50 μg·mL−1 P. aeruginosa LPS for 4 h. Lanes 1–3 represent treatment of nonsmoker PBEC's with media alone, 50 μg·mL−1 P. aeruginosa LPS (4 h) and 50 μg·mL−1 P. aeruginosa LPS (4 h) with 5% CSE pretreatment (24 h). Lanes 4–6 represent equivalent treatments in an ALI culture from a smoker without airflow obstruction and 7–9 in those from a COPD subject. b) Phospho-NF-κB to actin ratio (n=5 for each group). Data are displayed as medians and interquartile ranges. *: p<0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of lipopolysaccharide (LPS) with or without 5% cigarette smoke extract (CSE) on nuclear factor (NF)-κB activation in primary bronchial epithelial cell (PBEC) air–liquid interface (ALI) cultures determined using the TransAM NF-κB kit. Nuclear extracts from PBEC ALI cultures of control nonsmokers (NS), smokers without airflow obstruction (SWAO) and chronic obstructive pulmonary disease (COPD) patients after treatment with 50 μg·mL−1 Pseudomonas aeruginosa LPS for 4 h or 5% CSE treatment for 24 h followed by treatment with 50 μg·mL−1 P. aeruginosa LPS for 4 h were assayed for NF-κB p65 activation using the TransAM NF-κB p65 Kit. Results shown are from wells assayed in duplicate.
DISCUSSION
In this series of experiments we have demonstrated that the stimulated release of IL-6 and IL-8 from cultures obtained from controls and smokers without airflow obstruction is amplified by 5% CSE pretreatment, but this was not apparent in the COPD group. Our results also show cellular immunosuppressive effects of 5% CSE in the COPD cultures. These findings suggest that COPD PBECs are more susceptible to the immunosuppressive effects of CSE which may explain, in part, the increased susceptibility of this particular group to respiratory infections.
In our experiments, 5% CSE reduced the expression of TLR-4. Similar findings were recently reported using a human bronchial epithelial cell line, where CSE treatment led to a downregulation of TLR-4 expression, which was associated with a corresponding increase in the release of IL-8. Internalisation of the receptor was proposed as the mechanism, as there was a parallel increase in the expression of TLR-4 in permeabilised cells [4]. However, the reported effects of CSE on TLR-4 expression are not entirely consistent. Although there was a dose dependent downregulation of TLR-4 mRNA and protein expression in A549 cells, after stimulation with CSE [20], a publication the following year suggested that CSE increases TLR-4 expression in BEAS-2B cells [21]. CSE can also affect other important receptors, such as the RIG-1 receptors, by downregulating their surface expression [22]. Therefore, although our data indicate that CSE downregulates TLR-4 expression in COPD PBECs, we cannot be absolutely confident that this is the sole mechanism of its immunosuppressive effects. It is conceivable that CSE also affects other receptors, which may affect cell stimulation independent of TLR-4.
Activation of NF-κB regulates gene expression, promotes cell survival, and serves to protect cells from apoptosis in murine B cells and in a number of cell lines [23, 24]. Furthermore, activation of NF-κB through a TLR-dependent activation by hyaluronan in airway epithelial cells has also been shown to be protective of apoptosis [25]. Interestingly, as the absolute amount of TLR-4 was reduced in the COPD epithelial cell cultures after treatment with CSE, which was associated with a reduction in the activation of all MAPKs, this is a potential mechanism for apoptosis in these particular cultures to CSE treatment. However, other data do not support this concept, as treating PBECs obtained from healthy subjects with CSE for 48 h led to inhibition of NF-κB activation without any evidence of apoptosis, necrosis or caspase-3 activation [10].
As well as differences in soluble mediator release, we have demonstrated differences in phosphorylated NF-κB in our study groups. Although treating PBECs obtained from healthy control subjects and smokers without airway disease with P. aeruginosa LPS increased the amount of phosphorylated NF-κB, this was not the case for the COPD ALI cultures. Furthermore, the COPD ALI cultures had a reduced activation of all MAPK after treatment with 5% CSE. The latter observation was demonstrated in parallel with a reduced expression of TLR-4 and a diminished expression of the transcription factor, phosphorylated NF-κB.
The use of CSE for in vitro studies has been criticised as an unsatisfactory model for the long term, low concentration smoke which smokers are exposed to [26]. Concentrations of CSE used can range from 100% CSE for 15 min [17] to 1% CSE for 24 h (in those studies which use a single cigarette to prepare the initial “100%” stock CSE) [27]. Other investigators have prepared CSE using multiple cigarettes ranging from two cigarettes up to as many as five [13, 28]. Many of these studies assessed soluble mediator release and a single time point, but two studies highlight that caution is required as CSE can delay LPS-induced release of IL-8 and granulocyte–macrophage colony stimulating factor in primary epithelial cells, but yet it was not abolished completely at later time points [17, 18]. Therefore, if earlier time points were used in isolation, the findings would misleadingly suggest that CSE was solely immunosuppressive. There was unacceptable cytotoxicity at concentrations of CSE greater than 5% (supplementary figure S2), so we opted to use this preparation of CSE to study soluble mediator release from the ALI cultures.
The majority of the subjects with COPD used in this study were current smokers. Our data suggest that the increase in apoptosis, and the increase in caspase-3 activation relative to smokers without airflow obstruction and control subjects in response to CSE exposure was not related to cigarette smoking per se. It appears that the development of COPD itself further contributes to the increased susceptibility of the bronchial epithelium to the cytotoxic effects of CSE. Alternatively the epithelial cells from these individuals may be intrinsically more vulnerable to apoptosis.
Caspase activation has not consistently been shown to be important in CSE induced apoptosis [29], and in fact cell death can occur by mechanisms which are independent of caspase activity [30]. It is feasible that CSE alters the pro- and anti-apoptotic factors which are important players in this increasingly complicated cell death pathway. CSE alters levels of phosphorylated NF-κB in PBEC ALI cultures, and our data suggest that a heightened NF-κB activation may be protective of apoptosis in this context.
Although it could be argued that the process of cell culturing may alter the epithelial phenotype through the multiple cycles of proliferation involved in achieving ALI cultures, recent research suggests this is unlikely to be the case, at least for primary nasal epithelial cells [31]. In addition, there are varied methods used to obtain and establish airway epithelial cell cultures, each with their own merits and limitations. Furthermore, the airways of individuals with COPD may not be sterile. We have discussed these important aspects in the online supplementary material.
To conclude, although airway epithelial cells from patients with COPD exhibit a constitutional pro-inflammatory phenotype, these cells have a diminished inflammatory response to CSE compared to control subjects. Furthermore, these cells have an increased susceptibility to undergo apoptosis. It is not clear if this abnormality results from a change to the cells rendered by the disease process, or whether these particular cells are fundamentally more vulnerable to cell death. Regardless of which explanation holds true, this particular abnormality at least suggests that therapies aimed at reducing the amount of apoptosis may be beneficial. The development inhibitors of apoptosis are realistic ambitions. There are already broad spectrum inhibitors of apoptosis in clinical trials for the treatment of liver disease and they represent the first in their class of agents [32]. Furthermore, this data emphasises the importance of smoking cessation in COPD subjects in order to minimise loss of epithelial cells and to maintain integrity of the bronchial epithelium. Our findings support the notion that cigarette smoke reduces innate lung defence and has the capacity to increase susceptibility to respiratory infections.
Footnotes
This article has supplementary material available from www.erj.ersjournals.com
Statement of Interest
None declared.
- Received April 17, 2012.
- Accepted July 22, 2012.
- ©ERS 2013
REFERENCES










