





Abstract
Platelet-activating factor (PAF)-induced neutrophil lung sequestration may require cell surface adhesion molecules (macrophage‐1 antigen (MAC‐1) and lymphocyte function-associated antigen‐1 (LFA‐1)). In this randomised, double-blinded, crossover study, theneutrophil kinetics after PAF and Lyso-PAF (L‐PAF) airway challenge were investigated in nine mild-intermittent asthmatics.
Neutrophils were measured in peripheral blood (PB) before and at 5, 15, 45 and 240 min after bronchoprovocation, and in induced sputum before and at 240 min after challenge. MAC‐1 and LFA‐1 expression were assessed by immunocytochemistry, and leukotriene B4 (LTB4) was measured by enzyme-immunoassay in induced-sputum supernatants.
Compared with baseline, neutrophils in PB decreased 5 min after PAF, while at 240 min neutrophils in induced sputum increased. Compared with baseline and L‐PAF, PAF decreased the percentages of MAC‐1‐ and LFA‐1‐positive neutrophils in PB at 5 min, but increased the percentages of MAC‐1 and LFA‐1 in neutrophil-induced sputum. Moreover, compared with baseline and L‐PAF, PAF-induced sputum revealed higher LTB4 levels, a finding that correlated with the elevated number of neutrophils in induced sputum.
These findings suggest that macrophage‐1 antigen and lymphocyte function-associated antigen‐1 are involved in platelet-activating factor-induced neutrophil lung traffic, and that this process is modulated by enhanced leukotriene B4 release within the airways.
This study was supported by the Comissionat per a Universitats i Recerca de la Generalitat de Catalunya (2001-SGR00386), a Research Grant (2000) from the European Respiratory Society (ERS), Ministerio de Sanidady Consumo (Redes Temátions de Investigación Cooperativa, CO3/11) and a grant-in-aid from Esteve Group. A. Acuña was supported byFundación Gran Mariscal de Ayacucho, Caracas, Venezuela, and Instituto de Cooperación Iberoamericana (ICI), Spain.
There is growing evidence that neutrophils may play an important role in different forms of asthma 1–6. Indeed, increased numbers and different patterns of activation of neutrophils, and increased levels of neutrophil elastase and interleukin (IL)‐8 have been shown previously 7, 8. Abnormal expression of neutrophil and endothelial adhesion molecules has been shown in blood and sputum cells in mild atopic asthma 9 and during chronic obstructive pulmonary disease (COPD) exacerbations 10. This may result from the increased sequestration of neutrophils in the pulmonary circulation during COPD exacerbations, leading to compartmentalisation of activated neutrophils within the lung 11. However, the precise mechanism by which neutrophils traffic from pulmonary blood to the lung airspaces in both asthma and COPD remains, at least in part, undetermined. The release of leukotriene B4 (LTB4) may lead to neutrophil chemotaxis and neutrophil-endothelial interactions 12, and neutrophils are also an important source of LTB4 13. LTB4 levels in induced sputum are elevated in COPD, correlate with elastase activity 14 and LTB4 is one of the possible mediators involved in neutrophil sequestration in asthma and COPD.
Platelet-activating factor (PAF) is an inflammatory mediator that may be involved in the pathogenesis of bronchial asthma. It has been postulated that PAF-induced transient sequestration of neutrophils within the lung, which has been shown innormals 15–17 and asthmatics 18, could be a useful laboratory-induced model to investigate neutrophil kinetics invivo and some of its mechanisms have been previously investigated by the current authors' group 15, 17, 18. In thecirculating blood compartment, PAF causes a transient neutropaenia followed by a neutrophilia, with recruitment ofneutrophils in lung tissues and into the bronchoalveolar space 16, 17. There is also some evidence for activation of neutrophils, as the neutrophil volume was increased following PAF exposure 16. Whether PAF induces the expression of integrins on the neutrophil, so as to increase its adhesion to the vascular endothelium or respiratory epithelium, to allow egress of the neutrophil from the vascular compartment into the tissues or from the latter into the bronchoalveolar space, respectively, remains elusive. PAF can also promote macrophage‐1 antigen (MAC‐1) (CD11b/CD18)-dependent neutrophil adhesion to endothelial cells through P‐selectin 19, 20 and PAF by itself can increase leukocyte adhesion without altering vessel diameter 21. Likewise, PAF can facilitate CD11b upregulation and L‐selectin shedding induced by neutrophils 22, and can also cause neutrophil adhesion to the endothelium by upregulating intercellular adhesion molecule (ICAM)‐1 23. Furthermore, PAF has the potential to promote neutrophil recruitment by stimulating the release of LTB4 by airway cells in asthmatic patients 24, 25, a mechanism also observed following MAC‐1 overexpression by PAF 26.
Induced sputum is a useful technique to assess airway inflammation 27. Using this tool, the present authors tested whether PAF induces neutrophil accumulation in the airways of patients with asthma through the modulation of neutrophil integrins. The effects of PAF or Lyso-PAF (L‐PAF), its inactive metabolite 28, on the expression of β2‐integrins (MAC‐1 and lymphocyte function-associated antigen‐1 (LFA‐1)) in neutrophils isolated from peripheral venous blood and induced sputum, and the levels of LTB4 in sputum supernatants, were therefore investigated in patients with mild asthma.
Methods
Patients
Nine nonsmoking patients with mild asthma were recruited for the study, which was approved by the ethical research committee, and gave informed written consent. The inclusion criteria were: age >18 yrs and <45 yrs; no respiratory infection or exacerbation of asthma within the preceding 6 weeks; forced expiratory volume in one second (FEV1) ≥70% predicted and ≥1.5 L after discontinuing bronchodilators for 12 h, and positive provocative dose of methacholine causing a 20% fall in the FEV1 (PD20 <1.9 µmol) on their first visit (see below); positive PAF response, as assessed by ≥35% increase in total respiratory system resistance (Rrs) 5 min after PAF (18 µg) inhalation; no previous treatment with oralsteroids; and absence of any systemic or cardiopulmonary disease other than asthma. All subjects were atopic. Maintenance therapy included aerosol short-acting selective β‐adrenergic agonists as needed (table 1⇓).
Patient characteristics and baseline lung function before each challenge
Study design
Patients attended the laboratory on four separate occasions at 08:00 h. All visits were 1 week apart. On the first visit, the clinical and functional assessment was performed. On the second visit, the patient attended for baseline sputum induction. On the next two visits, the patient attended for PAF or L‐PAF bronchoprovocation, in a randomised, double-blinded, crossover manner. The PAF (C16, 1‐O‐hexadecyl‐2‐acetyl-sn-glycero‐3‐phosphocholine; 18 µg; Novabiochem AG, Laufelfingen, Switzerland) and L‐PAF (C16, 1‐O‐hexadecyl-sn-glycero‐3‐phosphocholine, fully saturated; 18 μg; Novabiochem AG) challenge was carried out as previously described 15. A set of duplicate measurements was performed before the bronchoprovocation and at 5, 15, 45 and 240 min following PAF or L‐PAF inhalation. These measurements consisted of Rrs (not at 240 min), ventilatory and cardiac frequency recordings, and peripheral venous blood sampling. Sputum was also induced 240 min after completing the bronchoprovocation, based on a previous study 16. All the challenges were clinically well tolerated by the participants who completed the study. As-needed short-acting β2‐agonists were only allowed until 24 h before the study.
Measurements
Rrs was measured by the forced oscillation technique 15 in each patient. A three-lead electrocardiogram, and cardiac frequency and arterial oxygen saturation via a pulseoximeter (HP M1166A; Hewlett-Packard, Boblingen, Germany), were continuously recorded throughout the study day. Peripheral blood (PB) samples were collected anaerobically through a catheter inserted into a peripheral vein.
Separation of peripheral blood neutrophils
Neutrophils and mononuclear cells were isolated using dextran sedimentation and centrifugation over Ficoll cushions, as previously described 29. Subsequently, neutrophils and mononuclear cell counts and viability were assessed by a haemocytometer and trypan-blue dye exclusion, respectively, and were obtained from 2×105 cells fixed in acetone and stored at −70°C for subsequent immunostaining 29.
Induced sputum and processing
Sputum was induced by the nebulisation of 3, 4 and 5% sterile hypertonic saline solution in sequence for 5 min from a De Vilbiss Ultraneb 99 ultrasonic nebuliser (Healthcare Inc., Somerset, PA, USA) through a mouthpiece without using valves or nose clips, as previously described 27, 29–31. Fresh samples of induced sputum were processed immediately afterwards. Sputum plugs were selected, weighed, mixed with a four-times greater volume of 0.1% dithiothreitol (DTT) and processed, as previously described 4. According to the weight of sputum plugs, sputum volume supernatants differed amongst samples. A minimal sample weight of 200 mg was considered satisfactory for subsequent analysis. Supernatants were stored at −70°C, while cell pellets was resuspended in phosphate-buffered saline (PBS) to assess cell viability, total and differential cell counts, and for cytospins preparation. Differential cell counts were performed on cytocentrifuge preparations stained with May-Giemsa-Gruenwald. In all cases, 200 nonsquamous cells were counted by two blinded observers and results were expressed as percentage of total nonsquamous cells.
Expression of adhesion molecules by peripheral blood and induced sputum neutrophils
Expression of CD11b/CD18 (MAC‐1) and CD11a/CD18 (LFA‐1) (DAKO A/S, Glostrup, Denmark) in both PB and induced sputum was evaluated in cytospins fixed in acetone using two specific monoclonal antibodies (mAbs): mAb 2LPM19c, reacting with the human 165-kd a‐chain (CD11b) of the MAC‐1 protein; and mAb E25.3, directed against the MHM24, reacting with the human 180-kd a‐chain (CD11a) ofthe LFA‐1 protein (DAKO A/S). These antibodies were used at dilutions of 1:100 for 1 h, respectively. An irrelevant antibody anti-immunoglobulin G1 (DAKO) at a 1:10 dilution was used as a negative control. Immunoreactivity was revealed by the streptavidin-biotin alkaline-phosphatase technique. A total of 400 cells per slide were read independently by two observers. Results were expressed as percentage of the positive neutrophils minus total PB or induced sputum cells ratio. The interobserver coefficient of agreement was excellent (Kappa=0.93).
Leukotriene B4 measurement in induced sputum
Sputum LTB4 levels were assessed by a specific enzyme-immunoassay method (Amersham International Plc, Little Chalfont, UK), as previously described 29. The limit of detection for this method is 6 pg·mL−1. The samples were diluted 1:5 before the assay and the value was corrected for the dilutions.
Neutrophil chemotaxis assay
Neutrophils obtained from PB of healthy donors were resuspended at a concentration of 1×106 mL−1 in PBS containing CaCl2 (0.5 mM) and MgCl2 (1 mM), and the supernatants from induced sputum of patients challenged with PAF and L‐PAF were tested for their chemotactic activity onneutrophils. In addition, in order to evaluate whether chemotactic activity of induced sputum samples related to thepresence of LTB4, experiments were performed in the presence and absence of an LTB4 receptor antagonist (LY-223982, 10 µM; Eli Lilly, Basinstoke, UK). Chemotaxis was performed using a 48-well microchemotaxis chamber (Costar; Neuro Probe Inc., Cabin John, MD, USA), as previously described 29. The volume of induced-sputum supernatant used in chemotactic assays was 27 mL. Samples were not diluted before assaying for neutrophil migration. Migration was assessed by counting the number of cells that had migrated into the filter paper of the chemotaxis chamber with a pore size of 3 µm. Each experimental condition was performed in duplicate and three to four fields were assessed for cell migration. Chemotactic activity of the LTB4 standard was also evaluated. Furthermore, the number of cells migrating spontaneously in the presence of a solution 0.1% DTT, at the same concentration of induced-sputum supernatants (i.e. negative control), was subtracted from all measurements before data analysis.
Statistical analysis
Data are expressed as medians (25–75 percentiles). Multiple comparisons for Rrs and neutrophils and expression of adhesion molecules were determined using one-way analysis of variance (ANOVA) with Bonferroni's correction. Sputum cell differences were assessed using Wilcoxon's signed test with Bonferroni's correction. Spearman's rank test was used for correlations. A p<0.05 was considered significant.
Results
Baseline findings
There were no differences in baseline variables between each challenge day (tables 1 and 2⇑⇓), thereby indicating that the 1‐week washout period after each challenge was sufficiently long enough to avoid any carry-over effects. Both ventilatory and cardiac variables (not shown) were within normal limits and remained unchanged throughout the whole study period.
Functional, and cellular and molecular peripheral blood findings before and after each challenge
Systemic, lung resistance and cellular changes after challenge
Eight patients noticed both facial flushing and breathlessness, two had cough, and one experienced mild rhinitis 5 min after PAF. By contrast, no symptoms were evident after L‐PAF. At 5 min after PAF, compared with baseline, Rrs increased (p<0.0001) (table 2⇑) while neutrophils in PB decreased (p<0.004) (fig. 1⇓). Subsequently, compared to 5 min, neutrophils in PB increased at 15 (p<0.0005), 45 (p<0.0002) and 240 min (p<0.0001). By contrast, no changes were seen after L‐PAF.

Peripheral blood a) neutrophils and their expression of b) macrophage‐1 antigen (MAC‐1) and c) lymphocyte function-associated antigen‐1 (LFA‐1) at their respective baselines (BL) and 5 min after Lyso-platelet activating factor (L‐PAF) and PAF. Horizontal bars denote median values. #: p<0.006; ¶: p<0.004; +: p<0.003; §: p<0.002; ƒ: p<0.02.
Compared with baseline and L‐PAF, both induced-sputum total cell count viability and the percentage of squamous cellsafter PAF were not significantly different (table 3⇓). However, neutrophil percentages (neutrophils in induced sputum) increased (p<0.04 and <0.008) (fig. 2⇓) whereas those of macrophages decreased (p<0.02 and <0.008), respectively. By contrast, the percentages of eosinophils, lymphocytes and epithelial cells remained essentially unchanged after each challenge.

Representative immunostainings (in red) for macrophage‐1 antigen (a, c, e and g) and lymphocyte function-associated antigen‐1 (b, d, f and h) by neutrophils isolated from peripheral venous blood (a, b, e and f) and induced sputum (c, d, g and h) at baseline (top row) and at 5 (in peripheral blood) and 240 min (in sputum) after platelet-activating factor (PAF) challenge (bottom row). The data show decreased (peripheral blood) and increased (sputum) expression of both molecules after exposure to PAF compared with their corresponding basal staining.
Sputum characteristics
Immunodetection of β2‐integrins in neutrophils from peripheral blood and induced sputum
Compared with baseline and L‐PAF, 5 min after PAF, the percentages of neutrophils in PB expressing MAC‐1 (p<0.003 each) and LFA‐1 (p<0.004 and <0.02) decreased, respectively (table 2⇑, and figs 1 and 2). A significant correlation was found between the reduction of the percentage of neutrophils in PB expressing MAC‐1 and LFA‐1 expression in PB and the decrease of neutrophils in PB (rho=0.89 and 0.87, respectively; p<0.02, each) 5 min after PAF. No changes were observed after L‐PAF.
By contrast, compared with baseline (10.1 (5.5–12.1) %) and L‐PAF (10.2 (7.1–17.8) %), the percentage of neutrophil-induced sputum expressing MAC‐1 increased (to 25.0 (16.5–35.4) %) (p<0.02 and p<0.008, respectively) after PAF exposure (figs. 2 and 3⇑⇓, and table 3⇑). Similarly, compared with baseline (10.1 (4.4–12.5) %) and L‐PAF (10.3 (7.2–17.7) %), the percentage of neutrophils in induced sputum expressing LFA‐1 increased (to 24.5 (20.1–38.6) %) (p<0.008 each) following PAF, although there were three patients who did not respond, indicating heterogeneity during PAF inhalation. A significant correlation was shown between the increased percentages ofneutrophils in induced sputum expressing MAC‐1 and LFA‐1 and the elevated neutrophils in induced sputum (rho=0.84, p<0.02; and rho=0.73, p<0.04, respectively) after PAF challenge.

Percentages of induced sputum a) neutrophils and their expression of b) macrophage‐1 antigen (MAC‐1) and c) lymphocyte function-associated antigen‐1 (LFA‐1) at their respective baselines (BL) and 240 min after Lyso-platelet-activating factor (L‐PAF) and PAF. Both MAC‐1 and LFA‐1 differences were still significant when their two highest individual values after PAF were excluded from the analysis (n=7; p<0.032 and <0.02, respectively). Horizontal bars denote median values. #: p<0.04; ¶: p<0.008; +: p<0.02.
Leukotriene B4 measurement in induced-sputum supernatant
Compared with baseline (0.32 (0.30–0.60) ng·mL−1) (p<0.02) and L‐PAF (0.44 (0.35–1.05) ng·mL−1) (p<0.03), LTB4 levels increased (to 1.36 (1.09–2.73) ng·mL−1) 240 min after PAF (fig. 4a⇓), an increase that showed a close correlation with the elevated number of neutrophils in induced sputum (rho=0.83, p<0.02) (fig. 4b⇓).
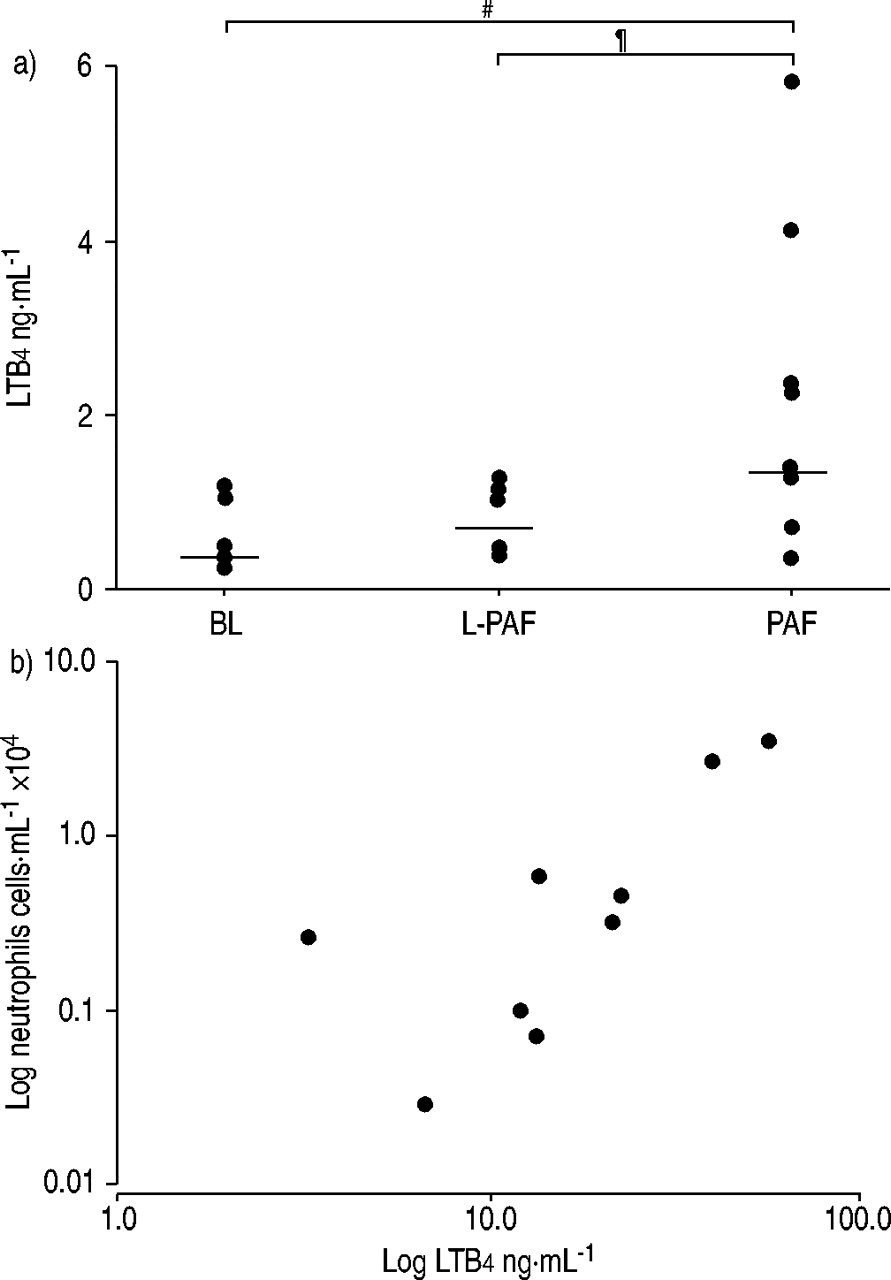
a) Individual leukotriene B4 (LTB4) levels in induced sputum supernatants at baseline (BL) and 240 min after Lyso-platelet-activating factor (L‐PAF) and PAF. These differences were significant when the two highest LTB4 values after PAF were excluded (n=7; p<0.05). Horizontal bars denote median values. #: p<0.02; ¶: p<0.03. b) Plot between increased LTB4 levels in induced sputum supernatant and absolute neutrophils in peripheral venous blood 5 min after PAF challenge, both expressed on a logarithmic scale. rho: 0.83; p<0.02.
Neutrophil chemotaxis
Compared with baseline (31.0 (25.7–40.7) cells·high-power field−1) and L‐PAF (46.0 (27.3–56.5) cells·high-power field−1), there was an increase in neutrophil chemotactic activity (to 169.2 (148.9–199.3) cells·high power·field−1) (p<0.008, each) in induced sputum after PAF (fig. 5⇓). Moreover, pre-incubation of neutrophils with the LTB4 receptor antagonist LY-223982 inhibited the chemotactic activity exerted by induced sputum after PAF by 52.8 (38.7–62.9) %, as shown by a decreased number of migrating neutrophils (to 89.0 (68.0–99.5) cells·high power field−1) (p<0.008) (fig. 5b⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
a) Chemotactic activity of induced sputum supernatant at baseline (BL) and 240 min after Lyso-platelet-activating factor (L‐PAF) and PAF. #: p<0.008. b) Effect of leukotriene B4 (LTB4) receptor antagonist (LY-223982) (LY) on induced sputum neutrophil chemotaxis activity assessed at BL (39.0 (38.7–40.7) cells·high-power field−1) and 240 min after L‐PAF (40.3 (38.2–43.5) cells·high-power field−1) and PAF. The latter was substantially inhibited compared with that induced by PAF. Data is expressed as individual migrated neutrophils per high-power field after substraction of cells migrated inthe negative control (horizontal bars denote median values). ¶: p<0.008.
Discussion
The most novel finding of the present study was an enhanced recruitment of neutrophils expressing MAC‐1 and LFA‐1 4 h after PAF exposure within the airways of patients with mild asthma, and that such enhanced recruitment appeared to be driven by an increased production of LTB4, a potent neutrophil chemoattractant. It was also demonstrated that all these events were associated with a fall in blood neutrophils immediately after PAF and with an increased percentage of induced sputum neutrophils 4 h later.
These findings provide further evidence about some of the mechanisms underlying PAF-induced transient neutrophil pulmonary sequestration in mild asthma, an event observed in both healthy individuals 15–17 and asthmatics 18. To the best of the current authors' knowledge, this is the first study in mild asthma showing that neutrophil recruitment into the airways following PAF exposure is the end result of the chemotactive effects elicited by increased levels of LTB4 on neutrophils expressing selective adhesion molecules, such as MAC‐1 and LFA‐1. The latter are critical for the regulation of neutrophil sequestration within the pulmonary capillaries and their subsequent transmigration across the endothelial pulmonary vascular barrier. Sequestration occurs in response to several intravascular inflammatory mediators and requires at least two sequential steps. First, rapid sequestration of neutrophils into the pulmonary circulation, which results in neutropaenia within 30 s 17, occurs throughout a process that does not require CD11/CD18 and is thought to involve astimulus-induced decrease in the neutrophil's ability to deform 11. Secondly, prolonged sequestration of these intracapillary neutrophils occurs for more than a few minutes, requiring CD11/CD18-mediated interactions with the endothelium 11. Neutrophil migration throughout the pulmonary vessels is tightly regulated by adhesion molecules not only on neutrophils but also on endothelial cells such that, at sites of inflammation 32, ICAM‐1 is highly expressed on activated endothelial cells. Thus, neutrophil adhesion may proceed via direct β2‐integrin ICAM-1 recognition 33. In a recent in-vitro study 34, exposure to PAF was shown to increase neutrophil surface expression of MAC‐1.
The results of the current study indicate that the increased expression of recruitment of neutrophils through both MAC‐1 and LFA‐1 by PAF-induced sputum in asthmatics may play a critical role in the increased ability of white cells to adhere to and migrate across the bronchial and pulmonary vascular endothelium. This may allow for the rolling of neutrophils to stop at the intersection of endothelial cells while establishing rapid adhesion throughout LFA‐1 33, 35. Subsequently, MAC‐1 may promote sustained adhesion of neutrophils to the endothelium, which is a prerequisite for their sequestration into the bronchial and pulmonary capillaries, and their subsequent transmigration into the pulmonary tissue. The fact that PAF challenge may play a key role in neutrophil recruitment is also suggested by the kinetics of the percentages of neutrophils expressing MAC‐1 and LFA‐1 expression in PB and sputum neutrophils. Indeed, compared with baseline and L‐PAF, while the percentages of sputum neutrophils over total cells expressing MAC‐1 and LFA‐1 expressions were substantially increased after PAF, the percentage of peripheral venous blood neutrophils over total cells was decreased at a much earlier phase (i.e. 5 min) after exposure. This data suggests, therefore, that MAC‐1 and LFA‐1 play a role in neutrophil recruitment after PAF, and represent a prerequisite for neutrophil migration across the endothelial barrier. This is further supported by in-vitro evidence showing that PAF increased MAC‐1 expression by neutrophils, which results in both integrin-dependent cell adhesion and extravasation 26, 34.
Nonetheless, expression of β2‐integrins per se, although important, does not seem to be the single mechanism required for an enhanced neutrophil accumulation in the lung. Indeed, leukocyte migration is a rather complex process in which the expression of adhesion molecules is only one of the steps involved. Indeed, for leukocyte transmigration to occur within the lung parenchyma a chemotactic gradient has to be established in order to allow for the recruitment of leukocytes from the bronchial and pulmonary bloodstream into the airway and pulmonary interstitium, respectively. Among the various factors that are capable of promoting airway neutrophil chemotaxis, a central role may be played by LTB4 12. The results of the present study indicate that either PAF challenge released LTB4 within the airways of asthmatic patients or that LTB4 was released by neutrophils recruited and activated by PAF, or both. In addition, it was found that increased LTB4 was significantly correlated with the elevated number of sputum neutrophils recruited by the airways after PAF, thereby suggesting that LTB4 may be involved in neutrophil recruitment within the airways of patients with obstructive airway disorders 36. This hypothesis is further supported by the in-vitro experiments that revealed the ability of sputum samples obtained after PAF to promote neutrophil chemotaxis in a significantly greater manner than that shown at baseline or after L‐PAF. It was also demonstrated that the chemotactic activity of induced sputum from patients with asthma challenged with PAF is significantly greater than that exerted by sputum supernatant obtained at baseline or after L‐PAF. Moreover, the data indicate an important role for sputum LTB4 in the process of neutrophil recruitment, since a LTB4 receptor antagonist inhibited the neutrophil-chemotactic activity exerted by sputum obtained after PAF challenge. Yet, the sputum chemotactic activity of neutrophils did not correlate with its LTB4 concentrations, probably suggesting that other mediators in sputum samples, such as IL‐8 and tumour necrosis factor‐α, may be involved in the process.
Asthmatic rather than normal individuals were chosen for this study for two reasons. First, it has previously been found that asthmatic patients respond to a larger degree of ventilation to perfusion imbalance than healthy individuals, leading to a greater degree of hypoxaemia in asthmatics 15, 18. It was assumed from this that both the inflammatory and integrin responses in asthmatics would be greater. Secondly, previous studies have indicated that neutrophils may play an important role in asthma, particularly in the more severe forms 1–5. Accordingly, the authors wanted to confirm these observations, and if so, examine the role of some of these integrins in neutrophil accumulation, hence the current study was designed to address, at least in part, the issue of neutrophil traffic in asthma.
All in all the data point to the view that the increased leukotriene B4 release in the airways and the subsequent enhanced recruitment of neutrophils may result from the ability of platelet-activating factor to stimulate 5‐lipoxygenase activity in airway cells of asthmatic patients. This hypothesis is further supported by a previous study showing that, after platelet-activating factor exposure, neutrophils and macrophages isolated from bronchoalveolar fluid in asthmatics released a greater amount of leukotriene B4 than in healthy individuals 25, 37. In summary, both β2‐integrins, macrophage‐1 antigen and lymphocyte function-associated antigen‐1, are involved in platelet-activating factor-induced neutrophil lung sequestration in patients with mild asthma, a process that may be modulated by leukotriene B4 release within the airways. Moreover, the current laboratory-induced model ofneutrophil trafficking within the lung may be of interest tofurther explore the mechanisms of neutrophil kinetics andtheir modulation by different interventions in chronic obstructive airway diseases.
Acknowledgments
The authors would like to thank M.T. Carrión, and C. Agustí for their technical advice.
- Received October 25, 2002.
- Accepted March 4, 2003.
- © ERS Journals Ltd
References









