





Abstract
Children with Down syndrome are at high risk for acute respiratory distress syndrome. In Down syndrome, both regulation of inflammation and apoptosis, important in acute respiratory distress syndrome pathophysiology, are abnormal. This has been linked to an imbalance in free radical scavengers. We investigated the expression of free radical scavengers and the effect of oxidative stress in terms of apoptosis and inflammation in respiratory epithelium from children with Down syndrome compared with control subjects.
We cultured primary nasal epithelial cells from Down syndrome children (n=12) and controls (n=17) and exposed them to oxidative stress by supplementing superoxide.
First we showed that the expression of the free radical scavengers CuZn-superoxide dismutase was 28% higher (p=0.06), catalase was 36% lower (p=0.04) and glutathione peroxidase was 73% lower (p=0.004) in Down syndrome children compared with controls. We found no significant difference in apoptosis, between Down syndrome and control subjects after exposure to oxidative stress. We also found no significant difference in levels of interleukin (IL)-1β, IL-6, IL-8, vascular endothelial growth factor and granulocyte colony-stimulating factor in primary nasal epithelial cell supernatant after exposure to oxidative stress between Down syndrome and control subjects.
We found an imbalance in free radical scavengers in respiratory epithelial cells from children with Down syndrome, but this did not result in increased levels of either apoptosis or inflammation upon exposure to oxidative stress.
Abstract
Free radical scavenger imbalance: respiratory apoptosis or inflammation not increased in Down syndrome children http://ow.ly/mxypE
Introduction
Acute respiratory distress syndrome (ARDS) is an acute life-threatening pulmonary condition and one of the major challenges of modern intensive care medicine [1]. In ARDS, the permeability of alveolar epithelial and endothelial barriers is increased, due to inflammation and apoptosis [2]. This results in severe hypoxaemia and respiratory failure. In 2012, the international consensus definition for ARDS has been updated [3]. The term acute lung injury for mild ARDS is now obsolete, and we therefore only refer to ARDS in this manuscript. Both pulmonary and extrapulmonary triggers, for example, pneumonia and aspiration or sepsis and neurotrauma, can induce ARDS. Patient characteristics, such as the presence of chronic disease or genetic make-up, can modulate the risk for developing ARDS [4].
We recently showed that children with Down syndrome have a substantially increased risk for developing ARDS [5]. In Down syndrome, both regulation of inflammation and apoptosis are abnormal, leading to increased risk of infectious diseases and Alzheimer's disease [6, 7]. Whether the increased risk for ARDS in Down syndrome is associated with abnormal regulation of inflammation and apoptosis of the respiratory epithelium has not been investigated.
In Down syndrome, it has been shown that several cell types, such as neurones and granulocytes, have an increased intrinsic level of apoptosis that has been linked to elevated oxidative stress [8, 9]. This increase in oxidative stress has been contributed to the overexpression of CuZn-superoxide dismutase (SOD1), a cytoplasmic free oxygen radical scavenger that is coded for on chromosome 21. SOD1 reduces superoxide to hydrogen peroxide (H2O2), which can be converted by catalase and glutathione peroxidase (GPX) to water. An imbalance in SOD1 and H2O2-reducing enzymes might lead to accumulation of H2O2 and hydroxyl radicals. This increases oxidative stress and may be injurious by increasing inflammation and apoptosis. Such an imbalance has been shown in Down syndrome neutrophils, erythrocytes and in Down syndrome fetal cerebral cortex [10–12]. The expression of SOD1 and H2O2-reducing enzymes and the response to oxidative stress have not yet been investigated in Down syndrome respiratory epithelium.
We hypothesise that an imbalance in free radical scavengers leads to increased oxidative stress resulting in an increased level of apoptosis and inflammation in respiratory epithelium that may contribute to the increased risk of ARDS in children with Down syndrome. In this study, we used nasal epithelial cells as a model for alveolar epithelial cells. This model has been used before as an alternative for lower respiratory tract epithelial cells that, for obvious reasons, are difficult to obtain in children. There is evidence that the response to inflammatory stimuli is similar in nasal and lower airway epithelium [13].
If we could show an increased susceptibility to oxidative stress-driven injury of respiratory epithelium taken from children with Down syndrome, there might be a role for antioxidant therapy to prevent progression to ARDS in critically ill children with Down syndrome. This could potentially decrease morbidity in these children and might provide clues into the importance of oxidative stress in the development of ARDS in the general paediatric population.
The aim of this study was to investigate in an in vitro setting the susceptibility to oxidative stress-driven injury of respiratory epithelium from children with Down syndrome in terms of apoptosis and inflammation, and link this to a potential imbalance in free radical scavengers in Down syndrome compared with control subjects.
Material and methods
Sampling and culturing of primary nasal respiratory epithelial cells
Primary nasal epithelial cells (PNECs) were obtained from children with Down syndrome who visited the outpatient clinic or daycare centre of the Emma Children's Hospital/Academic Medical Center (AMC) (Amsterdam, the Netherlands) for nonpulmonary disorders. Children with a normal karyogram served as controls. Children with concurrent respiratory infections were excluded.
A cytology brush (Cytobrush Plus; CooperSurgical, Trumbull, CT, USA) was introduced 1–2 cm into the nose and rotated along the nasal cavity. The cells were cultured at 37°C in ambient air, supplemented with 5% carbon dioxide, in BEGM-Bulletkit bronchial epithelial medium (Cambrex Cooperation, East Rutherford, NJ, USA) (fig. 1). Details are described in the online supplementary material. After ∼2 weeks of culturing and 1–2 passages, the PNECs were used for experiments at ∼90% confluency.
Primary nasal epithelial cells in a monolayer culture.
Protein expression
SOD1, catalase and GPX expression were measured by separation on SDS-PAGE followed by Western blotting. Details are described in the online supplementary material. Variation in loading was normalised by correlation to actin or tubulin-content. Bound primary antibodies were visualised by IRDye (LI-COR Biosciences, Lincoln, NE, USA) conjugated secondary antibodies. Quantification was performed by infrared fluorescence detection using the Odyssey Imager and software (LI-COR Biosciences).
Exposure to oxidative stress
Two methods to induce oxidative stress in the PNEC cultures were used. For the analysis of apoptosis upon oxidative stress, PNECs were exposed for 3 h to 0–0.2–0.5 mM xanthine and xanthine oxidase 0–16–40 mU·mL−1 (Sigma-Aldrich, St. Louis, MO, USA) generating exogenous superoxide [14]. The PNECs were subsequently detached by exposure to trypsin/EDTA for 5–10 min (Lonza, Basel, Switzerland). The cell suspension was immediately used for flow cytometry.
Cytokine response was undetectable after 3 h of exposure to xanthine/xanthine oxidase, which by then has stopped producing superoxide. Therefore, we used an alternative method to produce superoxide for the detection of the inflammatory response upon oxidative stress. PNECs were exposed for 24 h to 0–6–25–100 μM carbonyl cyanide m-chlorophenyl hydrazone (CCCP, Sigma-Aldrich) generating endogenous superoxide [15]. The supernatant was removed and stored at -80°C. Intracellular oxidative stress was measured by measuring 2',7'-dichlorodihydrofluorescein diacetate (DCFDA) fluorescence. Details are described in the online supplementary material.
Detection of apoptosis
Apoptosis and cell death after exposure to xanthine and xanthine oxidase were determined by flow cytometry. The cells were incubated with APC-labelled annexin-V (Invitrogen, Carlsbad, CA, USA) and propidium iodide (Sigma-Aldrich). The FACSCanto flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and FlowJo software (Tree Star Inc, Ashland, OR, USA) were used for analysis.
Inflammatory response
Cytokine, chemokine and growth factor levels in the supernatant after 24 h of exposure to CCCP were determined by multiplex fluorescent bead assay for interleukin (IL)-1β, IL-6, IL-8, vascular endothelial growth factor and granulocyte colony-stimulating factor (further referred to as “cytokines”) (Luminex; R&D Systems, Minneapolis, MN, USA). These cytokines were chosen based on a two-step selection process. First, we performed a literature search to identify cytokines that are expressed at different levels in Down syndrome compared with control subjects. Secondly, in a pilot study, we used a 30-plex human cytokine panel to investigate which cytokines were expressed in PNEC supernatant.
As a marker for cell damage, lactate dehydrogenase activity was measured using a colorimetric assay to determine NADH production (Cobas 8000, Roche Diagnostics, Mannheim, Germany).
Statistical analysis
Proportions in the patient groups were compared using two-sided Fisher's exact test. For normally distributed data, t-tests were used to compare means. For not normally distributed data, Mann–Whitney U-tests were used to compare medians. For analysis of repeated measures, data were log-transformed to obtain normality and allow for a general linear repeated measures model. Spearman's correlation coefficient was calculated to evaluate the association between protein-expression and flow cytometer and Luminex data. A two-sided p-value of <0.05 was considered statistically significant.
Online depository
In the online supplementary material, we describe details on cell culturing, intracellular oxidative stress measurements, Western blotting and mRNA measurements.
Ethical consideration
The PNEC sampling procedure was approved by the Academic Medical Center (Amsterdam, the Netherlands) ethical committee and informed consent was obtained from the parents of all participants.
Results
Nasal brushes were performed in a total of 34 children. Five brush samples (all from Down syndrome children) did not contain adherent cells (n=2), or the cells succumbed to infection (n=3). We were able to culture the PNECs up to experimental conditions of the remaining 29 children, 12 with Down syndrome and 17 control subjects. Therefore, the nasal brush technique proved to be suitable to harvest PNECs, with few culture failures. Patient characteristics are shown in table 1. The cell cultures produced enough material to perform all analyses in most, but not all experiments. For each analysis, n is given in the figure caption.
The expression of free radical scavengers in PNECs of children with Down syndrome differs from controls
SOD1, catalase and GPX expression are presented in figure 2. SOD1 expression is presented as the ratio SOD1/actin. The mean±sd expression of SOD1 in Down syndrome subjects was 2.30±0.59 versus 1.80±0.54 in controls (p=0.06). Catalase and GPX expression are presented as their ratio with tubulin. The mean±sd expression of catalase in Down syndrome subjects was 0.018±0.006 versus 0.028±0.016 in controls (p=0.04). The median (interquartile range) expression of GPX in Down syndrome subjects was 0.048 (0.026–0.060) versus 0.179 (0.128–0.418) in controls (p=0.004).
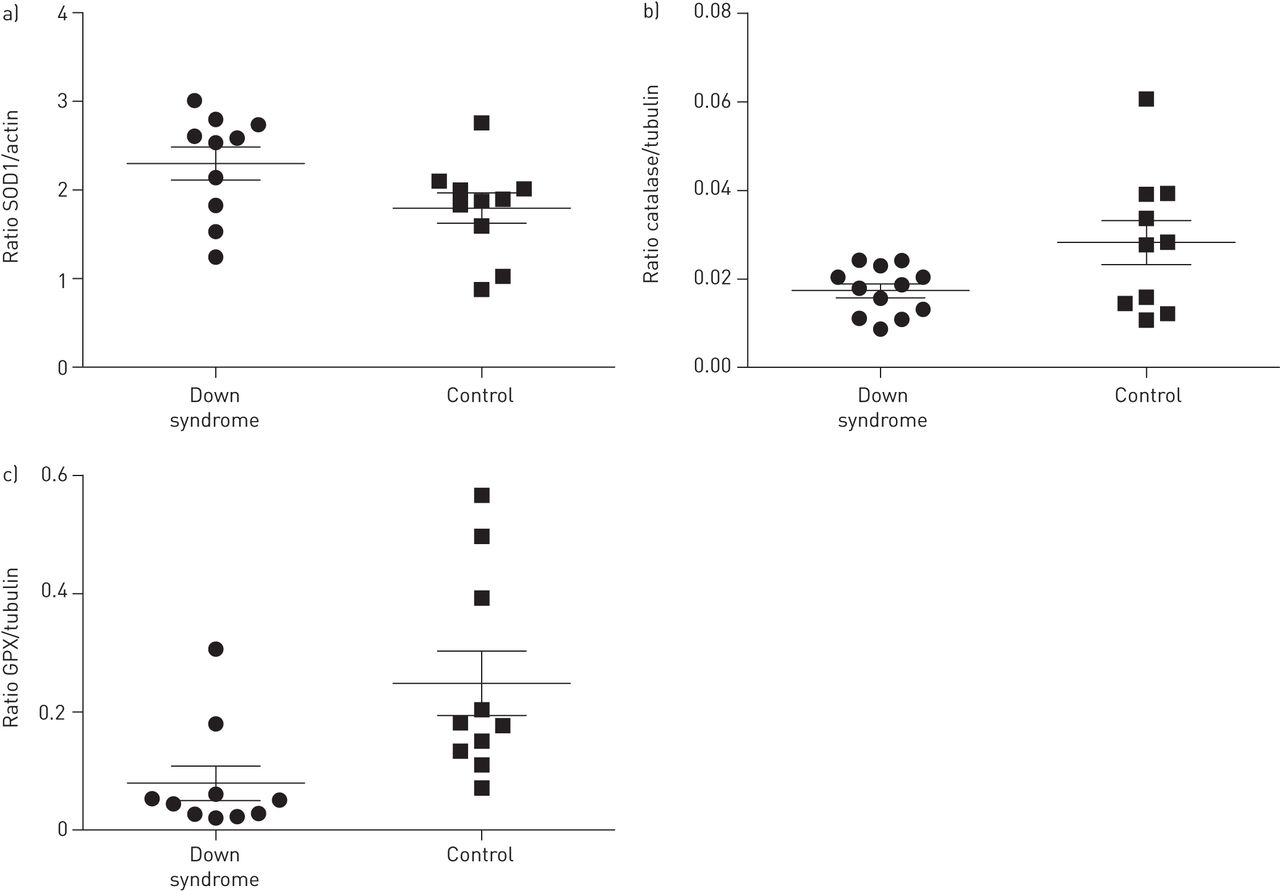
Protein expression of a) CuZn-superoxide dismutase (SOD1), b) catalase and c) glutathione peroxidase (GPX) in primary nasal epithelial cells. Lines represent the mean and bars SEM. Down syndrome: n=10 (for catalase n=12); controls: n=10.
Intracellular oxidative stress is increased by xanthine/xanthine oxidase and CCCP
The xanthine/xanthine oxidase system is known to generate superoxide, and, in far lower concentrations, some H2O2 [14]. By incubating cells with DCFDA prior to extracellular exposure to xanthine/xanthine oxidase, we showed in NCI-H292 cells that this method of extracellular generation of superoxide leads to an intracellular increase of oxidative stress (fig. 3a). CCCP induces the formation of superoxide by uncoupling the mitochondrial protein gradient, thereby preventing ATP synthesis. The production of intracellular oxidative stress by CCCP in our model is shown in figure 3b.

a) Intracellular oxidative stress as measured by 5-(and-6)-chloromethyl-2',7'-dichlorodihydrofluorescein diacetate (DCFDA) fluorescence is increased in H292 cells after exposure to 0–0.5–1.0 mM of xanthine and 0–40–80 mU·mL−1 of xanthine oxidase (XO). b) Intracellular oxidative stress as measured by DCFDA fluorescence is increased in H292 cells after 3 h of exposure to 200 μM of carbonyl cyanide m-chlorophenyl hydrazone (CCCP). There is no increase in DCFDA fluorescence when the cells have been simultaneously incubated with 2 mM of the antioxidant N-acetyl-l-cysteine (NAC).
Oxidative stress-induced apoptosis of PNECs is not significantly different between PNECs of children with Down syndrome and control subjects
Cell death, both necrosis and apoptosis, increased in a dose-dependent pattern after exposure to xanthine and xanthine oxidase in PNECs of both children with Down syndrome and control subjects. We found no difference in expression of annexin-V and/or propidium iodide after exposure to neither dose of xanthine/xanthine oxidase between PNECs from Down syndrome and control subjects (fig. 4). Transformation towards a normal distribution was not possible and, therefore, a GLM repeated measures analysis could not be performed.

Percentage of a) annexin-V positive (AnV), b) propidium iodide positive (PI) and c) both AnV- and PI-positive primary nasal epithelial cells after exposure to 0–0.2–0.5 mM xanthine and 0–16–40 mU·mL−1 xanthine oxidase for 3 h. Lines represent median with interquartile range. Down syndrome: n=9; controls: n=13.
No correlation between expression of SOD1 and cell death upon exposure to superoxide
There were no significant correlations between the expression of SOD1, catalase and GPX and the results of FACS analysis of cell death upon exposure to superoxide in PNECs of either Down syndrome or control subjects. In order to investigate whether an imbalance in H2O2-producing and H2O2-degenerating enzymes correlates with cell death, the ratio between SOD1 and the combined expression of catalase and GPX was computed and tested for correlation with cell death upon superoxide exposure. We did not find such a correlation.
mRNA expression of apoptosis regulating genes in response to oxidative stress in PNECs of children with Down syndrome is not different from controls
The results of the multiplex ligation-dependent probe amplification (MLPA) analysis of apoptosis related mRNA content is presented in figures S1 and S2. In summary, we found induction of Noxa, Nix and NIP3 upon exposure to oxidative stress. Expression of these proteins is known to be responsive to oxidative stress [16, 17]. Expression of survivin was completely inhibited upon exposure to >6 μM CCCP. Survivin is a member of the inhibitors of apoptosis protein (IAP) family and is functionally involved in mitosis. There were no differences between mRNA expression of apoptosis regulatory proteins between PNECs of Down syndrome and control subjects. This is in line with our observation that levels of apoptosis upon exposure to oxidative stress in PNECs are not different between Down syndrome and control subjects. At baseline, the expression of p21, a regulator of cell cycle progression, was lower in PNECs of Down syndrome compared with those of control subjects.
Inflammatory response to oxidative stress in PNECs of children with Down syndrome is not significantly different compared with controls
Results of the multiplex fluorescent bead assay of PNEC supernatant after exposure to oxidative stress are presented in figure 5. We did not find any significant difference in levels of IL-1β, IL-6, IL-8, vascular endothelial growth factor (VEGF) and granulocyte colony-stimulating factor (GCSF) for any condition between PNECs of Down syndrome and control subjects. Levels of IL-6 and GCSF in the supernatant of PNECs of Down syndrome subjects were consistently higher compared with those from control subjects. This finding was not confirmed by general linear repeated measures modelling (p=0.14 for IL-6 and p=0.18 for GCSF). Correlations between cytokine levels and expression of SOD1, catalase and GPX were examined. There were no significant correlations, with the exception of SOD1 and IL-6 levels after exposure to 100 μM CCCP (Spearman's rho 0.55, p=0.04). As shown by an increase of lactate dehydrogenase activity in the supernatant, cell lysis was evident after exposure for 24 h to 100 μM CCCP. Lower concentrations of CCCP (0–6–25 μM) did not result in cell lysis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Levels of a) interleukin (IL)-1β, b) IL-6, c) IL-8, d) granulocyte colony-stimulating factor (GCSF), e) vascular endothelial growth factor (VEGF) andf) lactate dehydrogenase (LDH) in supernatant of primary nasal epithelial cells after exposure to 0–6–25–100 μM carbonyl cyanide m-chlorophenyl hydrazone (CCCP) for 24 h. Box plots represent median, first and third quartiles, and range. Down syndrome: n=10; controls: n=15.
Discussion
We found an imbalance in H2O2-producing and -degenerating enzymes in primary nasal epithelial cells of children with Down syndrome compared with control subjects. However, this did not result in alterations in susceptibility for oxidative stress derived injury in terms of apoptosis or necrosis, or in an altered expression of pro-inflammatory cytokines, chemokines and growth factors.
To the best of our knowledge, this is the first report on expression of SOD1, catalase and GPX in respiratory epithelium of Down syndrome individuals. As SOD1 is coded for on chromosome 21, theoretically, disregarding gene expression regulation, SOD1 could be 50% overexpressed in trisomy 21. We found an overexpression of SOD1 of ∼28% in Down syndrome compared with control subjects.
There are also no previous reports on the effect of oxidative stress exposure on respiratory epithelium in Down syndrome. Most studies regarding oxidative stress and apoptosis in Down syndrome have been performed in various mouse models with a focus on neurons [18]. We recently showed, in human fetal respiratory epithelium, that the percentage of apoptotic cells is not different between Down syndrome and control subjects, but the role of oxidative stress was not evaluated in that study [19].
We hypothesised that overexpression of SOD1 in the absence of elevation of H2O2-reducing enzymes would lead to increased production of H2O2 in primary respiratory epithelial cells, as has been shown to occur in neurones and fibroblasts [8, 20]. However, in a study using a mathematical model, it was shown that overexpression of SOD1 can in fact lead to increased, decreased, or no difference in H2O2 production, depending on the presence of alternative pathways of superoxide consumption and the presence of negative feedback regulation of superoxide production [21]. In experimental studies in lung injury, overexpression of SOD1 seems to have a predominantly beneficial effect [22]. For example, overexpression of SOD1 in mice protects against alveolar apoptosis after ceramide-induced oxidative stress [23]. In our study, despite an unfavourable ratio in the expression of SOD1, catalase and GPX, we found no deleterious effect in terms of increased apoptosis or inflammation upon exposure to oxidative stress. However, the overexpression of SOD1 also did not lead to protection against oxidative stress. Thus, in our in vitro study, SOD1 expression does not seem to correlate with apoptosis in PNECs. This might be explained by an overcapacity or adaptation of other enzymes of the antioxidant system, eliminating the effect of SOD1 overexpression. Also, apoptosis is a highly regulated process and other apoptotic pathways may counterregulate the effects of SOD1 overexpression. Finally, other cell compartments that are important in the pathophysiology of ARDS, such as endothelial cells, might be more affected by oxidative stress caused by SOD1 overexpression.
We did not anticipate a decrease in activity of catalase and GPX in PNECs. Catalase and GPX are coded for on chromosomes 3 and 11, respectively. While SOD1 is consistently overexpressed in Down syndrome, the activity of catalase and GPX depends on the tissue being investigated. Previous studies have shown that their activity in Down syndrome is either comparable or increased compared with controls [24], but this is not a consistent finding [25].
We did not investigate other proteins that have been associated with oxidative stress and disease in Down syndrome, such as ETS-2, APP and DSCR1 [21]. However, we studied an end-point of the effect of oxidative stress on PNECs (apoptosis and necrosis) and found no difference between Down syndrome and control subjects. It therefore does not seem likely that these proteins play an important role in the effect of oxidative stress on cell death in Down syndrome PNECs.
Of the cytokines we have investigated in this study (IL-1β, IL-6, IL-8, VEGF and GCSF), all but GCSF have been shown to be increased in plasma of patients with ARDS [26]. In bronchoalveolar lavage fluid, all but VEGF have been shown to be elevated in ARDS. In vitro, we found an increase in IL-1β, IL-8 and VEGF in PNEC supernatant after exposure to increasing levels of oxidative stress. IL-6 and GCSF decreased in PNEC supernatant of both Down syndrome subjects and controls after exposure to increasing levels of oxidative stress. This might be determined by the time-point at which we sampled. Alternatively, in vivo, other cells beside the respiratory epithelium might be responsible for the increase in IL-6. While expression of VEGF was not different between Down syndrome and control subjects in our study, the angiogenic responses to VEGF might be inhibited in Down syndrome subjects, as was shown recently in a Down syndrome mouse model [27].
Although not significant, we found the level of both IL-6 and GCSF in the PNEC supernatant to be consistently higher in Down syndrome compared with control subjects, irrespective of the level of oxidative stress. IL-8 was not significantly increased in Down syndrome versus control subjects in all but the highest exposure to oxidative stress. This is in line with previous studies, showing an increased level of plasma IL-6 and IL-8 in children with Down syndrome [28, 29]. In granulocytes, GCSF has been shown to inhibit apoptosis in Down syndrome but not in controls [9]. In our study, in both Down syndrome and control subjects, we did not find a correlation between GCSF level and apoptosis.
We used MLPA to investigate for differences in regulation of apoptosis between Down syndrome and control subjects. These results should be interpreted with caution due to the small sample size. It is not very likely, but still not impossible, that regulation of apoptosis is different between the two groups, while the end result, percentage of apoptotic cells, is not different between the groups.
The results of our study do not support the concept that the high risk for ARDS in children with Down syndrome can be explained by increased apoptosis of alveolar epithelial cells and inflammation caused by an imbalance of free radical scavengers. Theoretically, other pro-apoptotic stimuli besides oxidative stress might be responsible for increased alveolar apoptosis in Down syndrome. However, whether there is in fact increased apoptosis in the lung in ARDS in Down syndrome versus controls has yet to be established.
Based on our results, it does not seem rational to perform studies into the capability of antioxidant therapy to prevent ARDS in critically ill children with Down syndrome. However, we cannot rule out that the nasal epithelial cells used in this study respond differently to oxidative stress than does alveolar epithelium. Besides, airway epithelial cells that are exposed to a trigger that could lead to ARDS, such as a severe infection, might react differently than the healthy cells used in this study. Furthermore, we did not study the alveolar endothelium, which also plays a key role in the pathogenesis of ARDS [2]. Endothelium might have a different sensitivity to oxidative stress in Down syndrome and thus contribute to the high risk for ARDS in Down syndrome. Finally, our results do not preclude the possibility that antioxidant therapy might be beneficial in the ARDS population as a whole, and its effect might be dependent of both the cause of ARDS and the stage of disease progression. Animal studies with both intravenous and aerolised SOD, catalase and their mimetics have been shown to protect against lung injury in several models. Treatment with other forms of SOD besides intracellular SOD1, such as extracellular SOD and mitochondrial SOD, have proven to be more protective than the intracellular form of SOD that is overexpressed in Down syndrome. However, few studies have been performed in humans and results have been disappointing [22, 30].
In conclusion, we found an imbalance in free radical scavengers in airway epithelium of children with Down syndrome, but this did not result in increased levels of either apoptosis or inflammation upon exposure to oxidative stress.
Acknowledgments
The authors wish to thank Barbara Smids-Dierdorp, Tamara Dekker, Annemiek Dijkhuis (Dept of Experimental Immunology and Respiratory Medicine) and René Leen (Dept of Genetic Metabolic Diseases, Academic Medical Center, University of Amsterdam, the Netherlands) for laboratory assistance; and Jan Pieter Marchal and Paul van Trotsenburg (Dept of Paediatric Endocrinology, Emma Children's Hospital/AMC, Amsterdam, the Netherlands) who have assisted in the inclusion of children with Down syndrome.
Footnotes
This article has supplementary material available from www.erj.ersjournals.com
Support statement: Support was granted by the following nonprofit organisations: Tysfonds (Middenmeer, the Netherlands), Steun Emma Kinderziekenhuis AMC Foundation (Amsterdam, the Netherlands), and the Janivo Foundation (Zeist, the Netherlands).
Conflict of interest: None declared.
- Received August 6, 2012.
- Accepted November 5, 2012.
- ©ERS 2013
References










