





Abstract
Mutations in the gene encoding bone morphogenetic protein (BMP) receptor type 2 (BMPR-2) have been reported in pulmonary arterial hypertension (PAH), but their functional relevance remains incompletely understood.
BMP receptor expression was evaluated in human lungs and in cultured pulmonary artery smooth muscle cells (PASMCs) isolated from 19 idiopathic PAH patients and nine heritable PAH patients with demonstrated BMPR-2 mutations. BMP4-treated PASMCs were assessed for Smad and p38 mitogen-activated protein kinase (MAPK) signalling associated with mitosis and apoptosis.
Lung tissue and PASMCs from heritable PAH patients presented with decreased BMPR-2 expression and variable increases in BMPR-1A and BMPR-1B expression, while a less important decreased BMPR-2 expression was observed in PASMCs from idiopathic PAH patients. Heritable PAH PASMCs showed no increased phosphorylation of Smad1/5/8 in the presence of BMP4, which actually activated the p38MAPK pathway. Individual responses varied from one mutation to another. PASMCs from PAH patients presented with an in vitro proliferative pattern, which could be inhibited by BMP4 in idiopathic PAH but not in heritable PAH. PASMCs from idiopathic PAH and more so from heritable PAH presented an inhibition of BMP4-induced apoptosis.
Most heterogeneous BMPR-2 mutations are associated with defective Smad signalling compensated for by an activation of p38MAPK signalling, accounting for PASMC proliferation and deficient apoptosis.
- Bone morphogenetic protein receptor type 2
- intracellular signal transduction
- mutation
- pulmonary hypertension
- smooth muscle cells
- vascular remodelling
Pulmonary arterial hypertension (PAH) is an uncommon disease with a poor prognosis and mysterious pathobiology, characterised by a progressive increase in pulmonary vascular resistance and eventual right ventricular failure 1. Mutations of bone morphogenetic protein (BMP) receptor type 2 (BMPR-2), a member of the transforming growth factor (TGF)-β receptor family, have been reported in a high proportion of patients with the heritable form of the disease, and in 10–30% of patients with sporadic idiopathic PAH 2. To date, more than 200 distinct BMPR-2 mutations have been described, widely dispersed across the gene, with the majority predicting premature truncation of the transcript 3. BMPR signalling involves heterodimerisation of two transmembrane serine/threonine-kinase receptor chains, the constitutively active BMPR-2 and the corresponding type 1 receptor BMPR-1A/ALK3 or BMPR-1B/ALK6 4, 5. With interaction of a ligand, for example BMP4, the activated kinase domain of BMPR-2 phosphorylates the corresponding BMPR-1, which in turn initiates intracellular signalling through the phosphorylation of a set of BMP-restricted Smad proteins (Smad1/5/8). Subsequently, these phosphorylated Smads associate with Smad4, translocate to the nucleus and then modulate the transcription of target genes. Alternative Smad-independent signalling pathways involving mitogen-activated protein kinase (MAPK), including extracellular signal-regulated kinase (ERK)1/2, Jun N-terminal kinase (JNK) and p38MAPK, have been reported to be activated by BMP ligands 6. The resulting imbalance is believed to be the cause of a proliferation of pulmonary artery smooth muscle cells (PASMCs), a major component of pulmonary arteriolar remodelling in PAH 1.
It is of interest that the histopathology and clinical picture of PAH with or without BMPR-2 mutations appear similar, except for an earlier age of onset, more severe haemodynamic compromise at diagnosis and less common reversibility at vasodilator testing 7, 8. Therefore, the functional consequences of BMPR-2 mutations remain incompletely understood, but it may be hypothesised that their phenotypic impact may vary with type of mutation or interaction with alternative signalling pathways.
It has previously been reported that PASMCs from idiopathic PAH patients present with an in vitro proliferative phenotype 9, 10. In the present study, we investigated the effects of BMP4 on Smad and p38MAPK signalling associated with mitosis and apoptosis in cultured PASMCs isolated from idiopathic PAH patients without detected mutations and from heritable PAH patients with mutations. The results are in keeping with the notion of a crucial role for BMP/Smad signalling in the prevention of abnormal growth and apoptosis of PASMCs that is lost in most but not all types of mutations.
METHODS
Tissue samples
Lung tissue and pulmonary arteries were sampled at lung transplantation and sequenced to screen for BMPR-2 mutations. After confirmatory cross-check with medical records, patients with PAH were segregated into two groups, according to the presence or absence of mutations. These two groups were respectively called heritable PAH (n = 9) and idiopathic PAH (n = 19) patients. Pulmonary specimens were also sampled in control subjects (n = 10) at lobectomy or pneumonectomy for a suspected localised lung tumour. These control subjects did not bear any BMPR-2 mutations or polymorphisms.
All PAH patients were in New York Heart Association functional class III or IV and were treated with i.v. epoprostenol. In the control subjects, transthoracic echocardiography was performed pre-operatively to rule out pulmonary hypertension, and pulmonary arteries were sampled at a distance from tumour areas. The study was approved by the local Institutional Review Board (Ethics Committee, CPP Ile-de-France VII, Le Kremlin-Bicêtre, France), and patients provided informed consent prior to their contribution to the study.
Screening for mutations in the gene encoding the BMPR-2 receptor
Mutations in the BMPR-2 gene in lung specimens from patients with PAH (n = 28) were screened as previously described 11, 12. Briefly, the entire protein-coding region (sequence corresponding to exons 1–13 of the BMPR-2 gene) was amplified from genomic DNA samples by PCR with specific primers. PCR products were then separated by electrophoresis in a 1% agarose gel and purified using the QIAquick PCR purification kit (QIAGEN, Courtaboeuf, France). Amplified and purified fragments were sequenced wih a dye-terminator cycle-sequencing system (ABI PRISM 377, Perkin-Elmer Applied Biosystems, Courtaboeuf).
Culture of human PASMCs and pulmonary microvascular endothelial cells
Human PASMCs were cultured from explants of pulmonary arteries (1.5–10 mm in diameter) derived from previously described patient groups transplanted for heritable and idiopathic PAH, and also from controls. PASMCs were cultured in 10% fetal calf serum (FCS) in Dulbecco’s modified Eagle medium (DMEM) and used between passages three and six, as previously described 13. The phenotype of cultured PASMCs was assessed for expression of muscle-specific contractile and cytoskeletal proteins, including smooth muscle α-actin (α-SMA), desmin and vinculin 13.
Human pulmonary microvascular endothelial cells (PECs) were obtained by dispase I (Roche Diagnostics, Penzbeg, Germany) digestion followed by immunomagnetic purification with anti-platelet endothelial cell adhesion molecule-1 (CD31) monoclonal antibody-labelled Dynabeads (Dynal Biotech, Compiegne, France) of a fragment of lung tissue isolated from heritable and idiopathic PAH patients and controls, as previously described 9, 14. To characterise the endothelial phenotype, PECs were labelled with acetylated low-density lipoprotein coupled to a fluorescent carbocyanine dye (1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (Dil-Ac-LDL); Tebu, Le Perray en Yvelines, France) and stained with antibodies against the endothelial cell-specific lectin Ulex europaeus agglutinin-1 (UEA-1; Sigma-Aldrich, Irvine, UK) 15. Experiments were also performed with monoclonal antibodies against desmin and vimentin (Dako, Glostrup, Denmark). Cells with positive staining for Dil-Ac-LDL and UEA-1 and negative staining for desmin and vimentin were taken as endothelial cells and constituted >95% of our PEC cultures. PECs were used between passages three and six 9, 14.
RNA extraction and cDNA preparation
Total RNA was prepared from snap-frozen human lung tissue samples (weight 100 mg) by homogenisation according to the method of Chomczynski and Sacchi 16, using TRIzol reagent (Invitrogen, Cergy-Pontoise, France). Total RNA was extracted from growth-arrested primary cultures of human PASMCs and PECs using Qiagen RNeasy Mini kit (QIAGEN), according to the manufacturer’s instructions. RNA concentration was determined by standard spectrophotometric techniques and the RNA integrity was assessed by visual inspection of ethidium bromide-stained denaturing agarose gels. First-strand cDNA synthesis was carried out using SuperScript II Reverse Transcriptase System (Life Technologies, Inc., Carlsbad, CA, USA), as previously described 9, 14.
Real-time quantitative PCR
Real-time quantitative (RTQ)-PCR primers were designed using the computer program Primer3 (Primer Express Software, Applied Biosystems) for human BMPR-1A, BMPR-1B, BMPR-2, Bax and Bcl2 mRNA, and for 18s ribosomal RNA as a housekeeping gene. To avoid inappropriate amplification of residual genomic DNA, intron-spanning primers were selected. RTQ-PCR was performed in triplicate on an ABI PRISM 7000 (Applied Biosystems, Foster City, CA, USA), in mixtures of 12.5 μL Sybr Green PCR Master Mix (Applied Biosystems, Warrington, UK), 300 nM (each) primer and 5 μL of diluted template DNA in a total volume of 25 μL. Signal detection and analysis of results were performed with ABI PRISM 7000 sequence detection software (Applied Biosystems). Relative quantification was achieved with the comparative 2-ΔΔCt method by normalisation with 18s ribosomal RNA. For assays of Bax and Bcl2 mRNAs, PASMCs were seeded and synchronised. The cells were then exposed to BMP4 (100 ng·mL−1) for 4 h and then used for mRNA extraction and RTQ-PCR.
Protein extraction and BMPR-1A, BMPR-1B and BMPR-2 western blotting
Proteins were extracted from snap-frozen tissue samples (weight 100 mg) by homogenisation in an appropriate amount of homogenising buffer (Complete Mini Protease Inhibitor Cocktail (Roche Diagnostics, Mannheim, Germany) in PBS and 0.1% Triton X-100). The homogenates were centrifuged at 4°C and the supernatants were collected. After determination of the protein concentration using the method of Bradford 17, 40 μg of protein from each lung sample were resuspended in 3× Laemmli buffer, boiled for 5 min, and separated on 10% acrylamide gel by electrophoresis. Proteins were electrophoretically transferred to a nitrocellulose membrane (Sigma-Aldrich, Irvine, UK) for 1 h at room temperature. After blocking with 5% bovine serum albumin (BSA) in 1× Tween (T)-TBS (10 mM Tris-HCl pH 8.0, 150 mM NaCl and 0.1% Tween 20) for 2 h at room temperature, the membrane was washed three times with T-TBS at room temperature for 5 min. The membrane was incubated with goat anti-human BMPR-1A, BMPR-1B or BMPR-2 antibody (1:500; R&D systems, Minneapolis, MN, USA) at 4°C overnight with rocking. Then the membrane was washed three times for 5 min and incubated with the secondary antibody (rabbit anti-goat immunoglobulin (Ig)G conjugated with horseradish peroxidase; Dako, Glostrup, Denmark; 1:2,000) for 1 h at room temperature. Immunoreactive bands were detected using the enhanced chemiluminescence western blotting analysis system (Amersham Pharmacia Biosciences, Little Chalfont, UK) and quantified by laser densitometry. Relative quantification was performed by normalisation with β-actin (Sigma-Aldrich, St Louis, MO, USA).
Immunoblotting for BMP signalling pathways
PASMCs were plated in fresh 10% FCS/DMEM medium for 24 h and then quiesced for 48 h in serum-free medium. BMP4 (100 ng·mL−1) or vehicle was then added to the cells for 20 min. Protein was harvested by washing cells in cold PBS and by scraping in 300 μL of 1× sample loading buffer (Tris-HCl pH 7.4, NaCl, NaF, sodium pyrophosphate (all at 25 mM), sodium vanadate (1 mM), EDTA, EGTA (both at 2.5 mM), phenylmethylsulfonyl fluoride (1 mM), aprotinine, leupeptine (both at 5 μg·mL−1), SDS, deoxycholate and NP-40 (all at 0.50%)) on ice. The samples were then stored at -20°C. After determination of the protein concentration, using the method of Bradford 17, samples (20 μg) were resuspended in 3× Laemmli buffer, boiled at 95°C for 5 min and electrophoresed on acrylamide gels (10%). Immunoblotting assays were performed as described above with monoclonal mouse anti-human phospho-p38MAPK(Thr180/Tyr182) (1:1000; Cell Signaling Technology Inc., Danvers, MA, USA), polyclonal rabbit anti-human phospho-Smad1(Ser463/465)/Smad5(Ser463/465)/Smad8(Ser426/428) (1:1000; Cell Signaling Technology Inc.) and polyclonal goat anti-human total Smad 1/5/8 (1:1000; Santa-Cruz Biotechnology, Santa Cruz, CA, USA). Relative quantification was performed by normalisation with total Smad1/5/8 for phospho-Smad1/5/8 and β-actin (Sigma-Aldrich, Lyon, France) for p38MAPK. Treatment with 100 ng·mL−1 for 20 min was chosen based on preliminary studies of BMP4 concentrations in relation to the capacity to activate downstream signalling pathways and to inhibit growth-promoting activity of mitogenic agents on PASMCs 18, 19.
PASMC proliferation assays
The growth of human cultured PASMCs was determined by [3H]-thymidine incorporation, representing DNA synthesis. Briefly, PASMCs were seeded in 24-well plates in 10% FCS/DMEM at a density of 5×104 cells·well−1 and allowed to adhere for 24 h. The medium was then removed and the cells subjected to growth arrest by incubation with serum-free DMEM. After 48 h, the medium was replaced with fresh DMEM containing 10% FCS or 10 ng·mL−1 platelet-derived growth factor (PDGF) in the presence or absence of 100 ng·mL−1 BMP4. PASMC proliferation was also assessed in response to 10% FCS and 10 ng·mL−1 PDGF alone. For each condition, [3H]-thymidine (0.6 μCi·mL−1) was added to each well. After incubation for 24 h, the cells were washed twice with PBS, treated with ice-cold 10% trichloroacetic acid and neutralised with 0.1 N NaOH (0.5 mL·well−1). [3H]-thymidine incorporation into DNA was counted and reported as counts per minute per well.
Apoptosis assays
Apoptosis evaluation was performed by flow cytometry analysis of the DNA content by propidium iodide incorporation and RTQ-PCR analysis of the Bax/Bcl2 ratio. For flow cytometry analysis of DNA content, PASMCs were seeded and treated for 24 h with fresh serum-free DMEM in the presence or absence of BMP4 (100 ng·mL−1). Culture medium was removed and saved. Cells were trypsinised and returned to the medium they had grown in and then centrifuged. Cells were then washed twice in ice-cold PBS and stored at 4°C in 75% ethanol. Fixed cells were centrifuged, washed with PBS and incubated with 200 μL Rnase I (1 mg·mL−1; Invitrogen) and 200 μL of propidium iodide (1 mg·mL−1; Sigma-Aldrich, Lyon, France). Cells were incubated at room temperature for 1 h in the dark. Samples were analysed by flow cytometry. The red fluorescence of single events was recorded using a laser beam at 488 nm excitation wavelength with 610 nm as emission wavelength, to measure the DNA index. For Bax/Bcl2 ratio determination, PASMCs were seeded, synchronised and treated for 4 h as described above. mRNA extraction, cDNA synthesis and RTQ-PCR were performed to determinate the expression of pro-apopototic Bax and anti-apoptotic Bcl2 genes compared with 18s as a housekeeping gene, as described above.
Statistical analyses
All data are reported as mean±sem. Effects of BMPR-2 mutations and BMP4 treatment were analysed by repeated-measures ANOVA. When the F-ratio of the ANOVA reached a critical value of p<0.05, nonparametric Mann–Whitney tests were used to compare specific situations 20. A linear squared regression analysis was used to calculate correlations between pulmonary vascular resistance and content (mRNA and protein) of the investigated BMP signalling molecules 20.
RESULTS
Clinical and haemodynamic characteristics of PAH patients
There were no differences between PAH patients with and without BMPR-2 mutations in terms of age (41±2 versus 39±3 yrs), female to male sex ratio (10/9 versus 5/4), mean pulmonary artery pressure (62±2 versus 63±3 mmHg), pulmonary vascular resistance (20±1 versus 20±2 U·m−2) and cardiac index (2.19±0.09 versus 2.41±0.25 L·min−1·m−2). None of the patients presented with reversibility at vasodilator testing.
Identification and description of BMPR-2 mutations
Germline mutations in the 13 exons encoding BMPR-2 were identified in the nine heritable PAH patients (fig. 1⇓). Three heterozygous nonsense mutations were identified in exons 1 (W16X), 2 (W70X) and 3 (S107X), which encode part of the extracellular domain of BMPR-2. One mutation consisted of total deletion of exon 1 (Δexon1). Two mutations were found in exon 5, which encodes the transmembrane domain of BMPR-2: one heterozygous nonsense mutation (E195X) and one causing loss of 22 bp (22 bp del). Two heterozygous missense mutations and one nonsense mutation were identified respectively in exons 7 (S301P) and 11 (R491W and Q495X), which encode parts of the kinase domain of BMPR-2. No ALK-1 mutation was found in the PAH patients with and without mutations.

Genetic characteristics of patients. Schematic representation of bone morphogenetic protein receptor type 2 (BMPR-2) functional domains, demonstrating the range of BMPR-2 mutations studied in this study and indicating the nature of amino acid substitution or nonsense mutations (X).
Pulmonary and cellular expression of BMPRs
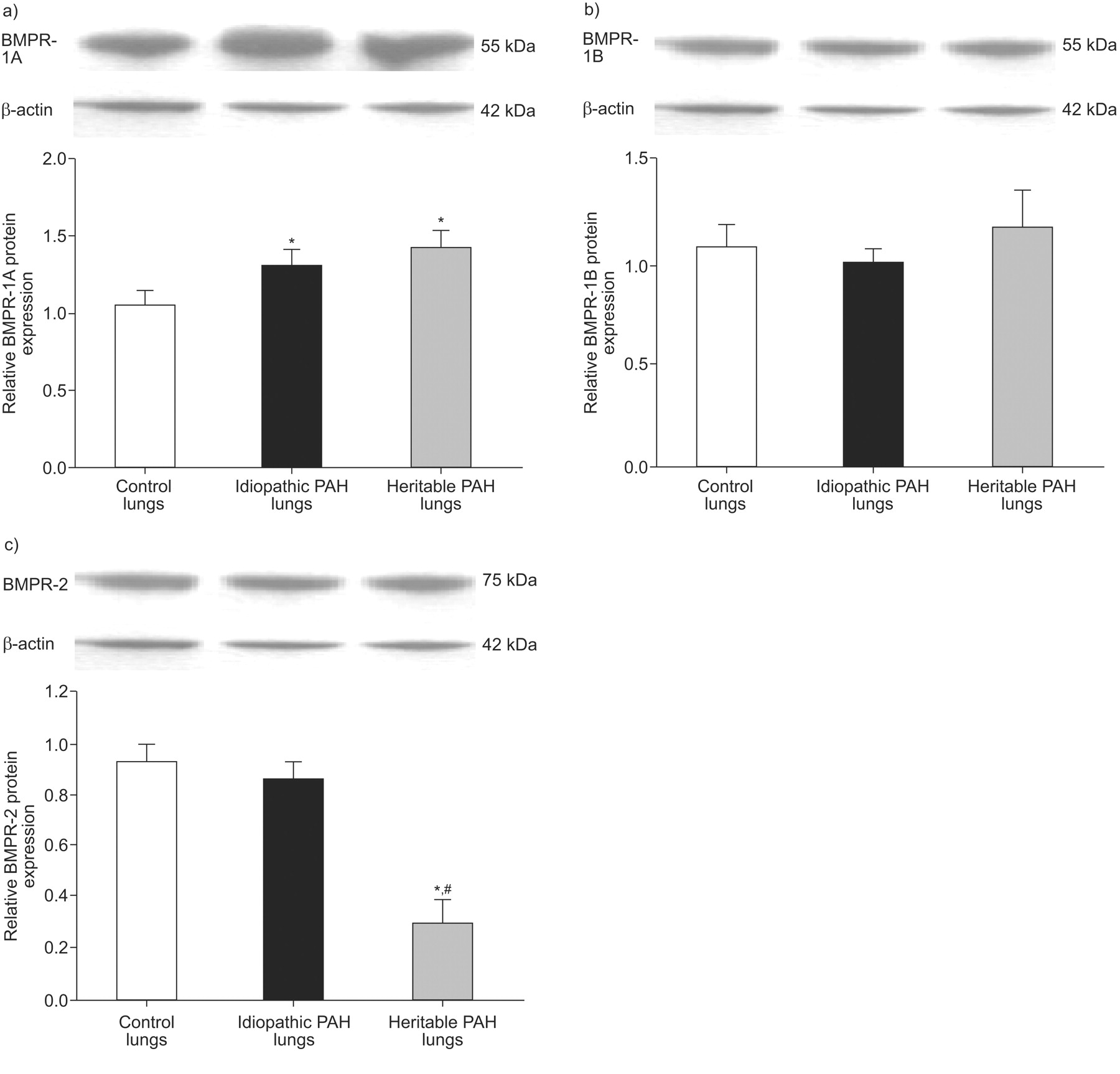
The expression of BMPR-1A mRNA and protein was increased in lung tissue from both heritable and idiopathic patients (figs 2a⇓ and 3a⇓), but mRNA expression was only increased in PASMCs from idiopathic PAH patients (fig. 2b⇓), and was not different from controls in PECs from heritable and idiopathic PAH patients (fig. 2c⇓). The expression of BMPR-1B mRNA was increased in PASMCs from heritable PAH patients only (fig. 2e⇓). The expression of BMPR-2 mRNA and protein was decreased in lung tissue from heritable PAH only (figs 2g⇓ and 3c⇓), and the mRNA expression was decreased in PASMCs from both heritable and idiopathic PAH (fig. 2h⇓), and was not different from controls in PECs (fig. 2i⇓). However, as indicated by relatively large sem values, the increased expression of BMPR-1A and BMPR-1B and decreased expression of BMPR-2 in PASMCs varied greatly from one mutation to another, with no consistent pattern (fig. 4⇓). No correlation was found between pulmonary vascular resistance and lung expression of BMPR-1A, BMPR-1B and BMPR-2 proteins.

Relative a–c) bone morphogenetic protein receptor type 1A (BMPR-1A), d–f) BMPR-1B and g–i) BMPR-2 mRNA expression. Whole lung tissue samples (a, d and g), cultured pulmonary artery smooth muscle cells (PASMCs; b, e and h) and pulmonary microvascular endothelial cells (PECs; c, f and i) from controls (□), pulmonary arterial hypertension (PAH) patients without (▪; idiopathic PAH) and with BMPR-2 mutations (▓; heritable PAH) were assessed by real-time quantitative PCR. Results are expressed as mean±sem. Statistical differences were assessed by the Mann–Whitney test. *: p<0.05 versus control conditions; ***: p<0.001 versus control conditions; #: p<0.05 idiopathic PAH versus heritable PAH conditions; ###: p<0.001 idiopathic PAH versus heritable PAH conditions.

Relative a) bone morphogenetic protein receptor type 1A (BMPR-1A), b) BMPR-1B and c) BMPR-2 protein expression in whole lung tissue samples from controls (□), pulmonary arterial hypertension (PAH) patients without (▪; idiopathic PAH) and with BMPR-2 mutations (▓; heritable PAH) were assessed by western blotting. Results are expressed as mean±sem and as expression relative to β-actin. Statistical differences were assessed by the Mann–Whitney test. *: p<0.05 versus control conditions; #: p<0.05 idiopathic PAH versus heritable PAH conditions.

Relative a) bone morphogenetic protein receptor type 1A (BMPR-1A), b) BMPR-1B and c) BMPR-2 mRNA expression in pulmonary artery smooth muscle cells isolated from heritable pulmonary arterial hypertension patients with naturally occurring BMPR-2 mutations (Δexon1, G/A = W16X, G/A = W70X, C/G = S107X, G/T = E195X, 22 bp del, T/C = S301P, C/T = R491W and C/T = Q495X). Results were assessed by real-time quantitative PCR.
Differential effects of BMP4 on Smad and p38MAPK signalling
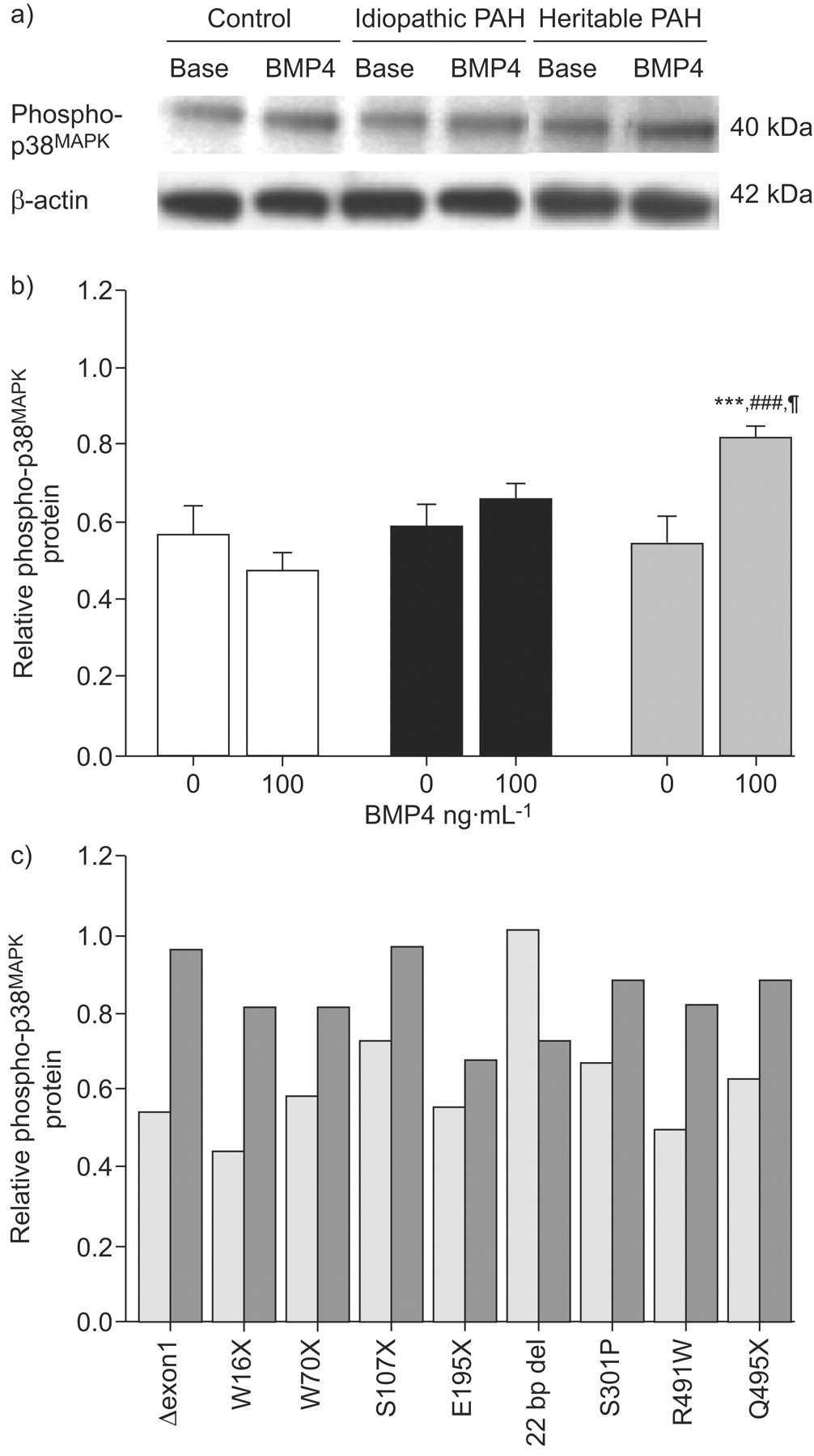
BMP4 (100 ng·mL−1 for 20 min) induced the activation (phosphorylation) of Smad1/5/8 in PASMCs isolated from idiopathic PAH patients and controls (fig. 5⇓), indicating that the transmission of BMP signalling was intact in these cells. In contrast, no BMP4-induced phosphorylation of Smad1/5/8 was observed in PASMCs from heritable PAH, with the exception of the 22 bp del mutation (fig. 5c⇓). BMP4 activated p38MAPK signalling in PASMCs from heritable PAH patients (with the exception of the PASMCs with 22 bp del mutation) but not from idiopathic PAH patients or controls (fig. 6⇓).

Smad 1/5/8 phosphorylation induced by bone morphogenetic protein (BMP)4. 90% confluent pulmonary artery smooth muscle cells (PASMCs) isolated from controls (n = 10), pulmonary arterial hypertension (PAH) patients without (idiopathic PAH; n = 10) and with BMP receptor type 2 (BMPR-2) mutations (heritable PAH; n = 9) were stimulated with 100 ng·mL−1 BMP4 for 20 min, followed by lysis for total protein. a) Representative western blots for phospho-Smad 1/5/8 and for total Smad 1/5/8 to show equal loading. b) Densitometry of phospho-Smad 1/5/8 and total Smad 1/5/8 bands from western blots of control (□), idiopathic PAH (▪) and heritable PAH (▓) PASMCs. Results are presented as relative protein expression ratio of phospho-Smad1/5/8 and total Smad1/5/8 band intensity. Data are presented as mean±sem. ***: p<0.001 versus basal conditions in nonstimulated PASMCs; ###: p<0.001 versus value for PASMCs isolated from controls stimulated with BMP4; ¶: p<0.05 versus value for PASMCs isolated from idiopathic PAH patients stimulated with BMP4. c) Relative densitometry ratio of phospho-Smad 1/5/8 in PASMCs isolated from defined naturally occurring BMPR-2 mutated patients (Δexon1, G/A = W16X, G/A = W70X, C/G = S107X, G/T = E195X, 22 bp del, T/C = S301P, C/T = R491W, C/T = Q495X). ░: nonstimulated; ▒: stimulated with 100 ng·mL−1 BMP4.

p38 mitogen-activated protein kinase (MAPK) phosphorylation induced by bone morphogenetic protein (BMP)4. 90% confluent pulmonary artery smooth muscle cells (PASMCs) isolated from controls (n = 10), pulmonary arterial hypertension (PAH) patients without (idiopathic PAH; n = 10) and with BMP receptor type 2 (BMPR-2) mutations (heritable PAH; n = 9) were stimulated with 100 ng·mL−1 BMP4 for 20 min, followed by lysis for total protein. a) Representative western blots for phospho-p38 and for β-actin to show equal loading. b) Densitometry of phospho-p38 and β-actin bands from western blots of control (□), idiopathic PAH (▪) and heritable PAH (▓) PASMCs. Results are presented as relative protein expression ratio of phospho-p38 and β-actin band intensity. Data are presented as mean±sem. ***: p<0.001 versus basal conditions in nonstimulated PASMCs; ###: p<0.001 versus value for PASMCs isolated from controls stimulated with BMP4; ¶: p<0.05 versus value for PASMCs isolated from idiopathic PAH patients stimulated with BMP4. c) Relative densitometry ratio of phospho-p38MAPK in PASMCs isolated from defined naturally occurring BMPR-2 mutated patients (Δexon1, G/A = W16X, G/A = W70X, C/G = S107X, G/T = E195X, 22 bp del, T/C = S301P, C/T = R491W, C/T = Q495X). ░: nonstimulated; ▒: stimulated with 100 ng·mL−1 BMP4.
Effects of BMP4 on PASMC proliferation induced by serum and PDGF treatment
PASMCs isolated from heritable and idiopathic PAH exhibited an increased proliferation, as assessed by [3H]-thymidine incorporation, in the presence of 10% serum but not 10 ng·mL−1 PDGF (fig. 7⇓). The addition of BMP4 induced an inhibition of [3H]-thymidine incorporation in both serum- and PDGF-treated PASMCs from idiopathic PAH and controls, but not in PASMCs from heritable PAH patients. The absence of BMP4-induced inhibition of proliferation was observed in PASMCs from all the heritable PAH patients, except in the patient with the 22 bp del mutation.

Basal [3H]-thymidine incorporation in pulmonary artery smooth muscle cells (PASMCs) derived from controls (□; n = 10), pulmonary arterial hypertension (PAH) patients without (▪; idiopathic PAH; n = 19) and with bone morphogenetic protein (BMP) receptor type 2 mutations (▓; heritable PAH; n = 9) in response to incubation with or without BMP4 (100 ng·mL−1), in the presence of 10% fetal calf serum (FCS) or 10 ng·mL−1 platelet-derived growth factor (PDGF). Data are presented as mean±sem. **: p<0.01 versus PASMCs isolated from controls; ***: p<0.001 versus PASMCs isolated from controls; #: p<0.05 versus basal conditions in BMP4-nonstimulated PASMCs; ###: p<0.001 versus basal conditions in BMP4-nonstimulated PASMCs.
Effects of BMP4 on PASMC apoptosis
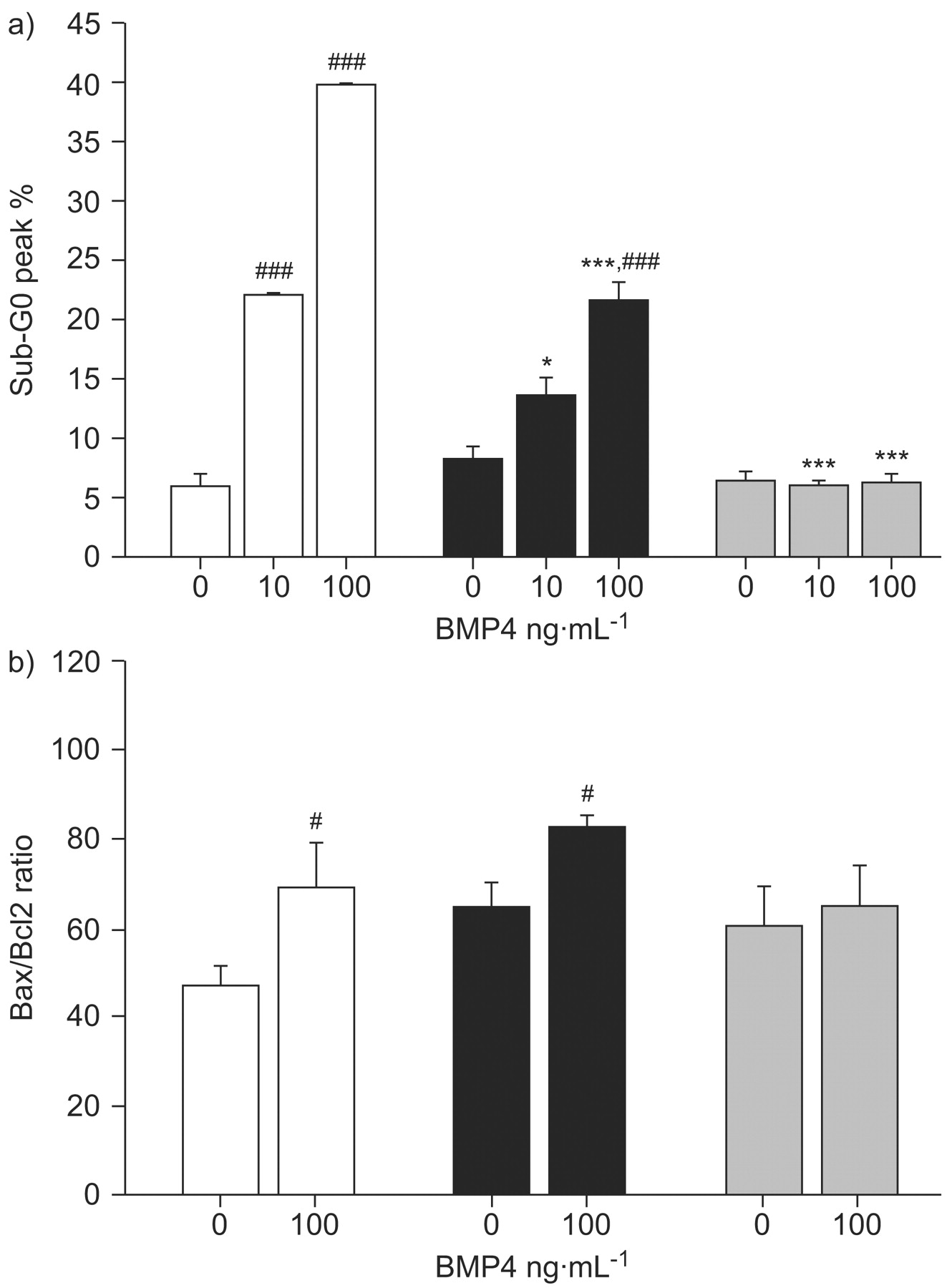
Flow cytometry analysis showed that BMP4 (10 and 100 ng·mL−1 for 24 h) increased apoptotic rates in PASMCs from idiopathic PAH patients and controls but to a lesser extent in PASMCs from idiopathic PAH and not from heritable PAH patients (fig. 8a⇓). The Bax/Bcl-2 pro-apoptotic ratio was increased by BMP4 at 100 ng·mL−1 in PASMCs from controls and idiopathic PAH, but not from heritable PAH patients (fig. 8b⇓).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bone morphogenetic protein (BMP)4-induced apoptosis of pulmonary artery smooth muscle cells (PASMCs) isolated from controls (□ n = 10), pulmonary arterial hypertension (PAH) patients without (▪ idiopathic PAH; n = 10) and with BMP receptor type 2 mutations (▓ heritable PAH; n = 9). a) Flow cytometric analysis of propidium iodide-stained PASMCs treated with 10 and 100 ng·mL−1 BMP4 to evaluate the distribution of cells in different phases of the cell cycle. Apoptotic cells are evaluated as percentage sub-G0/G1 phase of cell population. b) Apoptotic index (Bax/Bcl-2 ratio) of PASMCs that were seeded, synchronised for 48 h and treated with 100 ng·mL−1 BMP4. Relative mRNA expression (Bax/Bcl-2) ratio was determined by real-time quantitative PCR. Data are presented as mean±sem. *: p<0.05 versus similarly treated PASMCs isolated from controls; ***: p<0.001 versus similarly treated PASMCs isolated from controls; #: p<0.05 versus basal conditions in BMP4-nonstimulated PASMCs; ###: p<0.001 versus basal conditions in BMP4-nonstimulated PASMCs.
DISCUSSION
The present results show that 1) lung tissue and PASMCs, but not PECs, from heritable PAH patients present with decreased expression of BMPR-2 and variable increases in the expression of BMPR1-A and BMPR-1B, while only a (relatively less important) decreased expression of BMPR-2 is observed in PASMCs from idiopathic PAH patients; 2) PASMCs from heritable PAH patients show no BMP4-induced Smad 1/5/8 phosphorylation but do show BMP4-induced activation of the p38MAPK pathway; 3) PASMCs from PAH patients present with an in vitro proliferative and anti-apoptotic pattern, which can be inhibited by BMP4 in idiopathic PAH but not in heritable PAH; and 4) individual responses vary from one mutation to another, with in particular PASMCs from PAH patients with the 22 bp del mutation showing no difference compared with PASMCs from idiopathic PAH patients without identified mutations. The present work confirmed previous studies about BMPR-2 signalling in PAH and presented, for the first time, a large in vitro comparison of PASMCs with and without naturally occurring BMPR-2 mutations.
Although BMPR-2 mutations are identified in the majority of heritable PAH patients, and carry a significant risk of developing the disease in asymptomatic carriers, two large clinical studies have failed to disclose differences in clinical presentation, haemodynamics and histopathology between heritable and idiopathic PAH patients, except for an earlier onset of the disease, more compromised haemodynamics and maybe a less frequent reversibility at vasodilator testing in heritable PAH 7, 8. These observations suggest heterogeneous functional consequences of the various >200 BMPR-2 mutations hitherto reported, and also interactions with coexisting signalling abnormalities. In the present study, there was no difference in clinical presentation of heritable and idiopathic PAH, but this is probably related to minor phenotypic differences, individual variability and small sample size.
Our results confirm that the expression of BMPR-2 is decreased in idiopathic PAH and much more decreased in heritable PAH 21. The relative magnitudes of lung tissue and isolated PASMC and PEC expression were suggestive of a predominant PASMC location of BMPR-2. This is in contrast with predominant endothelial cell location previously reported in normal controls and in lungs from PAH patients 21. However, recent studies on human PASMCs and PECs showed relatively high levels of BMPR-2 on both cell types, but very low expression of BMPR-1A and -1B in PECs, consistent with a lack of BMP4 responsiveness 22. In the present study, the expression of BMPR-2, BMPR-1A and BMPR-1B was much lower in PECs compared with PASMCs, with no apparent impact of BMPR-2 mutations, so that further analysis focused on PASMCs. Moreover, in heritable PAH patients, the level of BMPR-2 expression was lower than predicted by the state of haploinsufficiency and the process of nonsense-mediated decay secondary to the presence of nonsense mutations. These observations are in keeping with the notion that some additional environmental and/or genetic factors may be responsible to further reduce BMPR-2 expression.
Decreased expression of BMPR-2 has also been reported to occur in experimental animal models of pulmonary hypertension, such as those induced by chronic systemic-to-pulmonary shunting 23, hypoxic exposure 24 and monocrotaline 25. In the latter study, decreased BMPR-2 expression was described in both lung tissue and PASMCs 25. Suprisingly, exclusive overexpression of BMPR-2 by gene therapy did not ameliorate monocrotaline-induced PAH in rats 26, indicating that reconstitution of the receptor was unable to restore BMP signalling and, thus, did not prevent disease onset or progression. The present results suggest that decreased expression associated with mutations in BMPR-2 seems to be crucial to explain the pro-proliferative and anti-apoptotic effects in PASMCs.
In the present study, the expression of BMPR-1A and BMPR-1B tended to increase, although quite variably, with significant increases of BMPR-1A in lung tissue of both heritable and idiopathic PAH patients but only in PASMCs from idiopathic PAH patients, and of BMPR-1B in PASMCs from heritable PAH patients. The expression of BMPR-1A was decreased in a study on patients with various causes of severe pulmonary hypertension, including PAH, but also mitral stenosis and thromboembolic pulmonary hypertension, and this was explained in relation to an overexpression of angiopoietin-1 27. The expression of BMPR-2 was unaltered in that study 27. We previously reported RTQ-PCR and western blotting of unchanged expression of angiopoietin-1 in whole lung tissue and in PASMCs from PAH patients 14. In monocrotaline-induced pulmonary hypertension, the expression of BMPR-2 and BMPR-1B was decreased, while the expression of BMPR-1A remained unchanged 25. An increased expression of BMPR-1B has been reported in one idiopathic PAH patient 28. The reasons for these discrepant results are unclear. Since BMPR-1A/BMPR-2 is the main BMP4-responsive receptor complex allowing for Smad 1/5/8 stimulation 22, the overexpression of BMPR-1A in the PASMCs of the idiopathic PAH patients could be somehow related to the normal BMP4 responsiveness of these cells. As for the overexpression of BMPR-1B in heritable PAH, this could be speculated to be related to alternative p38MAPK signalling activation.
Of the nine mutations identified in the present study, the W16X, R491W and Q495X have been previously reported 11, 29, 30. The six other mutations (Δexon1, W70X, S107X, E195X, 22 bp del and S301P), across all four domains of the receptor, are all novel, including one total deletion of an exon, three nonsense mutations, one partial deletion and one missense mutation. Previous studies showed that missense mutations in the ligand-binding domain by cysteine substitutions impair BMP signalling by mutant receptor mislocalisation in the cytosol 31. Moreover, noncysteine substitutions localise to the cell surface but also exhibit defects in BMP signalling activity 31. In contrast, mutations in the cytoplasmic C-terminal domain only moderately inhibit Smad signalling 31, 32. Therefore, in the present study, all reported mutations would be susceptible to be deleterious by changing the protein sequence at important functional sites of the receptor and the associated protein functions.
Mutations of BMPR-2 (including R491W) heterogeneously interfere with BMP downstream signalling, but all of them activate proliferative pathways 24. We previously reported that PASMCs isolated from idiopathic PAH patients present with enhanced growth responses to serum, but not to PDGF 10. In the present study, PASMCs with or without BMPR-2 mutations did not behave differently in this respect, but BMP4-induced growth inhibition and increased apoptosis was markedly more important in PASMCs with BMPR-2 mutations. It is of interest that all the BMPR-2 mutations identified in the present study, except the 22 bp deletion in the transmembrane domain, responded homogeneously to these effects of BMP4, supporting the notion that the majority of BMPR-2 mutations are functionally linked. Along the same line, decreased BMPR-2 expression in monocrotaline-induced pulmonary hypertension has been reported to be associated with decreased phosphorylation of Smad1 and a decrease in BMP-induced apoptosis of PASMCs 25.
In the present study, the application of BMP4 was associated in PASMCs with BMPR-2 mutations with a decreased activation (phosphorylation) of Smad 1/5/8 together with an increased activation (phosphorylation) of the p38MAPK pathway. We selected a dose of BMP4 of 100 ng·mL−1 on the basis of available literature 18, 19, 32 and preliminary testing showing maximum efficacy of 100 ng·mL−1 compared with 10 ng·mL−1 and 1 ng·mL−1 in discriminating PASMCs with and without BMPR-2 mutations. However, a recent study showed maximum efficacy at a lower dose of 10 ng·mL−1 of BMP4 33. The reasons for these discrepancies are not clear, and therefore the absence of complete dose–response curves may be a limitation to our findings. In monocrotaline-induced pulmonary hypertension, decreased BMPR-2 and phospho-Smad1 occurred without change in p38MAPK signalling in PASMCs 25. In hypoxia-induced pulmonary hypertension, downregulation of BMPR-2 did not affect Smad 1/5/8 phosphorylation, and was associated with decreased p38MAPK signalling 24. Although part of these differences may be related to model specificities and PASMCs versus whole lung measurements, the present data confirm previous reports 31, 32 that most mutations of BMPR-2 are associated with more profound changes in downstream signalling and are associated with increased p38MAPK signalling as a cause of increased PASMC proliferation.
The present results support the notion of altered BMPR-2/Smad signalling as a cause of increased proliferation of PASMCs playing an important role in the remodelling of pulmonary resistance vessels in PAH.
Support statement
This study was supported by grants from the Institut National de la Santé et de la Recherche Médicale, the Ministère de la Recherche, the Institut des Maladies Rares, the Délégation à la Recherche Clinique de l’Assistance Publique – Hôpitaux de Paris (all Paris, France), the Belgian Foundation for Cardiac Surgery and from the Fonds de la Recherche Scientifique Médicale (grant no. 3.4551.05; both Brussels, Belgium). This research project received financial support from the European Commission under the 6th Framework Programme (contracts LSHM-CT-2005-018275 and LSHM-CT-2005-018724, PULMOTENSION). This publication reflects only the authors’ views, and under no circumstances is the European Community liable for any use that may be made of the information it contains.
Statement of interest
None declared.
- Received December 2, 2008.
- Accepted March 12, 2009.
- © ERS Journals Ltd
References










