





Figures
- Fig. 2—
Hypoxia-inducible factor (HIF)-1-mediated oxygen sensing of transcription. The schematic illustrates how the α-subunit of the heterodimeric transcription factor HIF-1 conveys hypoxic conditions to transcriptional control. a) Under hypoxic conditions, HIF-1α dimerises with HIF-1β and binds to HIF-responsive target genes, such as erythropoietin or vascular endothelial growth factor. After recruitment of the transcriptional coactivator CREB (cAMP response element binding)-binding protein (CBP)/p300 to HIF-1α and additional binding of gene- or tissue-specific transcriptional cofactors, HIF-1-dependent transcription is induced. b) Under normoxic conditions, HIF-1α is absent because it is inactivated and immediately degraded by the proteasomal machinery. The oxygen sensors translating the oxygen concentration into changes in HIF-1α are oxygen-dependent hydroxylases: factor inhibiting HIF-1 (FIH-1) and prolyl hydroxylase domain-containing proteins 1–3 (PHDs). In the presence of oxygen, FIH-1 hydroxylates asparaginyl residue 803 (N) within the C-terminal transactivation domain of HIF-1α, preventing the recruitment of CBP/p300 to the HIF-1 complex, which leads to reduction of transcriptional activity. PHDs hydroxylate two prolyl (P) residues (P402 and P564) within the oxygen-dependent degradation domain of HIF-1α, allowing binding of the von Hippel–Lindau tumour suppressor protein (pVHL). pVHL functions as a recognition element for the multiprotein E3 ubiquitin ligase complex, which targets HIF-1α, via polyubiquitination, for proteasomal degradation. Whereas dimerisation and transactivation of the HIF-1 complex takes place in the cell nuclei, it is still under discussion whether the oxygen sensing process of FIH-1/PHDs, the binding of pVHL and the proteasomal degradation can take place in both cytoplasm and nucleus. OH: hydroxyl group.
- Fig. 3—
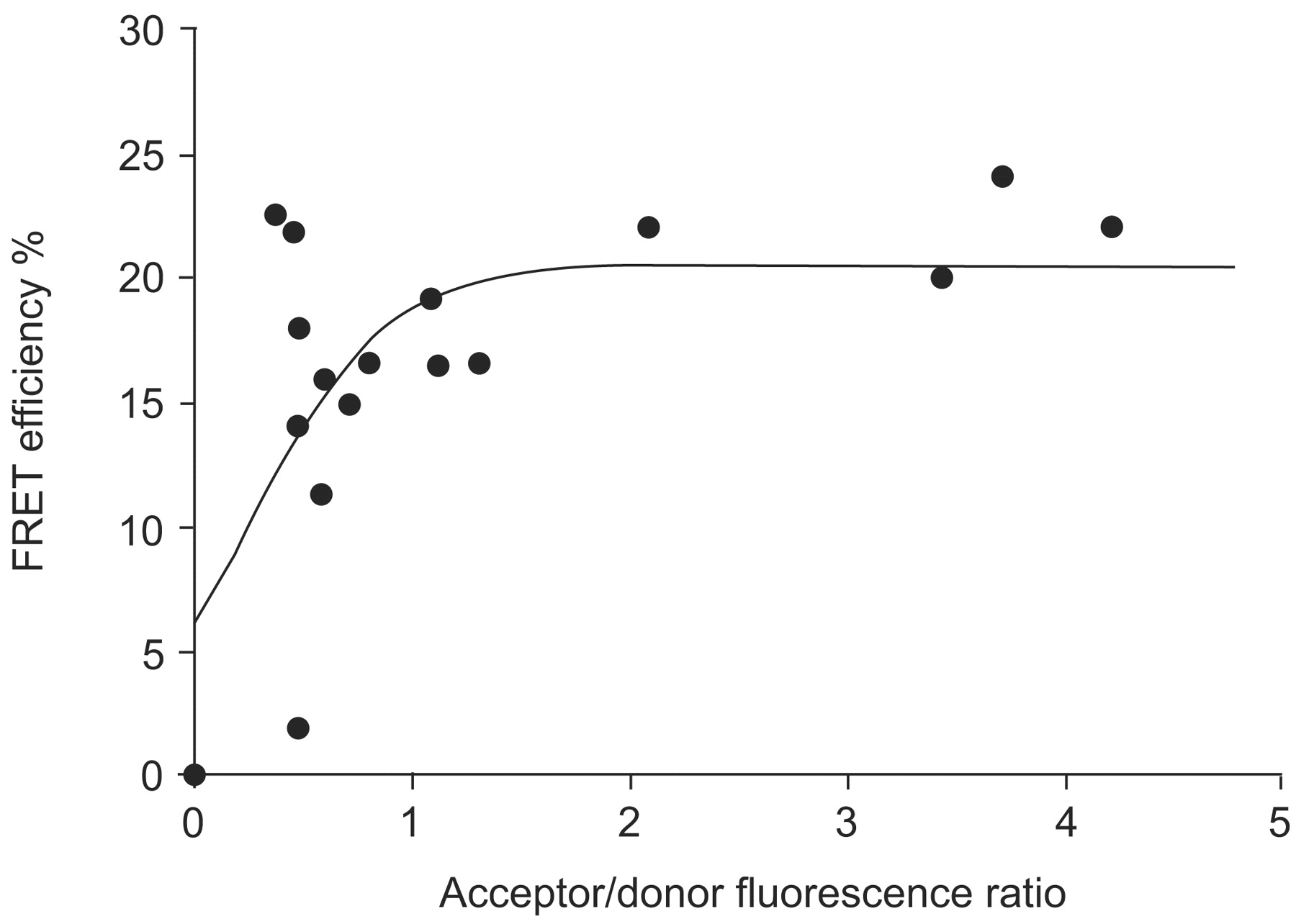
In vivo analysis of hypoxia-inducible factor (HIF)-1α and HIF-1β dimerisation using fluorescence resonance energy transfer (FRET). Enhanced cyan fluorescent protein (ECFP)-HIF-1α and enhanced yellow fluorescent protein (EYFP)-HIF-1β were coexpressed in U2OS osteosarcoma cell nuclei and imaged by two-photon laser scanning microscopy, while the cells were kept under hypoxia. According to the theory of FRET, energy from the donor molecule (ECFP-HIF-1α) is transferred to the acceptor (EYFP-HIF-1β) if the two molecules come closer than 10 nm. FRET efficiencies from cell nuclei were calculated using the acceptor bleaching method. The mean FRET efficiency of ECFP-HIF-1α and EYFP-HIF-1β interaction depends on their relative expression within the respective cell nucleus. Therefore, the distance between the molecules was determined (6.2 nm) when a plateau was reached 48.
- Fig. 4—
Subnuclear distribution of hypoxia-inducible factor (HIF)-1α and HIF-1β dimerisation, analysed by fluorescence resonance energy transfer (FRET). U2OS osteosarcoma cells were transfected with enhanced cyan fluorescent protein (ECFP)-HIF-1α and enhanced yellow fluorescent protein (EYFP)-HIF-1β. Under hypoxia, both fusion proteins were expressed in cell nuclei and FRET was imaged by laser scanning microscopy. The distribution of FRET efficiencies within a single cell nucleus was estimated using the sensitised FRET method 50 and visualised in false colour mode, as indicated by the colour bar. Subnuclear variations in FRET efficiency most likely reflect local variation in the mean distance between the ECFP- and EYFP-labelled HIF-1 subunits. Scale bar = 2 μm.



{kind=link}
{kind=link}
{kind=link}










