





Abstract
The aim of the present study was to assess the effects of possible interactions between β2-adrenoceptor gene polymorphisms and passive smoking on forced expiratory volume in one second (FEV1), forced vital capacity (FVC) and exhaled nitric oxide (eNO) in children aged 11 yrs.
A cross-sectional analysis of the longitudinal cohort was conducted for associations between β2-adrenoceptor gene polymorphisms and lung function and eNO with regard to passive smoking.
Among children exposed to tobacco smoke, those with Arg16 (at least one Arg allele) exhibited lower adjusted mean FEV1 (2.19 versus 2.38 L) and FVC (2.43 versus 2.64 L) than Gly16 homozygotes. Those with Gln27 (at least one Gln allele) also exhibited a lower adjusted mean FEV1 relative to Glu27 homozygotes (2.24 versus 2.39 L). Among children with no exposure to smoking, those with Arg16 or Gln27 showed lower adjusted geometric mean eNO levels compared with Gly16 homozygotes (15.4 versus 30.9 ppb) and Glu27 homozygotes (18.0 versus 49.7 ppb).
In conclusion, passive smoking had a significant effect on associations between β2-adrenoceptor gene polymorphisms and asthma-related phenotypes, enhancing the relationship between Arg16 and lung function and removing the relationship between Arg16 or Gln27 and exhaled nitric oxide levels.
The β2-adrenoceptor (β2-AR) is encoded by an intronless gene, located on chromosome 5q31–32 1, and contains several reported single nucleotide polymorphisms (SNPs) 2. The most commonly cited SNPs occur at nucleotide positions 46 (guanine/adenine) and 79 (cytosine/guanine), and result in the substitution of glycine (Gly) for arginine (Arg) at codon 16 (Arg16Gly) and glutamic acid (Glu) for glutamine (Gln) at codon 27 (Gln27Glu), respectively 3. Although the β2-AR gene (Β2AR) is not considered to be a major susceptibility gene for asthma 4, it has been suggested that Β2AR variant alleles may play a role in intermediate or asthma-associated phenotypes, such as response to medications 5, 6, airway hyperresponsiveness 7 and asthma severity 8. However, studies on the association between asthma-related phenotypes and Β2AR polymorphisms have reported inconsistent results 9. Environmental factors may interact with the Β2AR polymorphisms to affect asthma-associated phenotypes 7 or even asthma susceptibility 10. In terms of interactions between Β2AR and smoking, there is evidence that metabolites of the tobacco-specific nitrosamine stimulate the growth of human adenocarcinoma cells and are high-affinity agonists for β-ARs 11, 12. Moreover, a genomic screening study of inherited risk for asthma and bronchial responsiveness reported that the influence of susceptibility genes for asthma might not be apparent without the appropriate exposure to common environmental stimuli, such as passive smoking 13. Passive smoking exposure has been suggested to be a risk factor for the development of asthma 14. Since children are commonly exposed to cigarette smoke at home and in cars and public places, the interaction between cigarette smoke exposure and Β2AR polymorphisms is important for a better understanding of the association between these genetic variants and asthma-associated phenotypes. However, only a few studies 7, 10, 15 have investigated the interaction between cigarette smoking and Β2AR polymorphisms and none have examined children exposed to passive smoking.
An ongoing longitudinal study in Perth (Australia) measured lung function and airway responsiveness in a cohort between the ages of 1 month and 11 yrs 16, 17 and provided an opportunity to study the effects of interactions between Β2AR polymorphisms and passive smoking on asthma-associated phenotypes. The associations between Arg16 and decreased forced expiratory volume in one second (FEV1) at age 11 yrs in this cohort have been described previously 17. The complex relationships between exhaled nitric oxide (eNO), atopy and airway responsiveness have also been reported for this cohort 18. The aim of the present study was to analyse whether passive exposure to cigarette smoke might influence the association of Β2AR polymorphisms with both lung function and eNO. This is the first investigation of the consequences of possible interactions between Β2AR polymorphisms and passive smoking for lung function and eNO levels.
METHODS
Subjects
During the period 1987–1991, 253 Western Australian children were randomly and antenatally recruited from an unselected Caucasian population 19. They were all of European origin and born at full term, none showed major congenital anomalies and the infants were first assessed at the age of ∼1 month 16. At the age of 11 yrs, 194 children were followed up, with 180 children undergoing genotyping for Arg16Gly and Gln27Glu. Of these, 168 children underwent lung function measurement and 86 determination of eNO levels. The interaction between Β2AR polymorphisms and passive smoking was examined for the outcomes of lung function and eNO levels in this population.
Lung function, eNO and other asthma-associated phenotypes
Pulmonary function testing was performed using a spirometer (Pneumocheck Spirometer 6100; Welch Allyn, Skaneateles Falls, NY, USA) in accordance with published guidelines 20. Levels of eNO were measured using a fast-response chemiluminescence analyser, as previously described 18, 21. Airway responsiveness to histamine was evaluated using the rapid dosimeter technique 18, 22 and the dose–response slope (DRS) was determined. Skin-prick allergy tests were conducted against cow’s milk, egg white, ryegrass, mixed grass, Dermatophagoides farinae, D. pteronyssinus, cat dander, dog dander, Alternaria alternata and Aspergillus fumigatus, as described by Pepys 23. Atopy was defined as the presence of at least one positive skin-prick test result (with the longest dimension of the weal being ≥3 mm greater than that of the control). The eosinophil count was measured using a flow cytometer (Coulter Maxm; Beckman-Coulter Inc., Fullerton, CA, USA).
Genotyping of B2AR polymorphism
Blood was collected for DNA extraction on review at either age 6 or 11 yrs. SNPs at codons 16 and 27 were genotyped using techniques previously described in detail 17, 24. Haplotypes were constructed using the Bayesian statistic-based program PHASE, version 2.0.2 25.
Passive smoking assessment
In the study cohort, passive exposure to cigarette smoke was investigated on three occasions: on enrolment and on follow-up at both age 6 and 11 yrs. Specific questions related to maternal and paternal smoking during pregnancy were asked on enrolment. Information regarding current smoking, smoking history and smoking habit for parents and other smokers within households was also collected on enrolment and during follow-up at both age 6 and 11 yrs. Parents were asked the following questions with respect to cigarette smoke exposure. 1) Have you ever smoked cigarettes? 2) Are you a current smoker? 3) Do you now smoke cigarettes? 4) How old were you when you started smoking regularly? 5) If you stopped smoking completely, how old were you? 6) In addition to parents, were there any other smokers at home? 7) If a current smoker, when you are at home where do you smoke?
Based on the information on passive exposure to cigarette smoke collected on the three occasions, the children were divided into three groups: no exposure, significant exposure and unknown exposure. No exposure was defined as the child never having lived with a current smoker and their parents did not smoke during pregnancy. Significant exposure was defined as the child having lived with at least one smoker for >1 yr during infancy and childhood (up to the age of 11 yrs) and whose parents possibly smoked during pregnancy. The other children were defined as being of unknown exposure.
Statistical analysis
Both FEV1 and forced vital capacity (FVC) exhibited approximately normal distributions, and parametric analysis was employed directly. Levels of eNO and the confounders, DRS and eosinophil counts, were ln-transformed prior to parametric analysis. ANOVA was employed to compare the differences in FEV1, FVC and eNO level between genotypes of Arg16Gly and Gln27Glu in a univariate analysis. General linear models, adjusting for the confounders, were conducted to compare the adjusted/geometric mean FEV1, FVC and eNO levels both between Arg16 carriers (at least one Arg allele) and Gly16 between homozygotes, and between Gln27 carriers (at least one Gln allele) and Glu27 homozygotes. Age, sex and height were selected as confounders for lung function parameters and age, sex, height, atopy, DRS and eosinophil counts had been adjusted for eNO. Only the dominant effects of Arg in Arg16Gly and Gln in Gln27Glu were investigated in the current study because of the sample size. These models were fitted in the whole population, as well as separately in children with no exposure to tobacco smoke and with significant exposure to smoke. Data are presented as adjusted mean±sem or geometric mean (95% confidence interval (CI)). Linear regression was employed in order to identify associations of haplotypes of the Β2AR with FEV1, FVC and eNO, after adjusting for the confounders of interest. Significance was assumed at the 5% level.
RESULTS
Population characteristics
The study population included 180 children successfully genotyped for Β2AR polymorphisms and followed up at age 11 yrs. The characteristics of the population are shown in table 1⇓. In summary, the mean (95% CI) age of the population was 10.9 (10.8–11.1) yrs, with 77 (42.8%) being female. With regard to the Β2AR, 61.7% of the population had at least one Arg16 allele and 78.4% had at least one Gln27 allele. There was significant linkage disequilibrium between the Β2AR polymorphisms (p<0.001). Of the population, ∼46.1% was classified as having received no exposure to tobacco smoke and 37.2% as having received significant exposure. Of the 171 children, 89 (52%) were atopic, as defined by skin-prick test results at age 11 yrs. Lung function measurements were performed in 168 children and showed mean FEV1 and FVC values of 2.29 (2.23–2.36) and 2.51 (2.43–2.58) L, respectively. Levels of eNO were measured in 86 children, and the mean level was found to be 11.0 (9.1–13.2) ppb.
Characteristics of the population
Lung function and B2AR polymorphisms
Table 2⇓ shows the association between lung function parameters and Β2AR polymorphisms. On unadjusted univariate analysis, no significant differences in FEV1 and FVC were found between Arg16 (at least one Arg allele) and Gly16 homozygotes and between Gln27 (at least one Gln allele) and Glu27 homozygotes amongst the 168 children. After adjusting for age, sex and height, Arg16 was associated with significantly decreased FEV1 (p = 0.013) and FVC (p = 0.012) compared with Gly16 homozygotes. For lung function parameters expressed as a percentage of the predicted value, Arg16 was also associated with significantly decreased FEV1 (104 versus 108% pred; p = 0.027) and FVC (100 versus 104% pred; p = 0.018) compared with Gly16 homozygotes. The influence of Gln27Glu on lung function remained nonsignificant after this adjustment.
Lung function andβ2-adrenoceptor gene polymorphisms
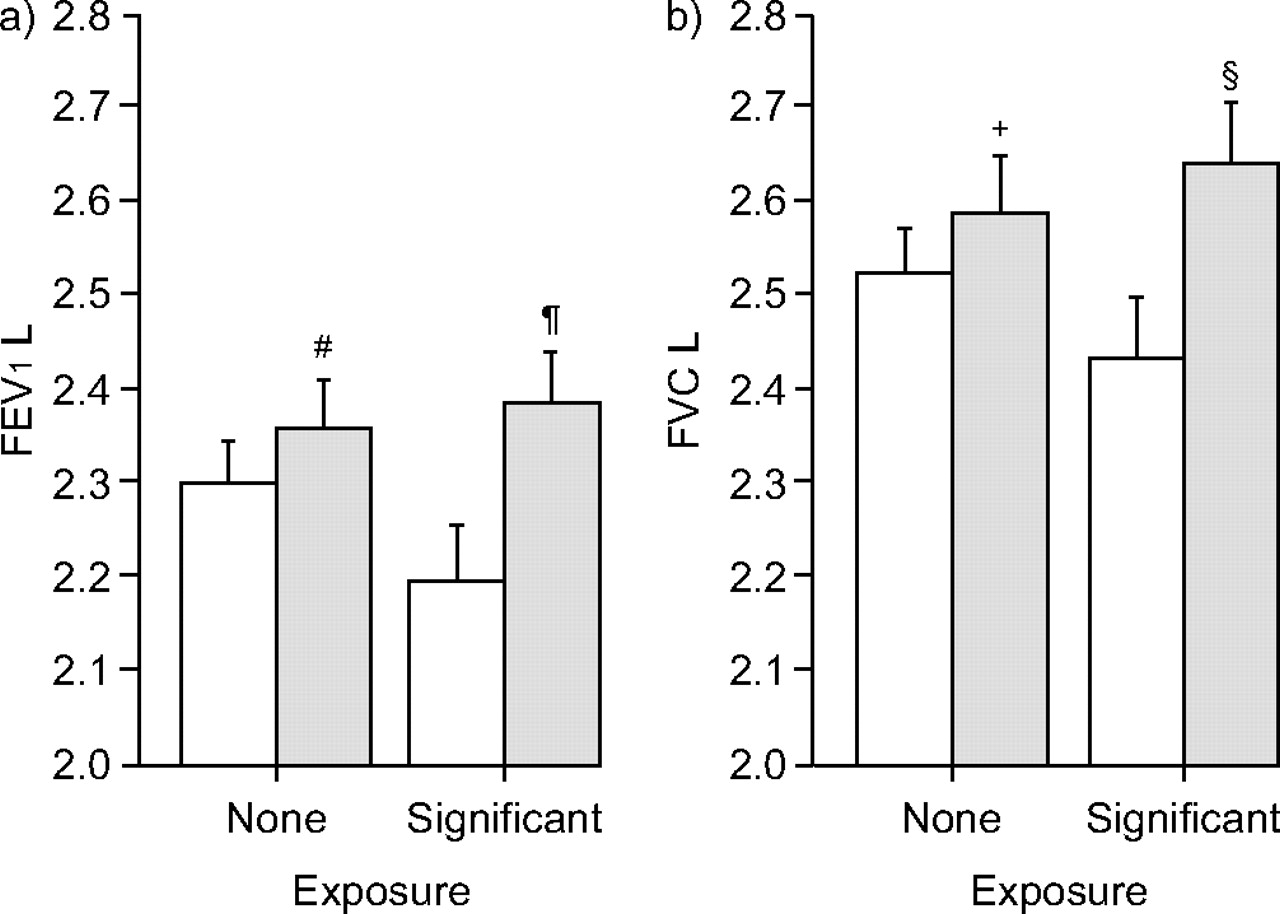
The relationship between Β2AR polymorphisms and lung function parameters (FEV1 and FVC) in children with no and significant exposure to tobacco smoke were analysed separately using general linear models. For the Arg16Gly polymorphism, Arg16 was associated with significantly lower adjusted mean FEV1 (2.19 versus 2.38 L; p = 0.006) and FVC (2.43 versus 2.64 L; p = 0.01) in children with significant exposure to tobacco smoke compared with Gly16 homozygotes (fig. 1⇓). Children with Gln27 exhibited a significantly lower mean FEV1 relative to Glu27 homozygotes (2.24 versus 2.39 L; p = 0.048) in those with significant exposure to tobacco smoke (fig. 2⇓). In children with no exposure to tobacco smoke, the association between lung function parameters and both Arg16 and Gln27 was not significant.

a) Forced expiratory volume in one second (FEV1); and b) forced vital capacity (FVC) in children with no or significant exposure to tobacco smoke: comparison between Arg16 (at least one Arg allele; □; no exposure: n = 53; significant exposure: n = 37) and Gly16 homozygotes (░; no exposure: n = 27; significant exposure: n = 30). General linear models were fitted to compare the difference in adjusted means. Data are presented as mean±sem. #: p = 0.34; ¶: p = 0.006; +: p = 0.35; §: p = 0.01 versus at least one Arg allele.

Forced expiratory volume in one second (FEV1); and b) forced vital capacity (FVC) in children with no or significant exposure to tobacco smoke: comparison between Gln27 (at least one Gln allele; □; no exposure: n = 68; significant exposure: n = 46) and Glu27 homozygotes (░; no exposure: n = 12; significant exposure: n = 21). General linear models were fitted to compare the difference in adjusted means. Data are presented as mean±sem. #: p = 0.12; ¶: p = 0.048; +: p = 0.29 versus at least one Gln allele.
eNO and B2AR polymorphisms
Of the children genotyped for Β2AR polymorphisms, 86 also underwent eNO level measurement. The concentrations of eNO in the different Β2AR genotype groups are shown in table 3⇓. Significant differences between the Β2AR Arg16Gly genotypes were found in eNO level in both the whole population (86 children; p = 0.017) and the 38 children with no exposure to tobacco smoke (p = 0.012).
Exhaled nitric oxide(eNO) levels in the different β2-adrenoceptor genotype groups
Children with Arg16 exhibited lower eNO levels than Gly16 homozygotes. The mean increase in eNO level in children with significant exposure to tobacco smoke was not significant. In addition, children with Gln27 exhibited lower eNO levels; this was also nonsignificant.
In order to adjust for covariates, general linear models were fitted for eNO level (the dependent variable) with either Arg16 or Gln27 as independent variable. The confounders included in the models for the adjustment were age, sex, height, atopy, DRS and eosinophil count. These confounding variables have previously been found to be significantly associated with eNO levels in children 18. After adjustment, children with Arg16 still showed a significantly lower eNO level than Gly16 homozygotes (p = 0.001). Furthermore, children with Gln27 also showed significantly lower eNO concentrations than Glu27 homozygotes (p = 0.018).
Figure 3⇓ shows the adjusted geometric mean eNO level by genotype in children with no and significant exposure to tobacco smoke. As found in the whole population, Arg16 was associated with significantly lower eNO levels compared with Gly16 homozygotes (15.4 versus 30.9 ppb; p = 0.01) in children with no exposure to tobacco smoke. Gln27 was significantly associated with decreased eNO levels in children with no exposure to tobacco smoke compared with Glu27 homozygotes (18.0 versus 49.7 ppb; p = 0.001). However, the decreases in eNO level for Arg16 and Gln27 were not significant in children with significant exposure to tobacco smoke.

{kind=link}
{kind=link}
{kind=link}
Exhaled nitric oxide (eNO) in children with no or significant exposure to tobacco smoke. Comparison between: a) Arg16 (at least one Arg allele; □; no exposure: n = 25; significant exposure: n = 20) and Gly16 homozygotes (░; no exposure: n = 13; significant exposure: n = 15); and b) between Gln27 (at least one Gln allele; □; no exposure: n = 31; significant exposure: n = 25) and Glu27 homozygotes (░; no exposure: n = 7; significant exposure: n = 10). General linear models were fitted to compare the difference in adjusted geometric means. Data are presented as adjusted geometric mean and upper 95% confidence interval. #: p = 0.01 versus at least one Arg allele; ¶: p = 0.20; +: p = 0.001 versus at least one Gln allele; §: p = 0.028.
B2AR polymorphism haplotypes
Four haplotypes were constructed for the two SNPs of the Β2AR, namely the Arg/Glu haplotype (ARGGLU), ARGGLN, GLYGLU and GLYGLN. ARGGLU was rare because of the linkage disequilibrium between the two SNPs; therefore, only the other three haplotypes were further investigated for association with lung function and eNO level stratified by passive smoking exposure. The frequencies of ARGGLN, GLYGLU and GLYGLN in the population were 37.5, 41.7 and 17.5%, respectively.
Table 4⇓ shows the association of haplotypes of the Β2AR with FEV1, FVC and eNO in children with and without exposure to passive smoking. Among children with no exposure, those with GLYGLN exhibited significantly higher FEV1 and FVC (p = 0.009 and 0.002, respectively). Moreover, those with ARGGLN exhibited lower eNO levels (p = 0.01) and those with GLYGLU higher eNO levels (p = 0.04). Among children with significant exposure, those with ARGGLN showed significantly lower FEV1 (p = 0.011) and FVC (p = 0.021). Those with GLYGLU showed relatively higher FEV1 (p = 0.024). Acknowledging the small sample size in children with GLYGLN, the relationships between GLYGLN and both lung function parameters and eNO were re-examined in linear regressions by grouping children with either one or two copies of GLYGLN together. GLYGLN was still significantly associated with FEV1 and FVC in children with no exposure.
Association ofβ2-adrenoceptor gene haplotypes with forced expiratory volume in one second (FEV1), forced vital capacity (FVC) and exhaled nitric oxide (eNO)#
DISCUSSION
The present study showed significant effects of passive smoking on the association of Β2AR polymorphisms with lung function and eNO. In children with exposure to tobacco smoke during pregnancy, infancy and childhood, Arg16 and Gln27 were associated with a decrease in FEV1 and FVC (which was not significant for Gln27). In children with no exposure, the effects of Arg16 and Gln27 on lung function were not significant. Arg16 and Gln27 carriers showed significantly decreased eNO levels, with the association more apparent in children with no exposure to tobacco smoke, suggesting that smoke exposure obviated the difference between the alleles. The haplotype analyses further confirmed the modifying effects of passive smoking on the associations.
Previous studies have found that Β2AR polymorphisms are of functional importance 2. The relationship between Arg16 and increased bronchial hyperresponsiveness at age 1 month and decreased lung function at age 11 yrs have previously been reported 17. The associations between bronchial hyperresponsiveness and lung function and haplotypes of the Β2AR have also been investigated 26. The present study clarified the associations of the Β2AR gene and lung function and eNO, particularly in children with exposure to the environmental factor of passive smoking.
There have been two epidemiological studies reporting the interaction between Β2AR polymorphisms and smoking. The characteristics and findings of the two studies, as well as the present study, are summarised in table 5⇓. There are significant differences in the characteristics of the three populations in that the current study involved passive smoking in children, whereas the other two analysed active smoking in adults. The ethnic background of the cohorts and observed outcomes also differ between the three studies.
Characteristics and findings of the three studies on interactions betweenβ2-adrenoceptor gene polymorphisms and smoking
Wang et al. 10 found that there was a significant dose–response relationship between tobacco smoke (cumulative consumption in pack-yrs) and risk (odds ratio) of asthma in Arg16 homozygotes in a middle-aged Chinese population. In that study, lower predicted FEV1 was one of the important discriminating criteria between asthmatic and control subjects. The findings of the present study are consistent with the report of Wang et al. 10 in that there was an interaction between Arg16 and smoking with respect to lung function. The marginally significant finding that Gln27 was associated with decreased FEV1 in children with significant exposure to smoking in the present study may possibly be due to linkage disequilibrium between the Arg16Gly and Gln27Glu SNPs. Litonjua et al. 7 did not find an association between Arg16 and airway responsiveness in ever-smokers in a relatively older white male population. They attributed their contradictory findings to the differences in ethnicity, sex and asthma phenotypes when their cohort was compared to that of Wang et al. 10. The present study also found that Arg16 and Gln27 were associated with significantly decreased eNO levels. Although the phenotypes and smoking exposure type were different from those reported by Litonjua et al. 7, the results are consistent in that the association was more apparent with no exposure to smoke. Litonjua et al. 7 suggested that the stronger association in nonsmokers resulted from smoke exposure overwhelming any effects of Β2AR variants in ever-smokers. However, this is unlikely to be the explanation in the present study as no significant difference was found in eNO levels between children with no and significant exposure to smoking. Wang et al. 10 reviewed the effects of tobacco smoke on the Β2AR density, as reported by Laustiola and co-workers 27, 28, and speculated that the smoking-induced downregulation of the Β2AR might differ in subjects carrying different polymorphic forms of this receptor 10. They suggested that this spectrum of gene–exposure interactions might contribute to variable expression of the asthma-related phenotypes 10. The complex interactions between Β2AR polymorphisms, passive smoking, lung function, eNO and asthma susceptibility require further elucidation.
Levels of eNO have been shown to be raised in atopy 18, 29, 30. Although inconsistently, exposure to environmental tobacco smoke appears to reduce eNO levels 31–34. In the present study, the unique finding of the association between Β2AR polymorphisms and eNO levels was independent of several asthma-associated phenotypes. Increased nitric oxide (NO) levels are the result of enhanced expression of NO synthases in various cells in the lung, including alveolar macrophages, airway smooth muscle and airway epithelial cells 35. No significant evidence supports a direct association between the Β2AR and NO synthases; however, there are several possible indirect links that have been suggested. First, NO synthases may be transcriptionally regulated in response to cytokines 36, 37, and cytokines may be regulated or inhibited by Β2AR 38, 39. Secondly, NO, as a messenger molecule, is related to endothelial mechanisms of vasodilation due to the stimulation of the Β2AR 40. It has been suggested that NG-monomethyl-l-arginine inhibits the response to β-agonists by activation of the endothelial l-arginine/NO pathway 40. These possible relationships between the Β2AR and eNO may partly explain the variations in eNO level associated with different Β2AR polymorphisms. Recognising the possible effects of passive smoking on eNO, it is possible that exposure to passive smoking may modify the effects of Β2AR polymorphisms on eNO.
The present study has certain limitations. Genotyping was only performed in 180 out of the 253 children recruited at the beginning of the study. Among the 180, some did not undergo lung function tests or measurements of eNO at age 11 yrs. In addition, males were dominant in the population. Analysis of only those subjects with complete information as regards genotyping, lung function or eNO may have introduced bias to the findings. Missing data and sex disproportions may have resulted in the population not being representative in general. Stratification by passive smoking further decreased the sample size for the linear regression analysis conducted in each group. Therefore, the findings should be interpreted with caution. However, no significant differences were found in terms of asthma, atopy and other asthmatic symptoms between the population studied and those lacking genotyping or lung function or eNO measurements. This suggests that the study population is likely to be representative of the whole cohort. Although passive smoking exposure was assessed several times using questionnaire surveys, no biomarkers of smoking exposure were measured due to financial and other constraints. Possible parental recall biases may thus have distorted the findings. However, as recently reported, questionnaire assessment reliably discriminates between different levels of passive smoking exposure in children 41. In addition, an unknown exposure group was defined to circumvent the possible uncertainty of smoking exposure in children. Those children with unknown exposure to cigarette smoke were not included in the analysis of the interaction between Β2AR polymorphisms and passive smoking.
In conclusion, it has been shown that β2-adrenoceptor gene polymorphisms are associated with decreased lung function in children significantly exposed to cigarette smoke and with decreased exhaled nitric oxide levels in unexposed children. There was significant interaction between the β2-adrenoceptor gene single nucleotide polymorphisms studied (Arg16Gly and Gln27Glu) and passive smoking in respect of lung function and exhaled nitric oxide in the population.
Acknowledgments
The authors are grateful to the many colleagues who contributed to the present cohort study, and acknowledge the important contributions of S. Young, who recruited the cohort, and V. Judge, P. Rye and L. Palmer, who undertook the assessment of the cohort at age 6 yrs.
Footnotes
-
For editorial comments see page 3.
- Received September 20, 2006.
- Accepted March 23, 2007.
- © ERS Journals Ltd
References










