





Abstract
Tumour necrosis factor (TNF)-α is associated with childhood wheezing. A genetic predisposition to increased TNF-α production, influenced by single nucleotide gene polymorphisms, may be important.
Frequencies of TNF-α-308G/A and lymphotoxin (LT)-α+252G/A polymorphisms were compared in 115 asthmatic children, 55 wheezy infants and 156 control school children from the UK. Genotype frequencies for the TNF-α-308 and LT-α+252 polymorphisms were significantly different from controls.
Haplotype analysis showed that TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252A was associated with a markedly increased risk for both asthma and infant wheezing. The TNF-α-308G, LT-α+252G/TNF-α-308G, LT-α+252A combination was protective for asthma and infant wheezing. These findings were confirmed by analysis of Caucasian data. Nasal TNF-α levels were measured in the infants during acute wheezing episodes and higher, but nonsignificant levels were produced in those with one or two LT-α+252A alleles. Unexpectedly, significantly lower nasal TNF-α levels were found in the presence of one or two TNF-α-308A alleles. TNF-α-308/LT-α+252 genotype combinations had a significant influence on nasal TNF-α levels.
In conclusion, these findings may have implications for future early intervention studies by helping to identify infants at increased risk for wheezing and childhood asthma.
It is well established that airway inflammation is of paramount importance in the pathophysiology of asthma and infant wheezing, and in particular during viral-induced episodes 1. Some children mount an exaggerated inflammatory response and it is possible that they are genetically predisposed to do so. A number of cytokines participate in the inflammatory cascade, however, the amount of each produced is quite variable amongst individuals. One example is tumour necrosis factor (TNF)-α, which is a pro-inflammatory cytokine that has been implicated in both asthma 2 and respiratory viral infections 1. In one study, nasal TNF-α levels were significantly increased in infants during acute, wheezy episodes associated with respiratory tract infections, but levels varied as much as 400-fold between individuals 3. Furthermore, in vitro work has shown marked inter-individual differences in TNF-α production by peripheral blood mononuclear cells taken from healthy subjects, however the production in the same subject was highly reproducible 4.
The TNF-α gene is located within the class III region of the major histocompatibility complex (MHC) close to the HLA-B locus on chromosome 6p. A polymorphism in the promoter region of the TNF-α gene at nucleotide −308, relative to the transcription start site, may be important in determining the host TNF-α response 5. It is important to note that although traditionally this polymorphism is said to be at the −308 position, it has since been pointed out that there was an initial misnumbering and this polymorphism is in fact sited at nucleotide -307, relative to the transcription start site 6, 7. However, most of the literature has stuck to the 308 nomenclature, as have the current authors for the sake of clarity. There are two alleles at the polymorphic site, TNF-α-308G (TNF1) and TNF-α-308A (TNF2). In normal populations, TNF-α-308G homozygosity is the predominant genotype. A number of studies have tried to determine whether the polymorphism influences TNF-α expression or susceptibility to certain diseases. Results are conflicting, but a recent review suggested that the TNF-α-308G/A polymorphism does have a small, but significant, functional effect, with the A allele being associated with higher constitutive and inducible levels of transcription for TNF-α than the G allele 7.
Within the TNF gene complex, in the first intron of the lymphotoxin α (LT-α; TNF-β) gene, there is an NcoI restriction polymorphism. This is known as TNFB*1, or LTB*1, in the presence of the restriction site and TNFB*2, or LTB*2, in its absence 8. In strong linkage disequilibrium with the TNFB*1 and TNFB*2 alleles there is a second variant, a noncoding G/A polymorphism, at position +252 after the transcription initiation site, known as LT-α+252. LT-α+252G corresponds to TNFB*1, and LT-α+252A corresponds to TNFB*2 9. Again, studies on the functional effect of this polymorphism are conflicting, with TNF-α expression said to be either increased in association with TNFB*2 8, 10 or not associated with it 9.
There have been numerous studies into disease associations with these TNF polymorphisms, which have been summarised by Haukim et al. 11. As regards their association with respiratory disease, the TNF-α-308A allele (either in the homozygous or heterozygous state) has been found to be associated with an increased risk of developing chronic bronchitis in adults 12, and with increased disease severity in children with cystic fibrosis 13. Work in asthma in various ethnic groups has, so far, produced conflicting results with studies showing an association with TNF-α-308A 14–19 and TNF-α-308G 20, 21, or none at all with the TNF-α-308 locus 22–28; with LTB*1 14, 15, LTB*2 20, 29 or none with the LT-α locus 18, 22, 23, 25, 29–31. The aim of the present study was to investigate the genetics of TNF-α production in terms of the TNF-α-308 and LT-α+252 alleles, and their relationship to childhood asthma and infant wheezing. In addition, in the wheezy infants, a novel attempt was made to relate the genotype to the functional phenotype, namely nasal TNF-α levels measured during an acute wheezing episode.
METHODS
Subjects
Asthmatic children aged between 2–17 yrs were recruited from respiratory clinics in two London hospitals 1 mile apart. Clinical diagnosis of asthma had been confirmed in all subjects by the same respiratory paediatrician. The subjects were subclassified as: 1) severe asthma if they were taking ≥800 mcg·day−1 inhaled corticosteroids (beclomethasone/budesonide equivalent dose), or 2) mild/moderate asthma if taking doses <800 mcg·day−1. Infant wheezers were children <2-yrs old admitted to one of the hospitals with an acute wheezing illness. Normal controls were children aged >8 yrs who were attending one of four local schools (<1 mile away) and who had no doctor-diagnosis of asthma.
Sampling
DNA was collected from all subjects using CEP buccal swabs (Life Technologies Ltd., Paisley, Scotland) to gently scrape the inside of the mouth. Nasal lavage was performed on the wheezy infant group using a previously described method, whereby 2 mL phosphate buffered saline, at room temperature, was instilled into each nostril while simultaneously aspirating the fluid back from the anterior nares using a soft rubber Jaques catheter 32. Inulin (0.45 g·L−1) was added to the nasal wash as a marker of dilution so that nasal TNF-α levels could be expressed in pg·mL−1 nasal lining fluid 32. The wheezy infants also had nasopharyngeal aspirate collected using standard methods for detection of respiratory syncytial virus (RSV), as part of the normal clinical management.
Cytokine polymorphism analysis
Genomic DNA was extracted from buccal swabs using 20% chelex 100, incubated at 56°C for 4 h, boiled for 8 min and then centrifuged at 13,000×g. DNA was stored at −20°C. Cytokine polymorphisms in genomic DNA were analysed using PCR with sequence specific primers (SSP). The primers and protocols used were based on those described by Perrey et al. 33. The primers were designed to be of ∼20 base pairs (bp) in length containing the known polymorphisms at the 3′ end, in order to yield a product between 250–300 bp and to be of similar G and C content to ensure efficient priming. Human growth hormone was used as internal control for the PCR-SSP. Following PCR amplification, the products were size separated by electrophoresis (280 V for 20 min) on a 2% agarose gel. The PCR products were stained with ethidium bromide (0.5 mg·mL−1) and visualised by ultraviolet chemi-luminescence.
Nasal tumour necrosis factor-α
The nasal lavage fluid was centrifuged at 3,000×g for 20 min, within 10 min of collection, and stored at −80°C. Nasal TNF-α was measured in the supernatant by ELISA, using a commercial Quantikine Human TNF-α immunoassay kit (R&D Systems Europe Ltd, Abingdon, UK). Manufacturers' instructions were carried out and the lower limit of detection was 20 pg·mL−1. Inulin was measured by spectrophotometry using an adaptation of the method described by Heyrovsky 34.
Respiratory syncytial virus
Identification of RSV was determined by standard direct immunofluorescence and viral culture.
Statistics
Allele frequency was calculated as follows:  Hardy-Weinberg equilibrium was calculated for genotypes in the controls using the standard equation with a Chi-squared goodness-of-fit test. The comparison of allele and genotype frequencies between patient groups, and the case control odds ratios (OR) with the 99% confidence intervals (CI), were calculated using the Chi-squared test with Yates's correction. Haplotype analysis was performed on PHASE software V2.1 (www.stat.washington.edu/stephens) using the -C and F0 flags with the best pair (0) output for haplotype determination. This programme implements a Bayesian statistical method for reconstructing haplotypes from population genotype data 35, 36. Bayes' theorem was applied as per Altman 37. Nasal TNF-α levels were compared between patient groups and controls using nonparametric analysis, Mann-Whitney U-tests and Kruskal-Wallis tests since data were not normally distributed. Values of p<0.01 were considered statistically significant, with 99% CI used as a compromise between multiple contrasts producing spurious statistical associations and missing clinically-relevant associations. Power studies were performed in advance. Assuming a background homozygosity rate of 7% for TNF-α-308A, to detect a 20% prevalence in a study group with an 85% certainty at the 5% significance level, required a total cohort of 240 subjects.
Hardy-Weinberg equilibrium was calculated for genotypes in the controls using the standard equation with a Chi-squared goodness-of-fit test. The comparison of allele and genotype frequencies between patient groups, and the case control odds ratios (OR) with the 99% confidence intervals (CI), were calculated using the Chi-squared test with Yates's correction. Haplotype analysis was performed on PHASE software V2.1 (www.stat.washington.edu/stephens) using the -C and F0 flags with the best pair (0) output for haplotype determination. This programme implements a Bayesian statistical method for reconstructing haplotypes from population genotype data 35, 36. Bayes' theorem was applied as per Altman 37. Nasal TNF-α levels were compared between patient groups and controls using nonparametric analysis, Mann-Whitney U-tests and Kruskal-Wallis tests since data were not normally distributed. Values of p<0.01 were considered statistically significant, with 99% CI used as a compromise between multiple contrasts producing spurious statistical associations and missing clinically-relevant associations. Power studies were performed in advance. Assuming a background homozygosity rate of 7% for TNF-α-308A, to detect a 20% prevalence in a study group with an 85% certainty at the 5% significance level, required a total cohort of 240 subjects.
RESULTS
Subjects
In total, 115 asthmatic children (88 mild/moderate, 27 severe), 55 infant wheezers and 156 controls were enrolled. In the study, as expected, there were age differences between the subject groups, but there was no difference in sex (table 1⇓).
Subject's characteristics
Tumour necrosis factor gene polymorphisms
Polymorphism data for TNF-α-308 were unavailable on 7% wheezing children and 4% controls, and for LT-α+252 on 12% wheezing children and 5% controls. Missing data were due to technical reasons relating to DNA extraction or failed PCR.
Allele frequencies for TNF-α-308G/A were not significantly different for either asthmatics or infant wheezers compared with controls (p = 0.04). The allele frequencies for LT-α+252G/A were not significantly different for asthmatics (p = 0.04), but were for infant wheezers (p<0.01) (table 2⇓). There were no differences when comparing severe with mild/moderate asthma.
Allele frequencies
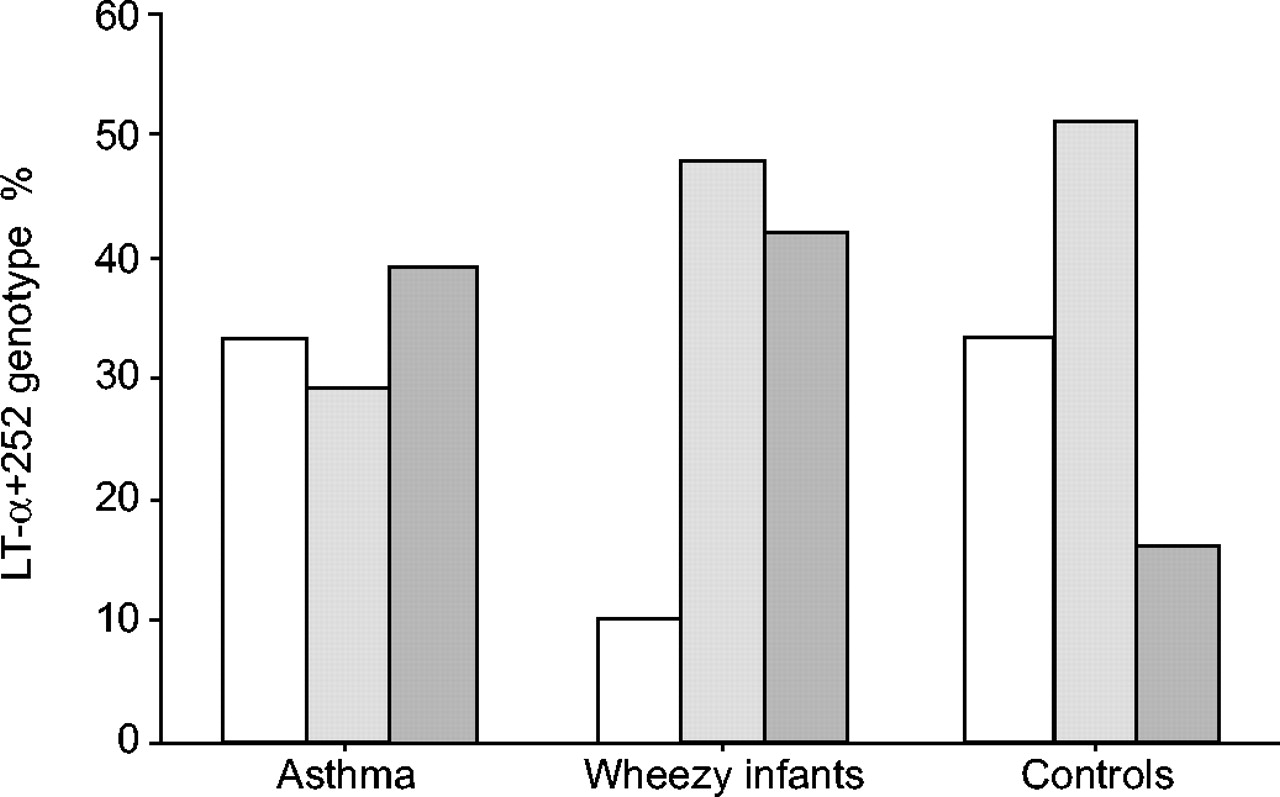
Genotype frequencies for the TNF-α-308 polymorphism for asthmatics and infant wheezers were significantly different from controls (p<0.001; table 3⇓; fig. 1⇓). Severe asthma was not different from mild/moderate asthma. The genotype frequencies for LT-α+252 polymorphism showed that asthmatics and infant wheezers were significantly different from the controls (p<0.001), and again severe asthma was not different from mild/moderate asthma (table 3⇓; fig. 2⇓). Genotype distributions were found to be in Hardy-Weinberg equilibrium in the controls. Linkage disequalibrium (LD) was calculated and there was evidence of LD between the two loci studied, with a D prime of 0.5 in the subjects and controls (subgroup analysis revealed D prime to be 0.31, 0.52 and 0.88 for asthmatics, wheezy infants and controls, respectively).

Genotype frequencies (%) for tumour necrosis factor (TNF)-α-308 polymorphisms in asthma (n = 108), wheezing infants (n = 50) and controls (n = 149). Frequencies were significantly different from controls (p<0.001). □: GG; ░: GA;  : AA.
: AA.

Genotype frequencies (%) for lymphotoxin (LT)-α+252 polymorphism in asthma (n = 101), wheezing infants (n = 48) and controls (n = 148). Frequencies were significantly different from controls (p<0.001). □: GG; ░: GA;  : AA.
: AA.
Genotype frequencies
Haplotype analysis using a permutation test demonstrated significant differences in haplotype frequencies between asthmatics, wheezy infants and controls, with a global p-value of <0.01 in both the whole sample and Caucasians only (table 4⇓).
Haplotype analysis demonstrating significant differences** in haplotype frequencies between asthmatics, wheezy infants and controls
In terms of individual haplotypes (table 5⇓), the largest effect was seen with the combination TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252A, where there was a markedly increased OR for both asthma (OR: 13.7; 99% CI: 2–94) and infant wheezing (OR: 12.5; 99% CI: 2–103) compared with controls. Haplotype analysis of TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252A for the risk of asthma or infant wheezing had a high specificity (100%), but a low sensitivity (17%) with a positive predictive value (PPV) of 100% and a negative predictive value (NPV) of 54%. Using Bayes' theorem and assuming a disease prevalence of 30% for both conditions in the general population, the overall PPV for disease, given the gene profile, was 100% and the NPV 74%. Assuming 20% disease prevalence, the PPV remained 100% whilst the NPV increased to 82%.
Inferred haplotype comparing asthma and infant wheezing with controls in the entire study group
Introducing an LT-α+252 G allele (to produce TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252G) counterbalanced the wheeze-promoting effect of TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252A leading to a nonsignificant OR for asthma. Although, there was still a significant effect on wheezy infants (OR: 3.0; 99% CI: 1.2–7.6). The introduction of a further second TNF-α-308 G allele (TNF-α-308G, LT-α+252G/TNF-α-308G, LT-α+252A) had an even greater counterbalance. This produced a protective effect with a reduced OR for asthma of 0.3 (99% CI: 0.1–0.7) and infant wheezing of 0.2 (99% CI: 0.04–0.7). For some of the combinations, numbers were too small to make valid comparisons.
Individual haplotypes with significant ORs were then analysed using data for Caucasian subjects only. The previous positive findings were confirmed in this subgroup (table 6⇓).
Inferred haplotype comparing asthma and infant wheezing with controls in Caucasians
Nasal tumour necrosis factor-α
In total, there were 55 infant wheezers who each had a nasal lavage. Of which, 32 were RSV-positive and 23 RSV-negative. There was no age difference between the two groups (median 5 versus 6 months, respectively). Nasal TNF-α levels were not significantly affected by RSV status, with median (interquartile ranges) of 697 (424–2187) in the RSV-positive infants versus 359 (101–1176) pg·mL−1 nasal lining fluid in the RSV-negative infants (p = 0.12).
Data on TNF-α-308 polymorphisms were only available on 50 out of the total 55 infant wheezers and LT-α+252 on 48 out of 55 for technical reasons. Nasal TNF-α levels were significantly higher in those with TNF-α-308GG genotype compared with TNF-α-308GA (p = 0.007; table 7⇓; fig. 3⇓). There were only two infants with the TNF-α-308AA genotype, but combining them with the heterozygotes confirmed that the presence of one or two A alleles led to significantly lower nasal TNF-α levels (p = 0.006). There was a trend to higher nasal TNF-α levels in those with the LT-α+252GA genotype compared with LT-α+252GG (p = 0.07), and a trend to higher levels in the presence of one or two A alleles (p = 0.08). However, overall differences were not significant (Kruskal-Wallis p = 0.16; fig. 4⇓). Analysing TNF-α-308/LT-α+252 genotype combinations showed that there was a significant influence on nasal TNF-α levels (Kruskal-Wallis p = 0.007; fig. 5⇓).

Nasal tumour necrosis factor (TNF)-α levels (pg·mL−1 nasal lining fluid: NLF) related to TNF-α-308 genotype in wheezing infants (n = 50). Levels were significantly higher in TNF-α-308GG compared with the TNF-α-308GA genotype (p = 0.007). The y-axis is a logarithmic scale. The horizontal line represents the median, and the vertical line represents the interquartile range, for each group of data. #: p = 0.007.

Nasal tumour necrosis factor (TNF)-α levels (pg·mL−1 nasal lining fluid: NLF) related to lymphotoxin (LT)-α+252 genotype in wheezing infants (n = 48). The y-axis is a logarithmic scale. The horizontal line represents the median, and the vertical line represents the interquartile range, for each group of data. #: p = 0.07. Differences were not statistically significant (Kruskal-Wallis p = 0.16), although there was a trend to lower levels in LT-α+252GG compared with the LT-α+252GA genotype (p = 0.07).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nasal tumour necrosis factor (TNF)-α levels (pg·mL−1 nasal lining fluid: NLF) related to TNF-α-308/lymphotoxin (LT)-α+252 genotype combinations in wheezing infants (n = 46). The horizontal line represents the median, and the vertical line represents the interquartile range, for each group of data. TNF-α levels were significantly different for the various combinations (Kruskal-Wallis p = 0.007). The y-axis is a logarithmic scale.
Nasal tumour necrosis factor-α levels in wheezing infants related to TNF gene polymorphisms
DISCUSSION
A definite relationship was found between two single nucleotide gene polymorphisms in the TNF gene complex and childhood wheezing in a mixed UK population. In keeping with some 14–19, but not all 20–26, 28 studies, TNF-α-308A was associated with childhood asthma. It was also found to be associated with infant wheezing. The only other study to look at this young age group found, in contrast, that TNF-α-308A was not a risk factor for the development of asthma by 12 months of age in 373 Canadian infants from atopic families 27. The reason for this variance may relate to the fact that only 12 out of 281 (4%) of their infants were labelled as having “probable asthma”. Therefore, the numbers are rather small for significant differences to be found. In addition, Zhu et al. 27 mislabelled the G and A alleles in their paper. There is a marked increase in the proportion of TNF-α-308GA in the wheezy infants (with a corresponding deficit of GG). Although the current authors have no explanation for this finding, it is believed to be a genuine (biological) rather than genotyping error. Laboratory analysis of cases and controls were mixed together (blinded) and the present figures for controls match previously published work. Therefore, if there had been a systematic genotyping error, control data would also have been expected to be abnormal as well, which was not the case. However, these data do need to be repeated as the number of wheezy infants analysed was relatively small.
The current authors found that LT-α+252A was associated with asthma. This was found to be in keeping with previous studies which investigated the equivalent TNFB*2 20, 29, but again there are others studies which have found either the reverse 14, 15 or no association at all 18, 22, 23, 25, 28–31. Haplotype analysis showed one particular combination (TNF-α-308G, LT-α+252A/TNF-α-308A, LT-α+252A) led to a markedly increased risk for both asthma and infant wheezing, although with wide 99% CI. It is important to note that the admittedly unexpected high OR, and consequent wide CI, represents a conservative estimate as no control child actually had this gene combination, which would, therefore, give an OR of infinity. Therefore, two control children were allocated this haplotype, representing the upper 95% CI for 0 out of 148. Introducing an LT-α+252G allele had a counterbalancing effect, which was further enhanced by the introduction of a second TNF-α-308G allele, so that the TNF-α-308G, LT-α+252G/TNF-α-308G, LT-α+252A combination was actually protective for both asthma and infant wheezing.
Two grades of asthma severity were studied (mild/moderate versus severe), using a deliberately simple definition based on daily requirement of inhaled corticosteroids. The cut-off used was 800 mcg·day−1 of beclomethasone equivalent, as this is the one used by the European Respiratory Society Task Force on difficult or therapy-resistant asthma 38. Neither allele nor genotype frequencies were different between the two groups, although the TNF-α-308AA genotype did seem to be more common in the severe asthmatics (12%) compared with mild/moderate asthma or controls (5 and 6%, respectively). Chagani et al. 16 found that the TNF-α-308A polymorphism was not significantly more prevalent in 159 adults with fatal or near fatal asthma compared with 92 with mild or moderate asthma. Louis et al. 26 also found no association with asthma severity (as measured by lung function) in 193 adults. It is perhaps not surprising that the severity of asthma is not associated with these TNF gene polymorphisms given the multiple complex factors that determine the severity of disease in any individual 39. However, firm conclusions cannot be drawn from the current study as it was only adequately powered for “all asthma” and not the subgroups.
There are contrasting findings reported in some of the published asthma studies, and there are a number of potential reasons for the discrepancies. The nomenclature used for TNF gene polymorphisms is complicated, and mistakes in the literature further confuse the picture 27, 40, 41. Many problems arise from study design faults and some of the pitfalls of candidate gene association studies have been detailed in the excellent critique by Hall 42. Underpowered studies are a cause for concern as the lack of an association, or a false positive one, may simply be due to a type II or I error rather than a genuine finding. A study on replication validity of genetic association studies showed that those with <150 cases were less likely to be replicated subsequently 43, although many of the asthma studies reporting no association seem adequately powered, given the known allele frequency of the polymorphisms being tested. However, one of two studies to report an association between TNF-α-308G with asthma was a relatively small one (74 asthmatics, 50 controls), which might account for the unexpected finding 20. The present study was planned around the known allelic frequency of TNF-α-308G/A in the UK population, and the numbers reached that required for “all asthma”, although not for the severity subgroups; the number of wheezy infants analysed was also smaller than ideal. The controls used can be a problem and it is important to ensure the same racial mix is used. In the current study, the subjects and controls were from the same area of London (UK) and racial mix was similar in the groups. However, subgroup analysis was performed on the Caucasian subjects (whose ethnicity is more easily defined), and this confirmed the authors' findings from the whole study group on the effect of haplotypes. Many of the published asthma studies are from quite different population groups, which could account for some of their different findings, as the prevalence of polymorphisms within different racial groups is often markedly different 42. Comparisons of findings between different ethnic groups may be irrelevant due to population-specific masking of gene variant effects, or gene-environment interactions specific to one population 44. It is also important to ensure that the control population is appropriately chosen to exclude those with the disease being studied. This can be a problem with questionnaire-based studies with their known recall and bias problems. In the study by Winchester et al. 17, only 20 out of 649 (3%) pregnant UK or Irish females recalled having childhood asthma, which is certainly less than expected. The control school children in the present study were selected on the basis of lack of previous doctor-diagnosed asthma, which although not infallible is likely to be correct, especially in a group of 8–12-yr olds. Whilst it is possible some may have undiagnosed asthma, this would not have exaggerated any differences in the results (if anything it leads to underestimates of differences if the controls contained some asthmatics). It is certainly likely that the control population contained some children who had wheezed as infants, but who had outgrown this by 8-yrs of age and, hence, had no current diagnosis of asthma. The only way around this problem would be to conduct lengthy prospective studies. Furthermore, the age differences between the controls and wheezing infants means that any influence of environmental interactions on the asthma genotype could not be assessed due to the cross-sectional nature of this study.
Defining the patient population itself can be problematic, the current patient group had a diagnosis based on a single respiratory paediatrician's assessment and is assumed to be correct. A skewed study population was inevitable as the authors set out to enrol wheezy children, thus, the ratio of patients to controls (approximately equal) does not reflect the proportions found naturally in the UK population. This leads to falsely high PPV and NPV (although the sensitivity and specificity remain the same). Hence, Bayes' theorem was employed and factored in the incidence of wheezing in the general population to get a more realistic PPV. The definition of asthma in infancy is even more problematic, so a simple pragmatic approach of enlisting infants with an acute wheezing episode was adopted. The authors decided against attempting to categorise them as infantile asthma or viral-associated wheeze, and preferred not to employ the complicated definition of “probable asthma” as used by Zhu et al. 27 in their 12 month old infants. Undoubtedly some of the wheezy infant group would end up being classified as asthmatic when older.
Another problem with genotype patient studies is that of assessing whether any association found with a disease state is genuine or whether it is due to linkage disequilibrium. This is the phenomenon whereby combinations of alleles occur together more or less often than expected by chance alone, due to their positions relative to each other on the gene. Any association found with a particular polymorphism may then be due to a nearby polymorphism, which is in linkage disequilibrium with the one under scrutiny. There is extensive linkage disequilibrium on chromosome 6 44. The TNF-α gene is located within the class III region of the MHC close to the HLA-B locus. The MHC is the most polymorphic region of the genome and there is strong linkage disequilibrium within the MHC itself. The TNF-α-308A allele is strongly associated with the HLA A1, B8 and DR3 alleles 45. Furthermore, the DR3 and DR4 haplotypes produce higher levels of TNF-α while DR2 is associated with low production 4. However, secretion of TNF-α has been shown to be elevated in individuals carrying the TNF-α-308A allele regardless of their DR3 status, indicating that linkage to DR3 is not responsible for elevated TNF-α production 46. It has been suggested that the association of certain MHC haplotypes with TNF-α phenotypes may not be due to polymorphism within the TNF gene itself, but rather to variation of a further linked gene 47. In this study, it was not possible to assess HLA-DR status in the subjects, making it hard to comment on whether linkage to the MHC is relevant to the current findings. Although, previous findings showing that LD exists between the two loci studied (TNF-α-308 and LT-α+252) was confirmed. However, it was surprising to find disparity between the degrees of linkage disequilibrium in the disease groups tested. This could mean that some of the present results reflect the effects of another, closely related polymorphic locus. Mapping of the flanking areas of theses genes was not performed as part of this study, but could be incorporated into future studies.
One way of determining the relevance of linkage disequilibrium is to study the associations with extended haplotypes 42. In the study by Moffatt and Cookson 14 of 413 Australian subjects from 88 nuclear families, questionnaire-diagnosed asthma was positively associated with the TNF-α-308A allele and also with the LT-α B*1 allele. Although they found the TNF-α-308A, LT-α B*1 and HLA-DRB1*03 alleles are in linkage disequilibrium, by studying haplotypes they showed the association with asthma was not due to the HLA-DR locus. In a larger study of >1,000 Australian subjects, Moffatt et al. 15 then showed the extended haplotype TNF-α-308A/LT-α B*1/HLA-DRB1*03 was weakly associated with questionnaire-diagnosed asthma, whilst the rarer TNF-α-308A/LT-α B*1/HLA-DRB1*02 was more strongly associated with asthma and bronchial hyperreactivity. Although the latter haplotype carried a higher risk of asthma, it was rare, so had less impact on the population-attributable risk of asthma. It was difficult to differentiate between the effects of these alleles in this haplotype, but a functional polymorphism contributing to asthma susceptibility is likely to be contained within this haplotype. In a study of 239 asthmatics, an association of asthma with TNF-α-308A/HLA-DRB1*03 was found only in those with positive skin prick testing to Lolium (ryegrass), whilst an association with TNF-α-308A/HLA-DRB1*1104 was found only in those sensitive to Parietaria (herb pollen) 48. Di Somma et al. 48 suggested this implied that the association of TNF-α-308A with asthma reflected linkage disequilibrium with genes influencing a specific immune response. Whilst studying haplotype and genotype combinations may give more useful results, if too many polymorphisms are included the subgroups become so numerous that huge patient numbers are required, otherwise statistical evaluation becomes difficult. In the study by Moffatt et al. 15 that combined three loci (TNF-α-308, LT-α NcoI and HLA-DRB1), there were 56 possible haplotypes, therefore even with over 1,000 subjects many of the haplotype frequencies were too small to be meaningful.
If a direct functional effect of a polymorphism can be demonstrated, it makes the clinical significance of the polymorphism greater. It also makes it more likely that the polymorphism itself is truly associated with the disease being studied, rather than the association being entirely due to linkage disequilibrium with the true aetiological gene 47. For this reason nasal TNF-α levels were measured in the wheezy infants, as a marker of the functional phenotype, and related to their genotypes. Although the data did not quite reach statistical significance, due to the relatively low numbers, higher nasal TNF-α levels were found to be produced by those infants with one or two LT-α+252A alleles, during an acute wheezing episode. This is consistent with the finding that the LT-α+252A allele was more common in the wheezing children compared with controls and confirms the LT-α+252 locus has some functional significance. It is also consistent with a study on severe sepsis, in which those patients homozygous for TNFB2 (equivalent of LT-α+252A) had higher plasma levels of TNF-α 49. By contrast, when the nasal TNF-α levels were related to the TNF-α-308 locus, the reverse of that expected was found. Significantly lower nasal TNF-α levels were found in the presence of one or two TNF-α-308A alleles, yet this allele was significantly more common in the wheezing children. In vitro work has also shown this allele to be associated with greater TNF-α production 7, 8, 10. There is no obvious explanation for this unexpected finding. Nasal TNF-α levels were also influenced by the TNF-α-308/LT-α+252 genotype combinations. However a recent review by Bayley et al. 44, suggested that the TNF-α-308GA polymorphism is nonfunctional and has no influence on the activity of the TNF promoter.
Studying gene polymorphisms may aid understanding of the pathophysiology of childhood wheezing, but an overriding question is whether there will be any practical or clinical use for it, and particularly whether it will be useful for an individual child. Most infant wheezers grow out of their condition in early childhood but some will end up diagnosed with genuine asthma. It has been suggested that early intervention with inhaled corticosteroids may prevent the development of asthma 50, but clearly it would be pointless to treat all wheezy infants as in most cases the condition will resolve spontaneously 51. Therefore, an identifiable marker that would indicate whether an infant is likely to develop genuine asthma would be most useful. Perhaps this is where the study of TNF-α and other gene polymorphisms might have a role. The present study has shown that a haplotype containing TNF-α-308A and LT-α+252A alleles gave the greatest risk for developing asthma, so should all wheezy infants with this genotype receive long-term inhaled corticosteroids? Although specificity was high, the sensitivity was rather low. This means if a child has the relevant genotype, it is quite likely they will develop asthma, but since the particular haplotype is relatively rare, most children with diagnosed asthma will have an alternative genotype. Nevertheless in the presence of the genotype, there might be a case for more aggressive early treatment with corticosteroids. Time and further studies might shed light on this area. Known cohorts could be studied, such as that in Tuscon, in which it is now known which children were “transient wheezers” and which “persistent wheezers” at 6-yrs of age 52. If such data confirmed the current findings, then the next stage would be a prospective early intervention study on those with the at risk genotype. It would need large numbers to identify sufficient patients with the relevant genotype, and of course there are so many other risk factors that must be taken in to account (for example atopy, passive smoking, pre-existing lung function), that matching groups would be extremely difficult.
In conclusion, it was found that polymorphisms that affected tumour necrosis factor-α production were associated with childhood wheezing, but did not seem related to the severity of asthma. In wheezy infants, nasal tumour necrosis factor-α levels were influenced by the genotype, but not always in the expected direction. Identification of certain high-risk haplotypes may be important for future early intervention studies.
Acknowledgments
The authors would like to thank the children and their parents who participated in this study. They would also like to thank J. Mak from National Heart Lung Institute (London, UK) who helped with the original DNA extraction and M. Howell from Southampton General Hospital (Southampton, UK) for helping I. M. Balfour-Lynn to understand the nomenclature. Finally, they would like to thank Chelsea and Westminster Hospital Special Trustees (London, UK) who funded this study.
- Received June 15, 2004.
- Accepted June 8, 2005.
- © ERS Journals Ltd
References









