





Abstract
Endogenous inhibitors tightly control the activity of proteinases in the extracellular space. Proteinase/antiproteinase imbalance may be caused by predominance of proteinases, resulting in severe tissue damage or abundance of proteinase inhibitors, leading to a shift in the balance of synthesis and degradation of extracellular matrix proteins and accumulation of these matrix components. Lung fibrosis is characterised by accumulation of fibrous matrix proteins in the alveolar interstitium.
The activity of cathepsin D and amounts of cathepsins D and B in bleomycin-injured rat lung tissue and alveolar macrophages were examined. In addition, the activities of cathepsins and cysteine proteinase inhibitors (CPIs) in bronchoalveolar lavage fluid (BALF) were determined.
No cathepsin but high CPI activity and large amounts of procathepsin B were detected in the BALF. In the alveolar lumen, the disturbed proteinase/antiproteinase balance for cysteine proteinases was clearly dominated by CPIs. In alveolar macrophages, the main source of increased cathepsin levels, large changes in cathepsin B and D content were observed during the inflammatory phase, corresponding to the occurrence of procathepsin B in BALF. With the end of the phase of tissue remodelling, imbalances in cathepsin and CPI activities were largely eliminated. Immunoblot data, revealing an increase in cathepsin D levels in myofibroblast-like cells compared to fibroblasts and in resting fibroblasts compared to proliferating cells, implicate this proteinase in the differentiation and conversion processes occurring at the beginning of the fibrotic phase of lung injury.
The results show that cathepsin amounts and activities are increased transiently in lung tissue during regeneration processes in bleomycin-induced lung injury. Imbalances of cathepsin and cysteine proteinase inhibitors activities are also a phenomenon of the phase of tissue remodelling initiated by lung injury.
This study was supported by the Ministry of Education and Research, Bonn, Germany (FKZ 01ZZ5904).
The antineoplastic antibiotic bleomycin exhibits pulmonary fibrosis as its major side-effect. Intratracheal administration of bleomycin into the lung of rats causes a sequence of morphological and biochemical changes, such as acute inflammation with oedema and damage to the alveolar epithelium aswell as influx of inflammatory cells. Finally, a phase of chronic injury, which is characterised by tissue remodelling with proliferation of fibroblasts and pneumocytes and thickening of the alveolar walls, follows 1–3. This fibrotic process is accompanied by excessive production and accumulation of connective tissue components. In rats and other rodents, bleomycin-induced lung injury remains restricted to a limited number of foci and subpleural areas, and, after tissue regeneration, the animals survive 4.
In inflamed tissue, extensive protein breakdown occurs 5–8. The activity of proteinases is tightly controlled and proteinase/antiproteinase imbalance may cause severe tissue damage 9–11. Previously, remarkable imbalances in metalloelastase and elastase inhibitory activity were reported in ratlung tissue following intratracheal bleomycin administration 12. These data on sustained high elastase activity infibrotic rat lung led to the conclusion that accumulation ofextracellular matrix proteins does not necessarily require decreased or inhibited proteinase activities. A disturbed balance might include increased proteolytic degradation with concurrent prevalence of matrix production. In the same study, it was shown that the activities of the cysteine proteinases cathepsins, B, H, L and S, in lung tissue homogenates and alveolar macrophages were elevated during the tissue regeneration phase 12.
Lysosomal enzymes recruited from the cysteine and aspartic proteinase families play an important role in degradation processes 5–7, 13. The cysteine proteinases cathepsins B, L and S are known to contribute to the degradation of several types of collagen and elastin in the extracellular space 5, 7. Lysosomal proteinases intracellularly complete the extracellular cleavage of collagens, elastin and proteoglycan core proteins by serine and metalloproteinases 7. They are partially secreted into the extracellular space and the bronchoalveolar lavage fluid (BALF) 6, 7, 14, 15. The activities of cysteine proteinases are controlled by their endogenous inhibitors, the cystatins 6, 7, 16.
The aim of the present study was to determine the catalytic activities of cathepsins and cysteine proteinase inhibitors (CPIs) (total residual activity of CPIs) in BALF and identify the cellular origin of the elevated cathepsin activities in bleomycin-injured rat lung tissue immunohistochemically and by comparison with immunoblot data from isolated lung fibroblasts. For studies on specific enzyme amounts, the study focused on cathepsins B and D, which account for the major portion of intralysosomal proteolytic activity and have been shown to be upregulated in the lung tissue of bleomycin-treated mice 15, 17, 18.
Materials and methods
Induction of pulmonary fibrosis
For induction of pulmonary fibrosis, male Wistar rats (n=58; two to four animals per time point) weighing 350–400 g received a single dose of 7.5 U·kg body weight−1 bleomycin sulphate (Mack, Illertissen, Germany) dissolved in 0.25 mL 0.9% NaCl solution by endotracheal instillation. Controls were given saline alone. The animals were sacrificed at the given time points and their lungs flushed free of blood across the right ventricle using phosphate-buffered saline (PBS) (pH 7.4) containing 50 U·mL−1 heparin.
Determination of collagen content
The collagen content of lung tissue was determined according to Walsh et al. 19. Pieces of lung tissue were homogenised in 0.5 M acetic acid and collagen was extracted for 12 h at 4°C. After centrifugation for 30 min at 10,000×g, 50 µL supernatant were dispensed into a microtitre plate. The samples were incubated for 16 h at 37°C (humidified) and dried for 24 h at 37°C. The plates were washed three times with 200 µL·well−1 distilled water and stained with 100 µL 1% Sirius Red F3BA (BDH Laboratory Supplies, Poole, UK) in saturated picric acid for 1 h at 20°C. After washing the wells five times with 200 µL 10 mM HCl, bound dye was eluted with 200 µL 0.1 M NaOH by shaking for 5 min. Absorbance at 540 nm was measured in a microplate reader. Rat tail collagen was used as a standard at concentrations ranging 0.01–0.4 mg·mL−1.
Bronchoalveolar lavage
BALF and alveolar macrophages were obtained by lavage of the lungs across the trachea with 10 PBS washes (each 5 mL). After centrifugation for 10 min at 500×g, the cells were washed twice with PBS and seeded into culture dishes (1×105 cells·cm−2). BALF proteins in the supernatant were concentrated by ammonium sulphate precipitation (final concentration 4.9 M) followed by centrifugation for 30 min at 20,000×g at 4°C and dialysis against 67 mM phosphate buffer (pH 7.4). These samples served for determination of protein content and cathepsin and CPI activity as well as immunoblotting. Cells were cultured in Dulbecco modified Eagle medium supplemented with 10% foetal calf serum (FCS), 50 International Unit (IU)·mL−1 penicillin and 50 µg·mL−1 streptomycin for 3 h. Nonadherent cells were then removed and the medium was changed to DIF 1000 (Biochrom Seromed, Berlin, Germany) for 12 h. Cells were harvested using a cell strainer. The isolated cells comprised ≥98% alveolar macrophages (by microscopy of cytospin preparations). Viability was ≥95% as checked by trypan blue exclusion.
Western blotting
For detection of the type I pneumocyte-specific protein rat type I cell marker (RTI40) in the BALF, proteins were sedimented by centrifugation for 2 h at 100,000×g at 4°C, as described previously 2. Proteins were redissolved in 360 mM tris-(hydroxymethyl)-aminomethane (Tris)/HCl (pH 6.8) containing 190 mM sodium dodecyl sulphate (SDS), 0.03% bromophenol blue and 55% glycerol (volume/volume) and incubated for 5 min at 95°C. Aliquots of 10 µL were subjected to SDS‐polyacrylamide gel electrophoresis under reducing conditions. For detection of cathepsin forms in BALF, samples of concentrated BALF were used. Proteins were separated by SDS‐polyacrylamide gel electrophoresis (5–15% gradient gels) under reducing conditions. The separated proteins were blotted on to nitrocellulose and, after blocking in 20 mM Tris/HCl (pH 7.6) containing 140 mM NaCl and 5% nonfat milk powder (BioRad, Hercules, CA, USA) for 2 h at 25°C, specific proteins were labelled using the following primary antibodies: monoclonal mouse anti‐RTI40 (kind gift of L.G. Dobbs, Cardiovascular Research Centre, University of San Francisco, CA, USA; 1:500 dilution); polyclonal rabbit antirat cathepsin B (Upstate, Lake Placid, NY, USA; 1:200 dilution); and polyclonal rabbit antihuman cathepsin D(Biogenex, San Ramon, CA, USA; 1:3,000 dilution, cross-reacts with rat cathepsin D). Horseradish peroxidase-conjugated goat antimouse immunoglobulin (Ig) (New England Biolabs, Frankfurt, Germany) was used as a secondary antibody. The blots were developed with luminol reagent and hydroperoxide (ECL; New England Biolabs) andexposed to Hyperfilm (ECL; Amersham, Freiburg, Germany). Bands were analysed densitometrically.
Preparation and culture of lung fibroblasts
Fibroblasts from the lungs of normal Wistar rats were prepared as described previously 20, by enzymic dissociation of lung tissue using 0.2% collagenase type I (Sigma, Deisenhofen, Germany), 0.15% pronase (Boehringer, Mannheim, Germany), 0.08% dispase (Boehringer) and 0.9% deoxyribonuclease (Sigma). After filtration (pore size 100 µm), cells were collected by centrifugation and seeded into culture dishes in Roswell Park Memorial Institute (RPMI) 1640 culture medium supplemented with 10% FCS (Biochrom Seromed), 50 IU·mL−1 penicillin and 50 µg·mL−1 streptomycin. The cells were passaged routinely and the media changed every 3 days.
Fibroblasts were cultured on either plastic tissue culture dishes for 5 days for initiation of transdifferentiation to myofibroblast-like cells or a matrix composed of 20% Engelbreth-Holm-Swarm (EHS) matrix (Becton Dickinson, Heidelberg, Germany) and 80% rat tail collagen for maintenance of the fibroblast-like cell status 21. For comparison of proliferating and resting fibroblasts, cells were cultured in tissue culture dishes in RPMI medium supplemented with either 10% FCS or 0.6% heat-inactivated FCS for 3 days. Cells were harvested, homogenised by sonication and stored at −80°C until used for immunoblotting. Cell culture experiments were repeated three times.
Immunohistochemistry
For immunohistochemical studies, the lower part of theright lobe of the lung was taken for fixation in 4% neutralised formaldehyde solution and routinely processed for embedding in paraffin. For the indirect immunoperoxidase technique, dewaxed paraffin sections were microwaved as described previously 22 and incubated with primary polyclonal rabbit antisera for 1 h at 37°C. The following antisera were used: rabbit antihuman cathepsin D (Dianova, Hamburg, Germany; 1:20 dilution); rabbit antirat cathepsin B (provided by H. Kirschke, University of Turku, Finland; 1:800 dilution); and rabbit antirat cathepsin H (H. Kirschke; 1:1,000 dilution). As negative controls, the primary antisera were replaced with either PBS or irrelevant rabbit IgGs. Goat antirabbit IgG coupled to peroxidase was used as the secondary antibody (1:400 dilution). The sections were stained with diaminobenzidine for 8 min and counterstained with haematoxylin. All immunostaining was performed in triplicate employing two different tissue samples of each lung.
Double-label immunofluorescence staining was performed employing polyclonal rabbit antisera directed against cathepsin B (1:800 dilution) or D (1:20 dilution), or monoclonal mouse antibodies directed against cytokeratins (MNF 116; Dako, Hamburg, Germany; 1:150 dilution) or smooth muscle actin (Dianova; 1:100 dilution). The corresponding secondary antibodies (1:80 dilution) were coupled to fluorescein isothiocyanate or Texas red (Dianova). Details of the double-staining method are described in 23.
For demonstration of collagen deposition, sections were evaluated using Goldner's trichrome stain.
Activity of cathepsins and cysteine proteinase inhibitors
For determination of the catalytic activities of cathepsin D, lungs were trimmed of heart, extrapulmonary airways and blood vessels. Lung tissue and isolated macrophages were homogenised in a Potter homogeniser and by sonication followed by centrifugation for 1 h at 20,000×g at 4°C. Protein concentrations were determined according to Bradford 24 using bovine serum albumin as standard.
Cathepsin D activities were determined in lung tissue, macrophages and concentrated BALF as described by Orlowski et al. 17 using H‐d‐Phe-Ser(Benzyl)-Phe-Phe-Ala-Ala-p-aminobenzoate (Calbiochem, Bad Soden, Germany) as substrate and 0.1 mM pepstatin (ICN Biomedicals, Eschwege, Germany) as inhibitor. One unit represents the activity liberating 1 micromole of p‐aminobenzoate per minute.
The activities of cathepsins B, L, S and H were measured asdescribed previously with some modifications 12. Briefly, samples of concentrated BALF were incubated with N‐α‐benzyloxycarbonyl-Phe-Arg‐7‐amido‐4‐methylcoumarin (cathepsins B, L and S) and H‐Arg‐7‐amido‐4‐methylcoumarin (cathepsin H) as peptide substrates according to Barrett andKirschke 25. For differentiation of the activities of cathepsins B, L and S, the following inhibitors were used: cathepsin B: (l‐trans-(propylcarbamoyl)oxirane‐2‐carbonyl)‐l‐isoleucyl‐l‐proline (CA‐074; Calbiochem, Schwalbach, Germany); cathepsin L: N‐α‐benzyloxycarbonyl-Phe-Tyr(t-Bu)CHN2; and cathepsin S: N‐α‐benzyloxycarbonyl-Val-Val-NleCHN2. All other peptide substrates and inhibitors were from Bachem (Heidelberg, Germany). CPI activities were determined in lung tissue, macrophages and concentrated BALF according to Thiele et al. 26 with some modifications. Briefly, aliquots of protein solution were added to papain (final activity 5 µU) in a total volume of 100 µL assay buffer (0.3 M sodium acetate (pH 5.5), 2 mM ethylenediamine tetraacetic acid, 0.15% Brij 35, 1 mM dithiothreitol) and incubated for 30 min at 30°C. After dilution with 400 µL 0.1% Brij 35, residual papain activity was determined using N‐α‐benzyloxycarbonyl-Phe-Arg‐7‐amido‐4‐methylcoumarin as substrate and a Shimadzu RF 5000 fluorimeter (Shimadzu Europa, Duisburg, Germany). CPI activity was calculated as milliunits of papain activity inhibited per milligram of protein.
Statistical analysis
All experiments were performed to investigate the effect of time. Therefore, one-way analyses of variance (ANOVA) were performed with time as the factor. They were completed by a priori comparisons of means building simple contrasts to test the time effect in relation to the basic mean.
For the experiments with a control group, univariate ANOVA was conducted. The impact of a different increase in time between cases and controls was tested by an interaction term included in the model. For comparison of means, contrasts or test procedures for multiple comparisons, such as the Newman-Keuls approach, were also used. If the assumptions underlying the analyses were violated, the robust Brown-Forsythe statistic was employed instead of the F statistic, and, for pairwise post-hoc comparisons, the test of Tamhane was used. The significance level was set at α=0.05, and, if multiple comparisons were performed, this value was adjusted according to Šidak.
Results
Assessment of lung injury and fibrosis
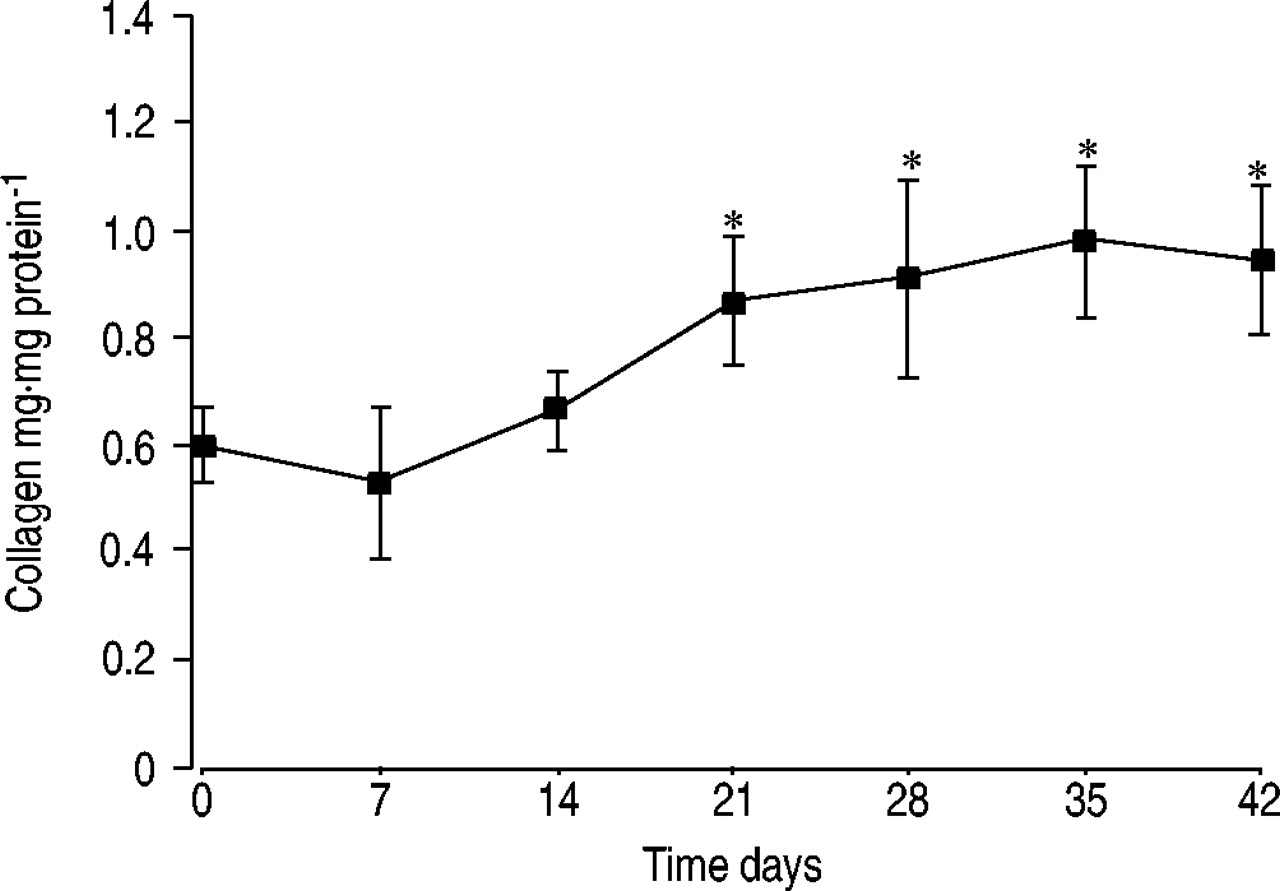
The progress of injury of the alveolar epithelium induced by intratracheal administration of bleomycin was monitored by detection of the soluble form of the type I pneumocyte-specific protein RTI40 in BALF by Western blot analysis (table 1⇓). The concentration of RTI40 was found to be increased by ∼80‐fold at day 7 and remained elevated for ∼6 weeks. The total protein concentration in the BALF increased to a maximum of ∼40 times that of the controls at day 7 followed by a decrease to approximately twice that of the controls at day 42. The ratio of RTI40 (in arbitrary units) to total protein (in milligrams per litre) reached maximum values of ∼40 at days 14 and 28 (table 1⇓). The development of pulmonary fibrosis was assessed by microscopic analysis of tissue sections stained using Goldner's trichrome stain (data not shown) and determination of the collagen content of lung tissue (fig. 1⇓). The collagen content increased between days 14 and 21 and remained elevated for ≥42 days after bleomycin treatment.

Collagen content of lung tissue following endotracheal bleomycin administration to rats. Data are presented as mean±sd (n=4 per time point). *: p<0.05 versus 0 days.
Concentration of total protein and the type I pneumocyte-specific protein RTI40 in bronchoalveolar lavage fluid from rats after bleomycin administration
Cathepsins in normal and fibrotic lung
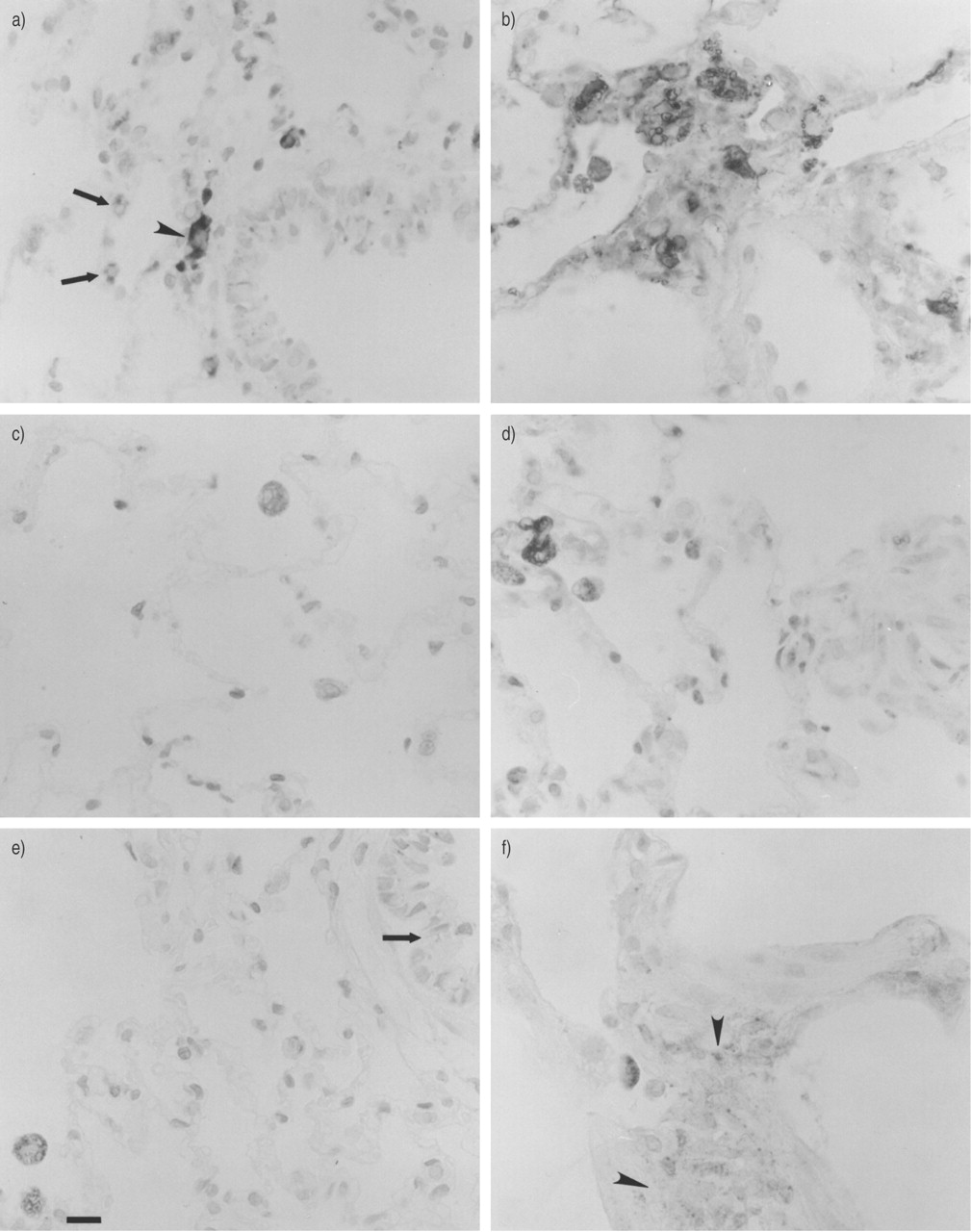
Cathepsin H was present in alveolar macrophages from normal rat lung; however, only a few macrophages were positive for cathepsins B and D (table 2⇓ and fig. 2a, c and e⇓). Type II pneumocytes were positive for cathepsin H (fig. 2a⇓). A few bronchiolar epithelial cells contained cathepsin B‐, H‐ and D‐positive granules (table 2⇓ and fig. 2a and e⇓).

Cathepsins in a, c, e) normal and b, d, f) fibrotic (day 28) rat lung. Immunoperoxidase staining of cathepsins H (a, b), B (c, d) and D (e, f) in paraffin sections. Arrows indicate a) cathepsin H‐positive type II pneumocytes (note the strong staining of an alveolar macrophage (arrowhead)) and e) weak staining of granules in the bronchial epithelium. Arrowheads indicate interstitial immunoreactivity for cathepsin D (f). Scale bar represents 20 µm.
Distribution of cysteine and aspartic proteinases in normal and injured (fibrotic) rat lung
In the lungs of bleomycin-treated rats (day 28), the immunoreactivity of alveolar macrophages for all cathepsins was increased in both intensity and number of positive cells, for cathepsin B in type II pneumocytes and for cathepsins B, H, and D in some interstitial cells of fibrotic foci (table 2⇑ and figs 2b, d and f⇑). Immunoreactivity for cathepsin D was observed in fibrotic foci in the extracellular space (table 2⇑). Double-label immunofluorescence staining employing the epithelial marker cytokeratin confirmed the presence of cathepsin B in the alveolar epithelium (fig. 3a and b⇓). Double-label immunofluorescence staining with antibodies directed against smooth muscle α‐actin and cathepsin D revealed the presence of this proteinase in myofibroblasts (fig. 3c and d⇓).

Colocalisation of a) cytokeratin and c) cathepsin B, as well as b) smooth muscle actin and d) cathepsin D in fibrotic rat lung (day 28) by double-label immunofluorescence staining. Arrows indicate colocalisation of cathepsin B and cytokeratin in alveolar epithelial cells and arrowheads indicate cytokeratin-negative but cathepsin B‐positive alveolar macrophages (a, c). Some myofibroblasts (arrowhead) were positive for cathepsin D (b). Cathepsin D is partially localised extracellularly in the interstitium (d). Scale bar represents 20 µm.
Cathepsin D activity and immunoblotting data for cathepsins D and B
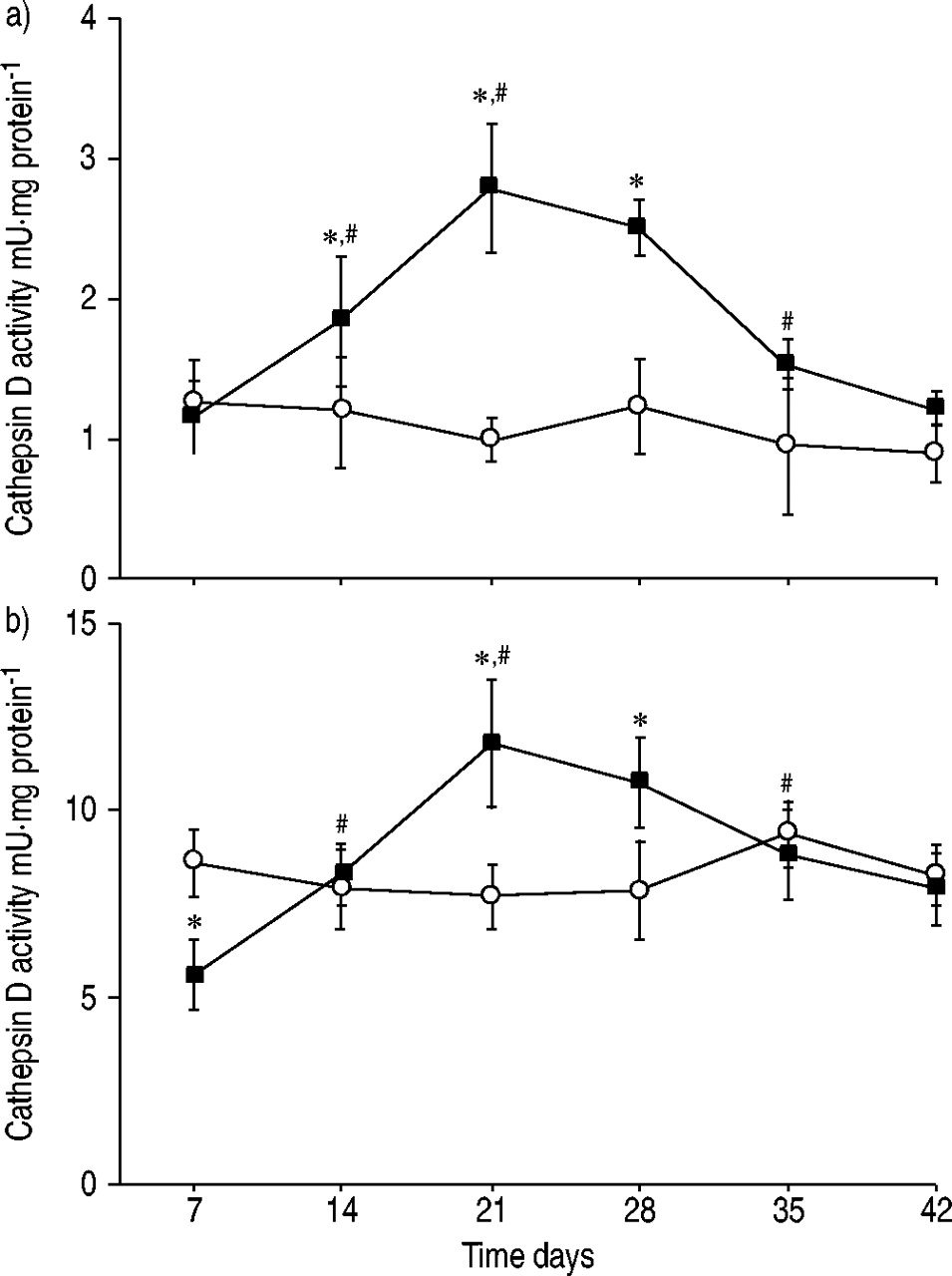
In lung tissue, the specific activity of cathepsin D increased after bleomycin administration and reached maximum levels of ∼2.5 times those of controls at day 21 (fig. 4a⇓). At day 42, the activity was at control level. In alveolar macrophages, cathepsin D activity was found to be decreased at day 7 after bleomycin administration (fig. 4b⇓). Thereafter, the activity increased and reached a maximum of ∼1.5 times that of controls at day 21. The activity reached control levels at day 42. No cathepsin D activity was found in the BALF.
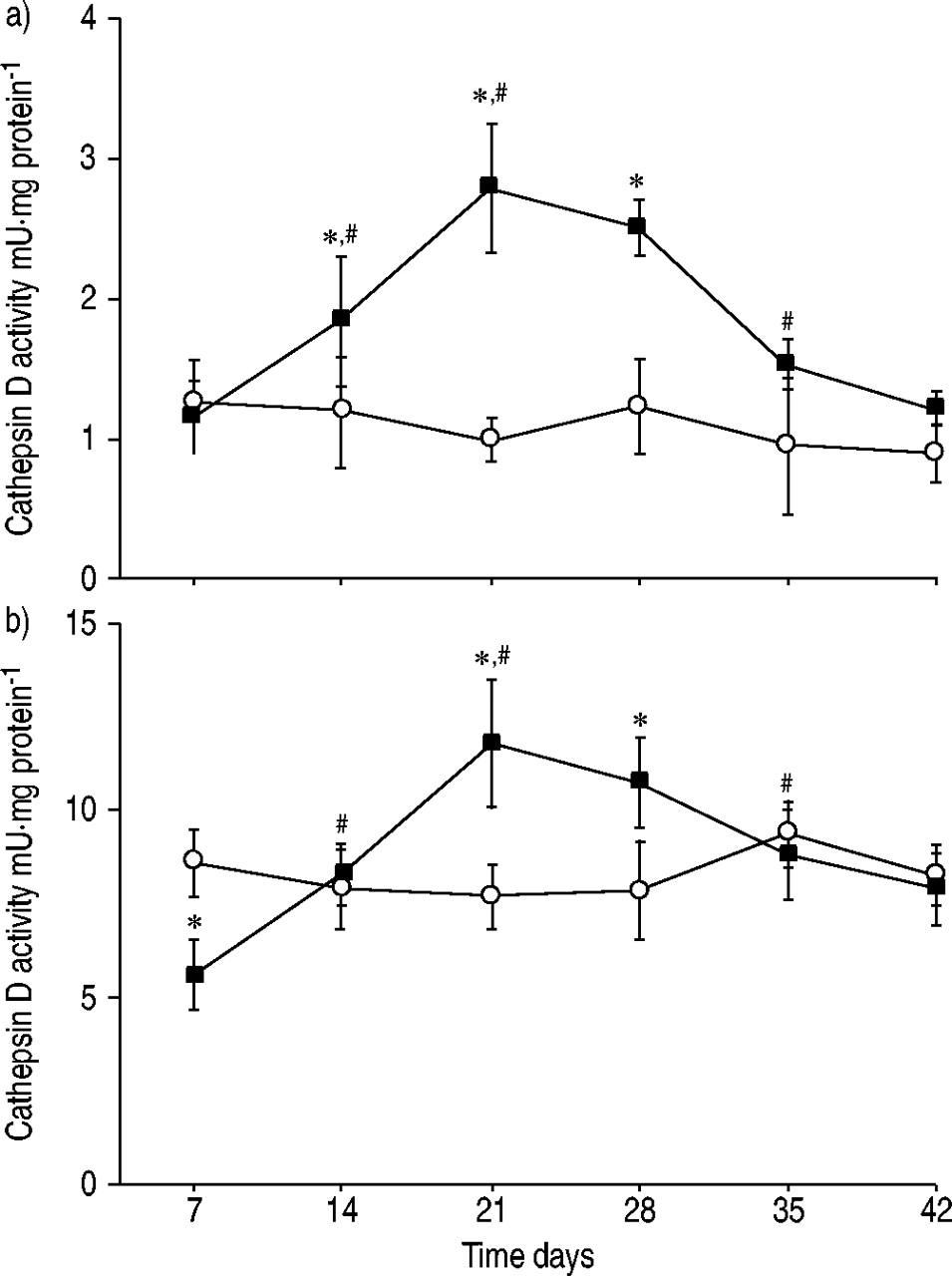
Specific activity of cathepsin D in a) lung tissue and b) isolated alveolar macrophages from rats following bleomycin administration (▪: bleomycin (n=4 per time point); ○: control (n=2 per time point)). Data are presented as mean±sd. *: p<0.05 versus control; #: p<0.05 versus preceding time point.
The time course of changes in cathepsin D levels in lung tissue is shown in figure 5a and b⇓ for the single- and heavy-chain species of the enzyme. The predominant form of cathepsin D in lung tissue is the single-chain species of ∼43 kDa. During maturation, this form is cleaved to give the double-chain form consisting of heavy-chain cathepsin D (33 kDa) and a light-chain species. The amount of single-chain species increased after bleomycin administration, reaching a maximum of approximately twice that of controls at day 35. The 33‐kDa cathepsin D form increased to a maximum of approximately five times that of controls at day 21. At day 42, the amount of this enzyme species was at control levels. In alveolar macrophages, the predominance of single-chain cathepsin D was more pronounced compared to lung tissue (fig. 5c and d⇓). The amount of this species was found to be reduced to ∼50% of control levels at day 7 after bleomycin administration. Thereafter, it increased, reaching control levels at day 21. The time course of changes in the heavy-chain species showed decreased amounts at day 7 and thereafter a broad maximum of ∼1.5 times that of controls at days 21–35. In the BALF, only traces of 43‐kDa cathepsin D were found (data not shown).
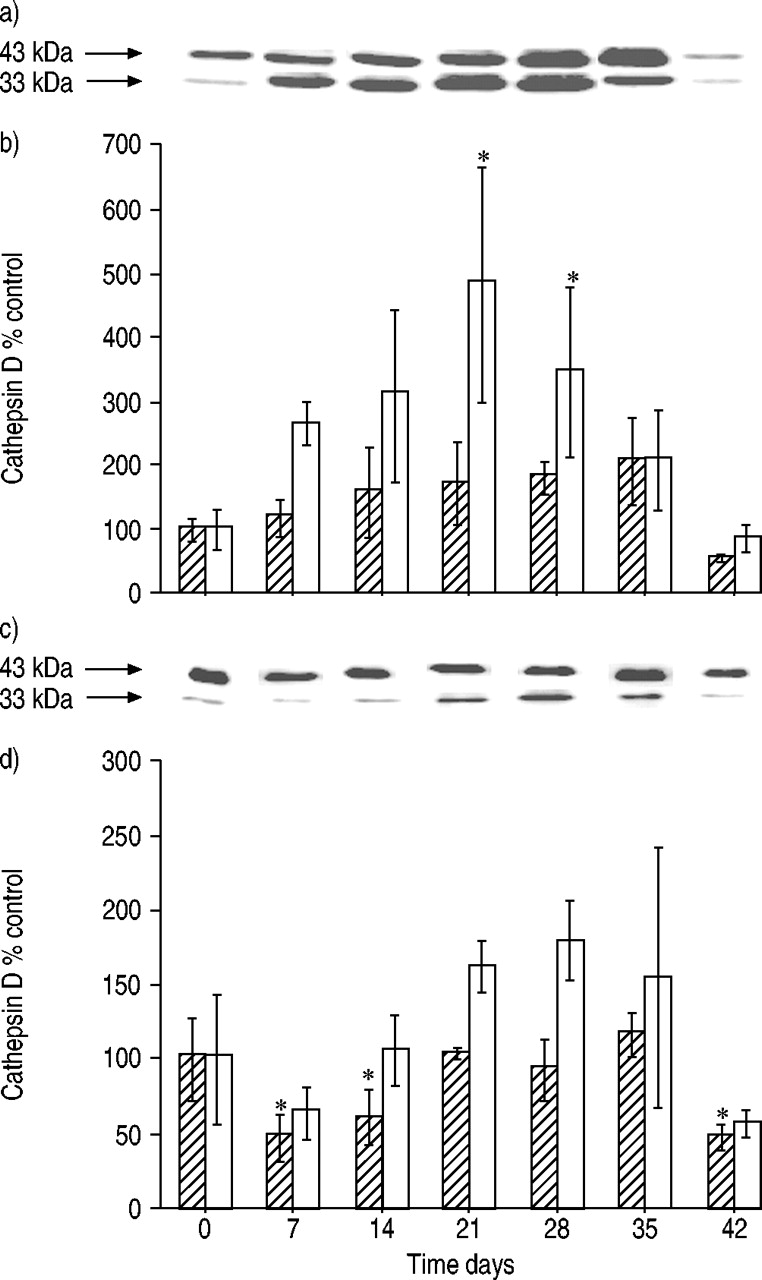
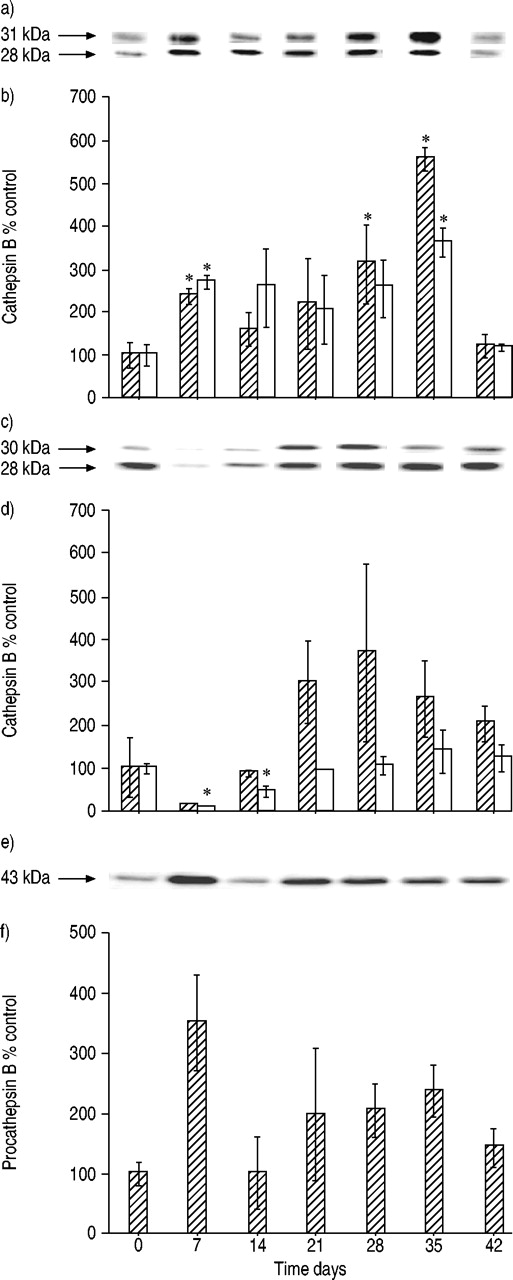
Amounts of single-chain (└ 43 kDa) and heavy-chain (□; 33 kDa) cathepsin D in a, b) lung tissue and c, d) alveolar macrophages isolated from the lungs of rats after bleomycin administration. a, c) representative immunoblots and b, d) densitometric evaluation of blot data. Data are presented as mean±sd (n=3 per time point). *: p<0.05 versus 0 days.
Cathepsin B occurs as a single-chain species of 31 kDa, resulting from cleavage of the 43‐kDa procathepsin B and matures to form a heavy-chain species of 28 kDa. Single- and heavy-chain forms were detected in lung tissue and alveolar macrophages (fig. 6a–d⇓). In normal lung tissue and isolated alveolar macrophages, heavy-chain cathepsin B was found in greater amounts than the single-chain species. In injured lung tissue, both forms of cathepsin B increased, reaching maximum levels of approximately three to five times that of controls at day 35 (fig. 6a and b⇓). At day 42, the amounts were at control levels. In alveolar macrophages, there was an initial marked decrease in cathepsin B single- and heavy-chain form levels followed by an increase to a maximum at day 28 (fig. 6c and d⇓).

Amounts of single-chain (└ 31 kDa) and heavy-chain (□; 28 kDa) cathepsin B in lung tissue (a, b), alveolar macrophages (c, d) and procathepsin B (e, f) (43 kDa) in bronchoalveolar lavage fluid after intratracheal bleomycin administration. a, c, e) representative immunoblots for each cathepsin form, and b, d, f) densitometric evaluation of blot data. Data are presented as mean±sd (n=3 per time point). *: p<0.05 versus 0days.
Immunoblotting for specific cathepsin B forms in the bronchoalveolar lavage fluid revealed the occurrence of the 43‐kDa procathepsin B, which was found to be increased by ∼3.5 times at day 7 after endotracheal bleomycin administration (fig. 6e and f⇑). No other cathepsin B forms were found.
Cathepsin D in fibroblasts and myofibroblast-like cells
In order to better assess the processes leading to an increase in cathepsin levels in injured lung tissue, isolated lung fibroblasts were cultured under conditions promoting transdifferentiation to myofibroblast-like cells (culture on plastic surfaces). For comparison, fibroblasts were cultured on an EHS/collagen matrix supporting the maintenance of a fibroblast-like differentiation status. Figure 7a⇓ shows the result of representative immunoblotting of homogenates of fibroblasts cultured under these various conditions. Both types of cells contained the 43‐kDa single chain of cathepsin D. Transdifferentiation of lung fibroblasts to myofibroblast-like cells resulted in a marked increase in the amount of cathepsin D.

Amounts of cathepsin D in isolated lung fibroblasts cultured under different conditions. a) Immunoblot for cathepsin D using cells cultured for 5 days on a matrix composed of 20% Ehgelbreth-Holm-Swarm matrix 80% collagen (1) or in plastic tissue culture dishes (2) (representative blot of three experiments). b) Comparison of the cathepsin D content of proliferating (1) and resting (2) lung fibroblasts, cultured in the presence of 10% foetal calf serum (FCS) or 0.6% heat-inactivated FCS, respectively (representative blot of three experiments).
The cathepsin D content of proliferating and resting fibroblasts was compared. Figure 7b⇑ demonstrates that transition of cells to a quiescent state led to an increase in cathepsin D levels.
Activities of cysteine proteinases and cysteine proteinase inhibitors
In samples of concentrated BALF, no catalytic activity was detected for cathepsins B, L, S and H. The specific activity of CPIs in BALF was found to be increased by approximately seven-fold at day 7 after bleomycin administration. Thereafter, it decreased rapidly and reached control levels at day 28 (fig. 8⇓). In homogenates of lung tissue and alveolar macrophages, no CPI activity was observed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cysteine proteinase inhibitor (CPI) activity in bronchoalveolar lavage fluid from rats following endotracheal bleomycin administration (└: bleomycin (n=4 per time point); □: control (n=2 per time point)). Data are presented as mean±sd. *: p<0.05 versus 0 days.
Discussion
When the present authors' studies on proteinases and proteinase inhibitors in injured rat lung started, the initial hypothesis implied the following: 1) cathepsins may participate not only in intracellular but also in extracellular degradation of proteins of the extracellular matrix; and 2) imbalances in the activities of cathepsins and endogenous cathepsin inhibitors may contribute to unwanted protein degradation and onset of lung injury 12. The principal findings of the present study demonstrate that, during the inflammatory phase of bleomycin-induced lung injury, CPI activity in BALF predominates regardless of the high amounts of procathepsin B that occur as the result of mistargeting or regulated secretion 7. The absence of processed active cathepsin B and of any cathepsin activity in BALF suggests that proteins in the alveolar lumen are protected against proteinase attack.
Two phases to the changes in cathepsin and CPI levels were identified. Approximately 7 days after lung injury, a marked increase in procathepsin B and CPI levels in BALF was detected, whereas the levels of cathepsin B and D forms and cathepsin D activity in alveolar macrophages were found to be decreased. During this period, markedly elevated levels of RTI40 and increased protein concentrations in BALF point to epithelial damage and leakage of blood plasma into the alveolar space. This phase was complete ∼14 days after bleomycin administration. This is in accordance with data reported earlier showing that the inflammatory phase of lung injury is complete ∼10 days after bleomycin administration 1, 12. During the following 2 weeks, characterised by the onset of collagen accumulation, levels of cathepsin B and D species as well as cathepsin D activity were elevated in lung tissue and alveolar macrophages. No further increases in collagen levels in lung tissue were observed 28 days after injury. The typical focal form of moderate fibrosis induced by bleomycin had developed 1, 4. At the time at which cathepsin levels in lung tissue and alveolar macrophages had reached maximum levels or started to decline, deposits of cathespsin D were detected in the extracellular matrix and in myofibroblasts in fibrotic areas. Cathepsin B levels in epithelial cells were found to be increased. Macrophages represent the predominant cell type contributing to elevated cathepsin levels, confirming earlier data 27, 28, but other cell types also participate.
In general, the cathepsin D activity in lung tissue and alveolar macrophages showed a similar time course to that reported in a previous study on cathepsins B, H, L and S 12. Thus, proteinases of both the cysteine proteinase and aspartyl proteinase families were found to be increased during the regenerative phase of lung injury, indicating participation inprotein degradation connected with tissue remodelling without any special function of individual cathepsins.
Immunoblot data showed increased catalytic activities of cathepsin B 12 and cathepsin D (present study), largely based on increased enzyme levels. These data demonstrate that differently processed forms of individual cathepsins represent the protein deposits detected immunohistochemically.
No mature form of cathepsin B and only traces of single-chain cathepsin D were found in the alveolar fluid, confirming the lack of any activity of these two proteinases. Furthermore, no activity of cathepsins B, L, S or H was detected in the fluid. Together with the prevalence of CPI activity in BALF, these data disprove the hypothesis of participation of lysosomal cathepsins in protein degradation in the alveolar space. In astudy on cysteine proteinases in alveolar macrophages andBALF, increased activities of cathepsins B and L were described for smokers compared with nonsmokers 29, and Takeyabu et al. 14 reported increased levels of cathepsin L, but not cathepsin B, in BALF from subjects with subclinical emphysema, a subgroup of a smokers cohort. Smoking represents a continuous burden on lung cells, particularly alveolar macrophages, with droplets of tar condensate. The present results regarding the lack of cysteine proteinase activities in the BALF of bleomycin-treated rats indicate a difference between lung damage induced by a single bolus of cytotoxic agent and chronic challenge of alveolar macrophages leading to the development of emphysema. However, procathepsin B was found constitutively and at elevated levels in BALF after bleomycin-induced injury. The coincident maximum procathepsin B levels in BALF and loss of cathepsin B by alveolar macrophages 7 days after bleomycin treatment indicates altered procathepsin B traffic in this cell type. Release of proforms of cathepsins is a common feature of macrophages. Kominami et al. 30 reported that ∼30% of newly synthesised cathepsin B is released by macrophages as the proenzyme, indicating a targeting pathway bypassing the lysosomal compartment. The present authors suggest that, during the inflammatory phase of lung injury, alveolar macrophages are stimulated to release procathepsins as has been described for sodium caseinate-elicited macrophages 30. A secretory mechanism is suggested, but mistargeting of the enzyme cannot be ruled out. Changes in the composition of cells in the alveolar space by influx of monocytes (data not shown) 31 and differentiation of these cells to form macrophages may contribute to an altered pattern of secreted proenzymes 32. The loss of cathepsin D by alveolar macrophages at day 7 of injury strongly resembles the course of changes in cathepsin B levels in this cell type. However, only traces of single-chain cathepsin D were found in the alveolar fluid, indicating rapid degradation of cathepsin D by other proteinases or endocytosis after complexation with proteinase inhibitors.
The high CPI activity may represent part of a simultaneously activated precautionary mechanism against unwanted proteinase attack during inflammation and influx of plasma-derived proteinase inhibitors 8, 33. The increased level of cystatin C observed in the BALF of smokers with subclinical emphysema described by Takeyabu et al. 14 and data on the elevated cystatin C levels produced by alveolar macrophages from smokers 34 are in good agreement with this hypothesis regardless of the underlying disease process. Cathepsin D deposits detected in the extracellular matrix in injured tissue do not preclude the possibility of extracellular cathepsin activities in the micromilieu of matrix proteins 35. Furthermore, cathepsins may disturb the proteinase/antiproteinase balance by destruction of specific proteinase inhibitors 36, 37. This might explain the proteolytic degradation of extracellular matrix and elevated elastase activity found in injured lung tissue 12, 37. The deposits of cathepsin B in epithelial cells and of cathepsin D in myofibroblasts might reflect participation of these proteinases in cellular processes involved in tissue remodelling, i.e. intracellular processes during proliferation or differentiation of cells 38. In a study on bleomycin-induced lung fibrosis in hamsters, it was shown that fibroblast proliferation ceased ∼3 weeks after treatment 39. Differentiation of fibroblasts to myofibroblasts occurs very early in bleomycin-induced lung injury. Zhang et al. 40 described increased numbers of myofibroblasts isolated from rat lungs 7 and 14 days after bleomycin instillation. The present findings of elevated cathepsin D levels during in vitro transdifferentiation of fibroblasts to myofibroblast-like cells and in the course of conversion of cells from a proliferative to a quiescent state support the hypothesis of participation of cathepsin D in differentiation processes in damaged lung tissue.
Taken together, the present data indicate that the transient increase in cathepsin amounts and activities reflect participation of these lysosomal proteinases in regeneration processes in bleomycin-induced lung injury. The imbalance between cathepsin and cysteine proteinase inhibitor activities was eliminated by the end of the phase of tissue remodelling initiated by lung injury.
Acknowledgments
The authors wish to thank P. Peche, I. Peterson and H. Seidel for their excellent technical assistance.
- Received December 5, 2002.
- Accepted April 14, 2003.
- © ERS Journals Ltd
References









