





Abstract
The pathogenesis of pulmonary arterial hypertension (PAH) remains uncertain. Both the serotonin and endothelin (ET) systems are believed to be involved. Recent studies pointed to the importance of the serotonin 2B receptor as a limiting step.
The current authors investigated the lung tissue expression of serotonin receptors and of the serotonin transporter (5‐HTT) by real-time-quantitative polymerase chain reaction in chronic overcirculation-induced PAH in growing piglets, with and without treatment with the dual ET receptor blocker bosentan. Pulmonary haemodynamic changes were described by pulmonary arterial impedance spectra.
Three months after the surgical anastomosis of the left subclavian artery to the pulmonary arterial trunk, there was a shift of the impedance spectra to higher ratios of pressure and flow moduli, with increases in both 0 Hz impedance and characteristic impedance, and these changes were completely prevented by bosentan therapy. There was an increase in the expression of the serotonin 1B receptor. There was no change in the expression of the 5‐HTT, and of the serotonin 2B, 1D, and 4 receptors. The overexpression of the serotonin 1B receptor was partially prevented by bosentan therapy.
The present authors conclude that this early pulmonary arterial hypertension model is characterised by an endothelin receptor-dependent increased expression of the serotonin 1B receptor.
- endothelin
- experimental
- pulmonary arterial hypertension
- pulmonary vascular impedance
- serotonin
- systemic-to-pulmonary shunting
The pathogenesis of pulmonary arterial hypertension (PAH), defined as primary pulmonary hypertension (PPH) with identifiable associated conditions 1, remains incompletely understood. Biological abnormalities have been identified in all the compartments of the pulmonary arterialwall 2. They concern the bone morphogenetic protein-transforming growth factor β (TGF‐β) signalling pathway, the serotonin system, smooth muscle cell potassium channels, extracellular matrix remodelling with increased tenascin and matrix proteinases, monoclonal endothelial cell proliferations in plexiform lesions, inflammatory reactions, activated coagulation, and an endothelium-derived vasodilator (mainly nitric oxide (NO) andprostacyclin)-vasoconstrictor (mainly thromboxane A2and endothelin (ET)) imbalance 2. However, it remainsunknown which of these perturbations initiates the disease. There has been interest in the serotonin system, because of an epidemic of PAH related to the intake of serotoninergic appetite suppressant drugs 3 and the report of increased serotonin levels inPAH patients 4. Serotonin causes pulmonary vasoconstriction, which is believed to be mediated mainly through serotonin receptors 5‐HT1B/1D and5‐HTA, and remodelling, which appear to require the serotonin transporter (5‐HTT) 2, 5. A series of studies point to the 5‐HT1B, rather than the 5‐HT2A receptor, as the mediator of serotonin-induced pulmonary vasoconstriction 5, while an overexpression of the 5‐HTT has been reported in lung tissue from PPH patients6.
Much of the difficulties in unravelling the pathobiology ofPAH result from the impossibility of studying early stages of the disease. The current authors recently reported on theBlalock-Taussig procedure in growing piglets as a model of congenital left-to-right shunting-associated early PAH, characterised by significant pulmonary hypertension, medial hypertrophy, and overexpression of the ET, NO, and vascular endothelium growth factor systems, that could apparently be completely prevented by the dual ET‐A and ET‐B receptor blocker bosentan 7.
While the publication of this work was in progress, a reportdrew attention to the possible determinant role of the 5‐HT2B receptor 8. The present authors therefore reinvestigated lung tissue from the experiments to investigate the expression of all possible components of the serotonin system. Since the haemodynamic data had been stored as high fidelity pressure and flow signals, the pulmonary vascular changes were also reanalysed with impedance spectra calculations, which could possibly be more sensitive to pulmonary vascular remodelling than pulmonary vascular resistance (PVR) determinations 9.
Material and methods
Thirty piglets were included in this study, which had been approved by the Committee on the Care and Use of Animals in Research of the Brussels Free University School of Medicine. Ten of the animals had a sham operation. In the other 20 animals, the left subclavian artery was anastomosed to the pulmonary arterial trunk (the so called “Blalock-Taussig” operation). Two animals died in the postoperative course. The other 28 animals were investigated 3 months later. The placebo-treated shunted animals presented with an increase in PVR and small arterioles medial hypertrophy. These changes were completely prevented by bosentan therapy. The shunted animals had increased expressions of the pulmonary ET, the inducible NO, and the VGEF systems. These results have been reported elsewhere 7.
The instantaneous pulmonary pressures and flow signals were digitised with a sampling rate of 200 Hz, stored and analysed on a personal computer. For each data collection, intervals of five end-expiratory heartbeats were analysed. Pulmonary vascular impedance (PVZ) was calculated from the Fourier series expressions for pressure and flow signals as previously reported 10. In this analysis, the impedance at 0 Hz (ZO) corresponds to total PVR, that is the ratio between mean pulmonary artery pressure (Ppa) and cardiac output (Q). Characteristic impedance (ZC) was calculated as the average of impedance moduli taken between 2 and 15 Hz. The first harmonic modulus (Z1) and the first harmonic phase angle were derived from PVZ spectra. Instantaneous pressure waves were separated into their forward and backward components 11. The reflected wave was characterised by its amplitude and time intervals between the electrocardiographic R wave and the foot, the upward zero crossing, the peak and the downward zero crossing of the waves.
The haemodynamic measurements were obtained after ensuring steady-state conditions (stable heart rate and vascular pressures, which were continuously monitored) for 60 min after clamping of the shunt.
Whole pulmonary tissue messenger ribonucleic acid levels were measured by Sybr Green® (Applied Biosystems, Foster City, CA, USA) real-time-quantitative polymerase chain reaction (RTQ‐PCR). Total ribonucleic acid (RNA) was extracted from frozen samples (400 mg) using TRIzol® (Gibco Life Technologies, Paisley, UK) as outlined by the manufacturer. RNA pellets were dissolved in 400 µL ribonuclease (RNase) free H2O. RNA concentration was measured by ultraviolet spectrophotometry (λ=260 ηm). RNA concentrations were adjusted to 0.25 µg·µL−1 using RNase free H2O and stored at −80°C. Reverse transcription (RT) was performed in a total volume of 20 µL, containing 1 µg of total RNA, 7.5 µM of random hexamers, Buffer 1, 220 µM of eachdeoxyribonucleotide (dNTP), 9 mM dithiothreitol, 20 U RNase inhibitor (Applied Biosystems), and 50 U Superscript enzyme (Gibco BRL, Life Technologies, Merelbeke, Belgium). First strand complimentary deoxyribonucleic acid (cDNA) synthesis was performed in the GeneAmp PCR system 2400 (Perkin Elmer, Foster City, CA, USA) as follows: 22°C for 10 min, 42°C for 1 h and 99°C for 5 min. After RT, final volume was adjusted to 40 µL using RNase free H2O. Samples were stored at −20°C. Using already described sequences for 5‐HT1B ( GenBank® Y11867) , 5‐HT1D (GenBank® Y11868) , 5‐HT2B (GenBank® Z48174) , 5‐HT4 (GenBank® Z48175) and 5‐HTT (GenBank® XM171519) , specific pig primers were designed on Primer Express (Applied Biosystems). Using those porcine specific sequences, primers were produced on an automated synthesiser (Applied Biosystems) according to the manufacturer protocols and purified by high performance liquid chromatography. The primer sequences for Sybr Green® RTQ‐PCR are summarised in table 1⇓. Sybr Green® RTQ‐PCR analysis was carried out using the ABI Prism 5700 sequence detector (Applied Biosystems). Amplification mixes (25 µl) containing 5 ηg of cDNA, 2.5 µl Sybr Green® buffer, 250 µM dNTP, 3 mM MgCl2, 400 ηM of each primer and 0.625 U of Amp Taq Gold Polymerase (Applied Biosystems) were processed as follows: 95°C for 10min, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C. Each cDNA was carried out twice. Each plate included two wells without template (no template control) for every mix to control contamination. For every mix a positive control was also included in duplicate.
Primers used for real-time-quantitative polymerase chain reaction in porcine pulmonary tissue
For RTQ‐PCR, statistical analysis was carried out using the Δ cycle threshold (Ct) value (Ctgene of interest-Ctcorresponding report gene). Relative gene expression was obtained by ΔΔCt methods (ΔCtsample-ΔCtcalibrator) using the sham-operated pig group as a calibrator, for comparison of every unknown sample gene expression levels. The conversion between ΔΔCt and relative gene expression levels was: fold induction=2−ΔΔCt 12.
Values are reported as mean±sem. Effects of shunt and drugs were analysed by a repeated measures analysis of variance (ANOVA). When the F‐ratio of the ANOVA reached a p<0.05 critical value, post hoc comparisons were done using Scheffe's tests 13. A value of p<0.05 was considered statistically significant.
Results
The piglets with a systemic-to-pulmonary shunting during 3months, had, after clamping of the shunt, a persistent pulmonary hypertension with a mean Ppa at 33±1 mm·Hg versus 21±1 mm·Hg in the controls. This increase in Ppa was completely prevented by bosentan therapy 7.
Shunt-induced pulmonary hypertension was associated with an upwards shift of the PVZ spectrum, with increases in ZO, Z1 and ZC, a nonsignificant displacement of the first minimum of the ratio of pressure and flow moduli to higher frequencies, and no change in phase angle (fig. 1⇓, table 2⇓). There was an earlier return of the backward wave, which was of unchanged amplitude and duration (table 2⇓). All these changes in PVZ spectra and wave reflection were completely prevented by bosentan therapy (fig. 1⇓, table 2⇓).

Composite pulmonary vascular impedance spectra in piglets 3 months after a sham operation (•), or systemic to pulmonary artery shunting, with intake of either a placebo (▴) or bosentan (□). a) The shunt was associated with increases of 0 Hz impedance (ZO; *: p<0.05, shunt versus baseline; #: p<0.05 shunt versus bosentan), first harmonic impedance and characteristic impedance (ZC; ¶: p<0.05 shunt versus baseline; +: p<0.05 versus bosentan), all of which were completely prevented by bosentan therapy. b) Phase angle was not different in the three groups.
Pulsatile pulmonary haemodynamics in piglets 3 months after a sham operation, or systemic to pulmonary artery shunting, with intake of either a placebo or bosentan
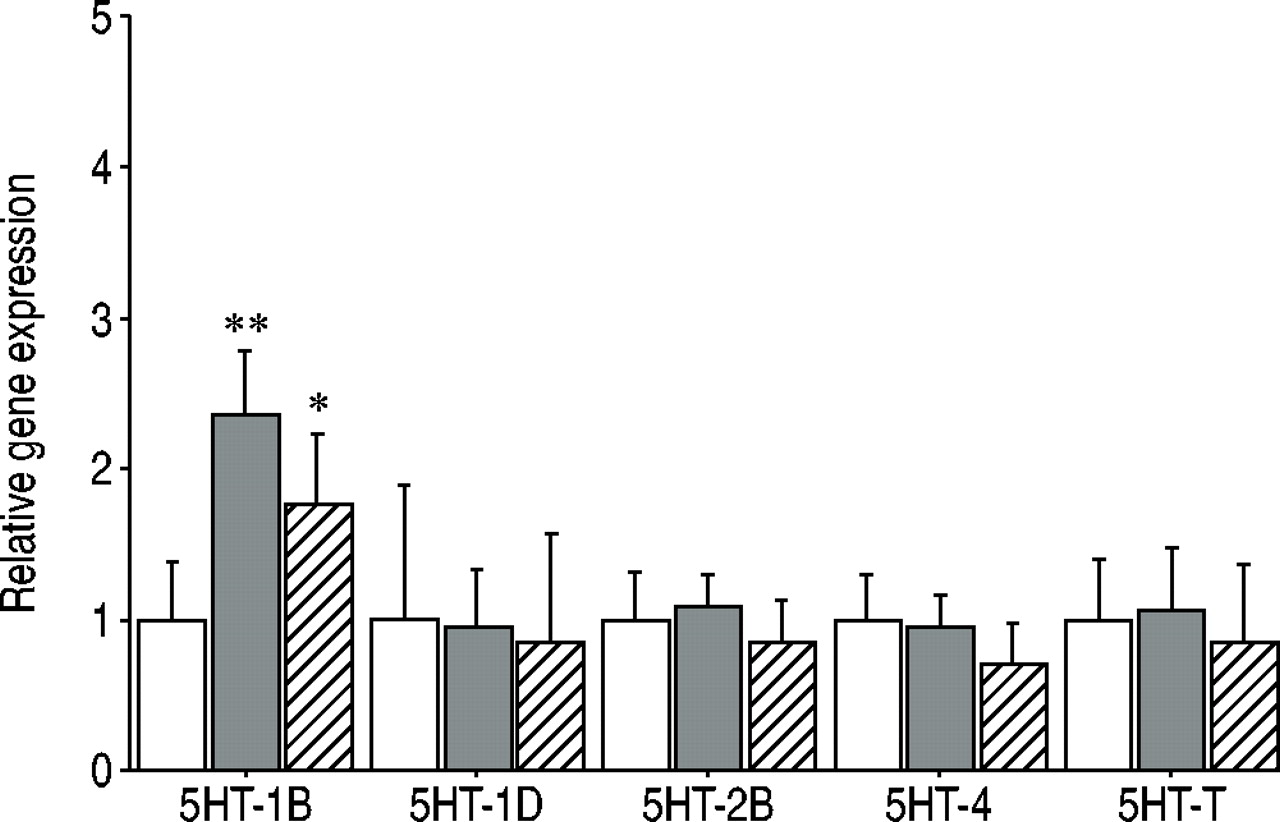
Shunt-induced pulmonary hypertension was associated with increased expression of the 5‐HT1B receptor, from 1.00±0.39 (mean±sem) to 2.35±0.42 units of relative geneexpression (p<0.01) while the expressions of the 5‐HT2B, 5‐HT1D and 5‐HT4 receptors remained unchanged (fig. 2⇓). The expression of the 5‐HTT also remained unchanged. Bosentan therapy was associated with a diminished increase ofthe expression ofthe 5‐HT1B receptor to 1.76±0.45 units ofrelative gene expression (p<0.1 versus placebo-treated pulmonary hypertensive animals, and p>0.05 versus sham-operated controls).
{kind=link}
{kind=link}
Bar graphs showing relative lung tissue messenger ribonucleic acid content for serotonin receptor (5HT)‐1B, ‐1D, ‐2B, ‐4, and serotonin transporter (5HT‐T) in piglets 3 months after a sham operation (□), or systemic to pulmonary artery shunting, with intake of either a placebo (▪) or bosentan (└). Shunt-induced pulmonary hypertension was associated with an increased expression of the 5‐HT1B receptor, and this was partially prevented by bosentan therapy. **: p<0.01, placebo-treated pulmonary hypertensive piglets versus sham-operated controls, *: p<0.05 placebo versus bosentan-treated pulmonary hypertensive piglets.
Discussion
The present results show that chronic overcirculation-induced pulmonary hypertension in growing piglets as a model of early congenital left-to-right shunting-induced PAH, is associated with an increased expression of the 5‐HT1B receptor, with no significant change in the expressions of other components of the serotonin system previously implicated in the pathgogenesis of PAH, such as the 5‐HT2B receptor or the 5‐HTT. Therapy with the dual ET‐1 receptor blocker bosentan partially prevented the increase in 5‐HT1B receptor expression.
Studies on the mechanisms of PAH have been limited untilnow by a lack of satisfactory experimental models. Moststudies have been devoted to pulmonary hypertension induced by chronic hypoxic exposure, or monocrotaline injection in rats. Hypoxic pulmonary hypertension in rodents is associated with an only moderate increase in Ppa 14, and clinical hypoxic pulmonary hypertension differs from PAH inclinical presentation, histology and response to treatment 1. Injection of monocrotaline induces severe pulmonary hypertension 15, but is preceded by permeability lung oedema 16, which is not a feature of PAH. Typical PAH is a classically described complication of congenital heart disease with left-to-right shunts 1, 17.
Previous attempts to reproduce PAH associated with systemic left-to-right shunting often led to disappointingly moderate increases in Ppa, due to poorly reactive animal species, such as dogs, or insufficient duration, pressure, or volume flow of surgically implanted shunting 18–21. In the present study, a Blalock-Taussig operation was performed in as young as possible growing pigs, allowing for shunt flow to increase progressively with the growth of the animals, exposing the pulmonary circulation to as high as possible volume flow and pressure. This approach produced marked pulmonary arteriolar medial hypertrophy and severe pulmonary hypertension, with mean Ppa between 30–40 mm·Hg atnormalised pulmonary blood flow, approaching values found in symptomatic PAH patients 7. The observation ofmedial hypertrophy as the major morphological change inovercirculation-induced PAH is compatible with earlier pathological studies, which showed this aspect to be the most early change, characteristic of less severe and postoperatively reversible case 22. Predominant localisation of pulmonary vascular changes at the smallest pulmonary arterioles explains a previously reported absence of significant changes in partitioning of PVR as determined by single occlusion, and shift of Ppa/Q plots in parallel to higher pressures 7. However, whether the present experimental model really represents congenital left-to-right cardiac shunt-associated PAH remains unproved in the absence of intimal and advential remodelling and plexiform lesions described in patients with this type of pulmonary hypertension 16, 21. Longer periods of observation will be necessary to validate the overcirculation-induced PAH piglet model.
Standard pulmonary haemodynamic investigations rely on mean Ppa and mean Q determinations, with derived PVR calculations, and neglect the information contained in pulmonary pressure and flow waves 9. This information can be apprehended using spectral analysis and derived PVZ computations 9. Accordingly, the current authors wondered whether ET receptor blocker therapy would have completely prevented PVZ changes in the shunted piglets in which mean Ppa/Q relationships were returned to normal 7. In the present study, the shunted piglets presented with a shift of PVZ spectra to higher ratios of pressure and flow moduli and an increase in ZC, which is in keeping with previous observations 19. There was also an earlier return of reflected waves. All these pulsatile haemodynamic changes are compatible with a proximal increase in pulmonary arterial elastance 23. However, the morphometry showing predominantly peripheral medial hypertrophy, culminating at 50–100 µm diameter arterioles 7, would suggest that increased elastance could be entirely accounted for by increased pulmonary artery distending pressures, rather than to vessel wall remodelling.
Overcirculation-induced PAH was associated with increased circulating ET‐1, increased pulmonary tissue gene expression for ET‐1 and ET‐B receptor, and pulmonary endothelial ET‐1 protein, indicating activation of the pulmonary endothelial ET system 7. The dual ET receptor blocker bosentan prevented this early PAH, as also reported in hypoxic- or monocrotaline-induced pulmonary hypertension 14, 15, suggesting that the ET system might be a nonspecific initiating mechanism of pulmonary arterial remodelling.
Several observations point to serotonin as a potential mediator of PAH. Serotoninergic appetite suppressants are associated with an increased risk of developing PPH 3, and plasma serotonin is increased in PAH 4. Serotonin binds to at least 17 different subtypes 5. Vasoconstricting effects of serotonin appear to be mediated mainly by the 5‐HT2A receptors in the systemic arteries, and by the 5‐HT2B or the 5‐HT1D receptors in the pulmonary arteries 6. It has been shown that the circulating levels of serotonin measured in PAH patients contract isolated human arterioles, and that this effect is mediated by the 5‐HT1B receptor 24. On the other hand, serotonin exerts potent mitogenic effects on pulmonary artery smooth muscle cells that appear to require the expression of a 5‐HTT 2, 5. This 5‐HTT has been shownto be overexpressed in lungs from transplanted PAH patients 6.
Most recently Launay et al. 8 reasoned that the 5‐HT2B could also be involved. Indeed, the active metabolite of dexfenfluramine, which is the serotoninergic appetite suppressant drug most clearly related to an increased risk of developing PAH 3, is a selective 5‐HT2B receptor. The authors observed that chronic hypoxia-induced pulmonary hypertension in mice was associated with vascular proliferation, elastase activity and TGF‐β levels, that were potentiated by dexfenfluramine treatment. In contrast, hypoxic mice with genetically or pharmacologically inactive 5‐HT2B receptors did not develop pulmonary hypertension and associated elastase and TGF‐β changes 8. This observation would explain why dexfenfluramine is associated with PAH while it's inhibiting effects on the 5‐HTT, shared by serotoninergic antidepressant drugs, should actually protect against pulmonary arterial remodelling.
The current authors could not find primers to investigate the expression of the 5‐HT2A receptor. Therefore, the importance of the 5‐HT2A receptor in this PAH model is not known. Another important limitation to the present study is that no antibodies were available to measure tissue serotonin and transporter protein levels, which would have allowed more to be known about the functional state of all the components of the serotonin receptors. In addition, the question of the nature of possible interactions between ET and serotonin receptors remains unresolved.
However, the present results are compatible with a participation of the serotonin system, essentially by means of an increased expression of the serotonin receptor 5‐HT1B rather than that of the serotonin receptor 5‐HT2B or of the serotonin transporter, in the most early stages of congenital left-to-right shunt-induced pulmonary arterial hypertension.
Acknowledgments
Supported by grant number 3.4567.00 from the Fonds de la Recherche Scientifique Médicale, and by a grant from the Foundation of Cardiac Surgery, Belgium. B. Rondelet was a fellow of the Erasmus Foundation, Brussels, Belgium. S. Motte is “Aspirant” from theFonds National de la Recherche Scientifique, Belgium.
- Received April 2, 2003.
- Accepted May 22, 2003.
- © ERS Journals Ltd
References









